1. Introduction
Non-alcoholic fatty liver disease (NAFLD) is a global health issue, particularly in children who have adopted a sedentary lifestyle and received home or virtual schooling. NAFLD is characterized by excess fat in the liver cells, known as hepatocytes. The chronic stress caused by lipid accumulation within the liver may lead to non-alcoholic steatohepatitis (NASH), an inflammatory type of fatty liver. NASH is considered a risk factor for liver fibrosis, cirrhosis, and eventual hepatic carcinogenesis
[1]. The prevalence of NAFLD is increasing over time with an estimated global rate of about 38%
[2]. The etiology behind NAFLD is complicated due to the multifactorial nature of its pathogenesis. While the existence of a metabolic disorder plays a role, other factors in a metabolically healthy individual may contribute to NAFLD development. Several genetic mutations have been identified as playing a role in NAFLD development and progression to NASH, including mutations in the Patatin-Like Phospholipase Domain Containing 3 (PNPLA3) and Membrane-Bound O-Acyltransferase Domain Containing 7 (MBOAT7) genes
[3]. Furthermore, a growing body of evidence supports an epigenome contribution to NAFLD development and susceptibility
[4].
Scientists have come to recognize the crucial role of non-coding RNA (ncRNA) in cellular functions. NcRNA is a class of RNA that does not code for proteins and comprises most of the human genome. Since its discovery, ncRNAs have been used in therapeutic interventions and as diagnostic biomarkers for multifactorial diseases, including liver disorders
[5]. Several types of ncRNAs have been shown to contribute to chronic liver diseases, including micro-RNA (miRNA), long non-coding RNAs (lncRNAs), P-element-induced wimpy testis (PIWI)-interacting RNAs (piRNAs), and circular RNAs (circRNAs). The mechanism of action varies for different ncRNAs when regulating gene expression. LncRNA acts at the genomic level and participates in chromatin remolding and histone modification, which alter gene expression
[6]. MiRNA plays an essential role in post-translational modification by interacting with the 3′-untranslated region (3′UTR) of target mRNAs and inducing the degradation or hindering the translation of the target mRNA into a functional protein
[7]. Upon its discovery, circRNA was known to mainly function as an miRNA sponge leading to miRNA suppression. CircRNAs were also found later to interact with proteins involved in alternative splicing and gene transcription fields
[8]. PiRNAs were initially identified as regulators of transposon silencing in
Drosophila ovarian germline cells
[8]. PiRNAs were shown later to regulate mammalian gene expression by interacting with the PIWI protein family and forming a piRNA-induced silencing complex (piRISC). PiRISC plays a vital role in protein expression, genome rearrangement, and reproductive stem-cell maintenance
[9].
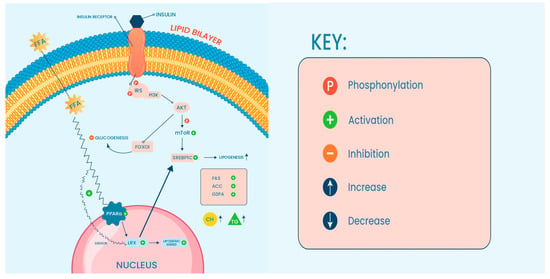
With the global rise of NAFLD prevalence, it is anticipated that NASH will also become increasingly burdensome. Consequently, increased morbidity and mortality associated with liver disease are also expected. Understanding molecular mechanisms underlying the disease will provide a means to develop effective therapeutic interventions and may provide insight into how these conditions can be prevented from developing in the future (Figure 1).
Figure 1.
Key signaling pathway deregulation is seen in NAFLD (see text for details).
2. Liver Cellular Function and Development of NAFLD/NASH
The liver is composed of several cell types; every cell population has a distinct function in regulating liver function and homeostasis. Most liver cells are hepatocytes (HCs), comprising approximately 80–85% of the liver mass. The rest of the liver mass is made up of Kupffer cells (KCs), hepatic stellate cells (HSCs), liver sinusoidal cells (LSECs), and cholangiocytes (biliary cells)
[10]. Studies have shown that all liver cells can be involved in the development of and progression to NAFLD with each cell type having a distinct role in the pathology of disease progression.
The primary functional unit of the liver is the hepatocyte in which most of the cellular functions take place, including the metabolism of macromolecules (carbohydrate, lipid, and protein), the execution of detoxification pathways, and the activation of immune cells. The pathological role of the hepatocytes in developing NAFLD/NASH is well documented, and studies agree that hepatocytes can act as an inflammatory activator in the liver
[11]. With most metabolic activity occurring in the hepatocytes, these cells can be exposed to various stressors in the regular physiological state and are vulnerable to injury. The ingestion of toxins, drugs, alcohol, and dietary overload can all lead to hepatocyte injury, which results in the activation of several inflammatory cascades. Injured hepatocytes are more susceptible to cell death and thus trigger the release of reactive oxygen species (ROS), inflammatory cytokines, and repair signals. This also leads to hepatocyte organelle dysfunction, including mitochondrial damage, lysosomal disruption, and endoplasmic reticulum stress. Ultimately, this increases the intercellular communications between the hepatocytes and the other cells within the liver, especially the KCs
[12][13][14][12,13,14]. Kupffer cells are the liver-resident macrophages. KCs release several chemokines and cytokines upon activation to enhance their polarization in physiological and pathological conditions. Persistent inflammation of the liver tissue increases the activation of the pathological type of KCs, eventually leading to hepatocellular damage
[15]. Injured sinusoidal cells caused by liver steatosis can also activate KCs. Injured LSECs are inflammatory mediators that contribute to the recruitment of other inflammatory cells, leading to liver injury
[16]. Stellate cells, conversely, are dynamic and can be found in inactive or active states. Quiescent stellate cells in the healthy liver act as a storage site, mainly for fat and vitamin A as a retinyl ester. In contrast, the active cells act as the primary fibrogenic cells during liver injury
[17].
Intercellular communications between liver cells are complex and dependent on the microenvironmental state. Cell response to metabolic or immune stimuli will change over time depending on the type of signal released, the gene expression, and the duration of the inflammatory event
[18]. Several types of liver cells are implicated in the complex process involved in the development of NAFLD and subsequent progression to NASH.
3. NcRNA and Intercellular Communication in Liver Cell Populations
With the advancement in RNA-sequencing technologies (RNA-seq), several studies have shifted focus to identify functional RNA transcripts that can reveal molecular mechanism dysfunction in liver diseases, including NAFLD. These functional or ncRNAs are classified based on length, structure, function, and genomic location. Several classes of ncRNA have been found to modulate gene expression within the liver and to participate in protein regulation. Among these ncRNAs are miRNA, piRNA, ciRNA, and lncRNA.
Intercommunication between liver cells is a complex process in both physiological and pathological states. It could be assumed that the proinflammatory environment within the liver caused primarily by HSC signals will activate the resident liver macrophages (i.e., KCs) and possibly other cell types involved in NAFLD/NASH pathophysiology. The ncRNA molecules play a crucial role in regulating the inflammatory responses of liver cells in healthy and compromised liver tissue.
4. NcRNA and the Dysregulated Pathways Related to NAFLD/NASH
The vicious cycle of insulin resistance and lipid accumulation leads to a cascade of biological events as NAFLD progresses to NASH, including oxidative stress, lipotoxicity, inflammation, and eventually fibrosis. ncRNAs, including lncRNAs, circRNAs, and miRNAs, are known to be significant regulators in the pathogenesis of NAFLD as indicated by high-throughput sequencing methods
[19][67]. More studies are needed to fully comprehend how these molecules interact within and between the hepatocytes concerning the pathogenesis. A list of ncRNAs involvement in NAFLD is listed in
Table 1.
Table 1.
List of ncRNAs involved in NAFLD.
|
Non-Coding RNA
|
Expression
|
Primary Function
|
Reference
|
|
miR-96-124-182-183
|
Upregulated
|
These ncRNAs modulate lipid synthesis through SREBP-1c
|
[20][21][89,90]
|
|
miR-206
|
Downregulated
|
[22][91]
|
|
Gm16551
|
Downregulated
|
[23][92]
|
|
lncHR1
|
Downregulated
|
[24][93]
|
|
miR-122-3075-21-192-5p
|
Upregulated
|
These ncRNAs are involved in the stimulation or inhibition of HCs and HC-related injuries
|
[25][26][27][28][29][30[33][46,49][,5431][,5632,57],58,74,77,94]
|
|
piR-823
|
Upregulated
|
[34][85]
|
|
circBNC2
|
Upregulated
|
[35][78]
|
|
circ_0071410
|
[32][77]
|
|
circPI4KB
|
[31][74]
|
|
circRNA_0046367
|
Downregulated
|
[36][72]
|
|
miR-26
|
Downregulated
|
miR-26 expression depends on LXRs and has a protective role
|
[37][95]
|
|
miR-34a
|
Upregulated
|
Both ncRNAs were found to be associated with the regulation of hepatic fat content through PPARα
|
[38][96]
|
|
circRNA_0046366
|
Downregulated
|
[38][96]
|
|
miR-451
|
Downregulated
|
Involved regulation of glucose homeostasis
|
[39][97]
|
|
miR-103/miR-107
|
Upregulated
|
[40][98]
|
|
LncARSR
|
Upregulated
|
The expression of LncARSR correlates with the activated state of PI3K/AKT/mTOR pathway
|
[41][99]
|
|
miR-155
|
Upregulated
|
These ncRNASs have an influence on the induction of inflammation via NF-κB pathway
|
[42][43][100,101]
|
|
Platr4
|
Upregulated
|
[44][102]
|
|
LncTNF
|
Upregulated
|
[45][103]
|
|
miR-125
|
Upregulated
|
The expression of these ncRNAs contributes to the activation of pro-inflammatory mediators
|
[46][104]
|
|
circFBXW4
|
Downregulated
|
[14]
|
|
miR-29b
|
Downregulated
|
These ncRNAs are involved in fibrosis mediation through TGF-β1
|
[33][94]
|
|
H19
|
Downregulated
|
[47][48][65,105]
|
|
piR-823
|
Upregulated
|
[34][85]
|
|
miR-26a-143-145
|
Upregulated
|
These ncRNAs have been found to be modulators in insulin sensitivity
|
[49][50][51][106,107,108]
|
|
AK085787-uc009kuu.1-uc008txr.1
|
Upregulated
|
[23][92]
|
|
CircRNF111-circRNA_0001805
|
Downregulated
|
[52][53][73,109]
|
|
CircRNA SCAR
|
Upregulated
|
This ncRNA may play a role in activating liver fibroblasts
|
[54][71]
|
5. The Role of ncRNA in NAFLD-Associated Metabolic Dysfunction
Recently, it was suggested to rename NAFLD to metabolic-associated non-alcoholic fatty liver disease (MAFLD)
[55][132]. The renaming of this disease would reflect the pathological factors that contribute to the development of fatty liver and would provide a comprehensive understanding of the disease progression and management. The new proposed name also highlights the role of metabolic drivers in developing liver steatosis. The established criteria and recommendations for NAFLD screening generally apply to individuals with a pre-excitant metabolic disease, in particular for diabetic or obese patients and people with hyperlipidemia. These metabolic syndromes are considered a risk factor for NAFLD, even in asymptomatic patients. The presence of one or more metabolic risk factors will lead to overall systematic disturbance, insulin resistance, and an increase in lipolysis. Thus, an increase in FFA being sent to the liver will promote hepatic-triglyceride synthesis, reduce FFA oxidation, and enhance fat deposition within the liver
[56][133]. It was estimated that obesity and hyperlipidemia accounted for most metabolic comorbidities associated with NAFLD, with a prevalence of 50% and 70%, respectively. Diabetes accounted for more than 20% of the metabolic comorbidities, while other metabolic induced-NAFLD accounted for approximately 40% of the global prevalence
[57][134].
Obesity is linked to impaired blood glucose tolerance as well as to hyperinsulinemia, which increases the risk of developing diabetes and hyperlipidemia
[58][135]. Obese patients might be present with or without metabolic syndrome, but this will not rule out fatty liver
[59][136]. This complex crosstalk between metabolic syndromes is not fully understood, but it has been proposed that impaired insulin signaling seen in patients with a preexistent metabolic condition is the key player in NAFLD progression. Yet, the dysregulation in insulin secretion or function is linked to multiple metabolic imbalances in the liver, the pancreas, the muscles, and the adipose tissue
[60][137].
A large body of evidence indicates that the role of ncRNA in metabolic modulation involving insulin signaling
(Figure 4) has been shown to prevent or contribute to metabolic abnormalities within hepatic cells. Using microarray analysis, Trajkovski et al. identified miR-103 and miR-107 upregulation correlating with insulin sensitivity in liver tissue of obese mice
[40][98]. Interestingly, loss of function analysis of miR-103/miR-107 lowered blood glucose levels, enhanced adipose and liver insulin sensitivity, and decreased hepatic glucose production by targeting caveolin-1 (Cav1) mRNA, an essential regulator in glucose and lipid homeostasis.
A study conducted in 2011 showed a significant association between miR-143 and dysregulation of glucose metabolism in a transgenic mouse model through insulin resistance induction
[51][108]. Molecular analysis of the mode of action of miR-143 identified an under-reactive AKT signaling within the liver tissue of obese mice, suggesting that miRNA may impair AKT phosphorylation and reduce insulin sensitivity. Furthermore, miR-143-145 deficient mice showed improved insulin sensitivity and active AKT signaling. In terms of insulin-resistant development, a member of the oxysterol binding protein family (OSBP) known as ORP8, seems to be a direct target for miR-143. OSBP, among other functions, regulates cholesterol efflux and activates AKT signaling. In this study, miR-143-145 deficient mice showed an almost 2-fold increase of hepatic ORP8 compared to control mice, which increased or accelerated hepatic insulin action. Other miRNAs were also found to be deregulated in obese animal models as well as in human hepatic tissue and were found to regulate both glucose and lipid homeostasis which included miR-26a
[49][106], miR-206
[22][91], and miR-451
[39][97].
It has been found that lncRNAs, on the other hand, were also linked to impaired metabolic signaling related to obesity. Yang and his group used integrative transcriptomic approaches to identify lncRNA-mRNA networks crucial for metabolic regulation. The group identified lncRNA Gm16551 to be enriched in the liver, and its expression was lost in obese mice. To determine the nutrient-sensitive lncRNA, nutrient and fasting-induced hepatocytes were used. The expression of three lncRNA including AK085787, uc009kuu.1, and uc008txr.1 seemed to be nutrient-dependent
[23][92] and tended to be induced by insulin. The precise role of these lncRNAs in response to insulin is still not elucidated. However, the data suggest that metabolic regulation might be controlled by a single or a group of lncRNA and they could have a role in the pathogenesis of insulin-related pathologies.
Deep sequencing analysis followed by gain and loss of function approaches found that the circRNA was protective in metabolic syndrome patients
[53][109]. In this study, the circRNF111 molecules were found to interact with miR-143 and inhibit the mediated insulin resistance caused by miR-143 overexpression. In another study, the group also identified IGF2R as a direct target for miR-143
[50][107]. These data agree with what was previously reported by Jordan and his group regarding miR-143 as discussed earlier
[51][108]. Considering the findings from these recent studies, it is evident that a holistic approach should be followed when investigating metabolic drivers, ncRNA molecules, and their roles in the development of NAFLD/MAFLD.