Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Vijay Kumar and Version 2 by Camila Xu.
Pattern recognition receptors (PRRs) play critical roles in embryonic development, immune homeostasis, neurodevelopment, and neurodegeneration. PRRs are highly conserved germline-encoded proteins that recognize microbe/pathogen-associated molecular patterns (MAMPs or PAMPs) and death/damage-associated molecular patterns (DAMPs); thus, they regulate innate and adaptive immunity and contribute to the pathogenesis of many diseases ranging from infections to cancers.
- PRRs
- cGLRs
- cGAS
- STING
- type 1 IFN
1. Introduction
Pattern recognition receptors (PRRs) play critical roles in embryonic development, immune homeostasis, neurodevelopment, and neurodegeneration. PRRs are highly conserved germline-encoded proteins that recognize microbe/pathogen-associated molecular patterns (MAMPs or PAMPs) and death/damage-associated molecular patterns (DAMPs) [1][2][3][1,2,3]; thus, they regulate innate and adaptive immunity [1][4][5][1,4,5] and contribute to the pathogenesis of many diseases ranging from infections to cancers [5][6][7][8][9][10][11][5,6,7,8,9,10,11]. Traditionally, PRR family members have included toll-like receptors (TLRs), C-type lectin receptors (CLRs), nucleotide-oligomerization domain (NOD)-like receptors (NLRs), absent in melanoma-2-like receptors (ALRs), retinoic acid-inducible gene (RIG)-1-like receptors (RLRs), C-type lectin receptors (CLRs), and different scavenger receptors (SRs) [2][9][12][13][14][15][16][17][2,9,12,13,14,15,16,17]. Cytosolic guanosine monophosphate (GMP)-adenosine monophosphate (AMP) synthase (cGAS)-like receptors (cGLRs) have been recently recognized as new PRRs that potentially interact with other PRRs to regulate immune homeostasis.
2. Emergence of cGLRs (Harboring cGAS and STING) as a Novel Family of PRRs
cGLRs have emerged or evolved as critical PRRs providing defense through different mechanisms in prokaryotes and eukaryotes, including humans. Table 1 shows different cGLRs and their known ligands. The first enzyme responsible for synthesizing cyclic guanosine monophosphate (GMP)-adenosine monophosphate (AMP) (cGAMP) or cyclicdinucleotides (CDNs), dinucleotide cyclase (DncV), or cGAS-DncV-like nucleotidyltransferase (CD-NTase), evolved in bacteria and has been reported in Vibrio cholerae. CD-NTases generating CDNs, like animal cGASs, are very diverse proteins numbering over 6000 to date [18]. In addition to producing CDNs, bacterial CD-NTases can also produce cyclic trinucleotides (CTNs) and linear oligonucleotides [19][20][19,20]. Although DncV lacks primary sequence homology to h-cGAS (synthesizes 2′3′-cGAMP), it synthesizes conventional 3′3′- cGAMP involved in bacterial chemotaxis [21][22][21,22]. Bacteria encode thousands of cGAS/DncV-like NTases (CD-NTases) to control their highly divergent anti-phage defense system. Furthermore, DncV NTases, cGAS homologs, and 2′-5′-oligoadenylate synthase 1 (OAS1, having aspartic acid (Asp) in place of E225) have five highly conserved active sites [22]. The CD-NTases of the cyclic oligonucleotide-based antiphage signaling system (CBASS, analogous to the cGAS/STING signaling pathway of metazoans) that are activated by the binding of a folded fragment of RNA share distant homology with the cGAS [23][24][25][26][27][23,24,25,26,27]. Furthermore, CBASS-activating bacteriophage RNA (cabRNA), via binding to the surface of positively charged CdnEO3 cyclase (a bacterial cGAS), promotes cGAMP synthesis to activate the CBASS-mediated immune response [28].Thus, the CBASS in the bacteria producing cGAMP serves as a defense system against bacteriophages [29]. The bacterial cGAS or CBASS comprises a four-gene operon that encodes bacterial cGAS, which is associated with phospholipase, two enzymes with the eukaryotic-like domains E1 and E2, and a Janus kinase-binding protein (JAB) domain, which provides resistance against several bacteriophages. cGAMP production activates phospholipases to disintegrate membranes and bacterial cell death before the completion of the bacteriophage life cycle or the production of mature bacteriophage [29].Table 1. Different cGLRs (including mammalian cGAS), their ligands, cleaved products (cyclic dinucleotides, and CDNs such as 2′3′-cGAMP and/or 3′3′-cGAMP) activating STING and downstream signaling molecules throughout the animal kingdom from metazoans to mammals. Kindly see the article text and referenced articles for details.
| Species | cGLR | Ligand/Product | STING | IKKε | TBK1 | NF-κB | IRF3 | IRF7 |
|---|---|---|---|---|---|---|---|---|
| Homo sapiens/Human | cGAS | dsDNA/2′3′-cGAMP | + | + | + | + | + | + |
| Mus musculus/Laboratory mouse | cGAS | dsDNA/2′3′-cGAMP | + | + | + | + | + | + |
| Gallus gallus/Red junglefowl | cGAS | dsDNA/2′3′-cGAMP | + | + | + | + | - | + |
| Xenopus tropicalis/Western clawed frog | cGAS | dsDNA/2′3′-cGAMP | + (without CTT domain) | + | + | + | + | + |
| Branchiostoma floridae/Florida lancelet | 2 cGAS homologs | dsDNA/2′3′-cGAMP | +, 2 STING candidates (STING-1 and STING-2, | + | + | + | - | - |
| Danio rerio/Zebrafish | 2 cGAS | Cytosolic dsDNA/2′3′-cGAMP | + | + | + | + | + | + |
| Strongylocentrotus purpuratus/Pacific purple sea urchin | ? | ? | - | + | + | + | - | - |
| Ceanorhabidits elegans/Roundworm | - | - | - | + | - | - | - | - |
| Drosophila melanogaster/Common fruit fly | Dm-cGLR1 and Dm-cGLR2 | dsRNA/3′2′-cGAMP and 2′3′-cGAMP | + | - | + | + | - | - |
| Tribolium castaneium/Red flour beetle | Tc-cGLR | dsRNA/2′3′-cGAMP | + | - | + | + | - | - |
| Microplitis demolitor/Wasp | Md-cGLR | dsRNA/2′3′-cGAMP | + | - | + | + | - | - |
| Frankliniella occidentalis/Western flower thrip | Fo-cGLR | dsRNA/2′3′-cGAMP | + | ? | + | + | - | - |
| Nicrophorus vespilloides/Common sexton beetle | Nves-cGLR | dsRNA/2′3′-cGAMP | + | ? | + | + | - | - |
| Aethina tumida/Small hive beetle | At-cGLR | dsRNA/2′3′-cGAMP | + | ? | + | + | - | - |
| Asbolus verrucosus/Blue death feigning beetle | Av-cGLR | dsRNA/2′3′-cGAMP | + | ? | + | + | - | - |
| Trichogramma pretiosum/Wasp | Tp-cGLR | Unknown/2′3′-cGAMP | + | - | + | + | - | - |
| Chlamydophila felis/Cat flea | Cf-cGLR | Unknown/2′3′-cGAMP | ? | ? | + | + | ? | ? |
| Pocilloporidae damicornis/Cauliflower coral | Pd-cGLR | Unknown/2′3′-cGAMP | + | - | + | + | - | ? |
| Crassostrea gigas/Pacific oyster | Cg-cGLR1 | dsDNA/2′3′-cUA | + | + | + | + | -, but have cgIRF1 | -, but have cgIRF8 |
| Crassostrea virginica/Eastern oyster | Cv-cGLR1 | dsDNA/2′3′-cUA | + | + | + | + | -, but have cgIRF1 | -, but have cgIRF8 |
| Cv-cGLR2 | Unknown/Unknown | |||||||
| Sp-cGLR1 | dsRNA/3′3′-cUA | |||||||
| Stylophora pistillata/Stony coral | Sp-cGLR2 | Unknown/2′3′-cGAMP | + | - | ? | + | ? | ? |
| Sp-cGLR3 | dsRNA/3′3′-cAA | |||||||
| Amphimedon queenslandica/Sponge | ? | ? | - | - | + | + | - | - |
| Exaiptasia pallida/Glass anemone | Ep-cGLR | Unknown/2′3′-cGAMP | + | - | + | + | - | - |
| Monosiga brevicollis/Choanoflagellate | Mb-cGAS | dsDNA/3′3′-cGAMP | + | - | - | - | - | - |
| Nematostella vectensis/Starlet Sea anemone | Nv-cGAS or nvA7SFB5.1 | dsDNA/3′3′-cGAMP | + | - | + | + | - | - |
| Hydra magnipapillata/Hydra vulgaris | Hv-cGLR | dsRNA/2′3′-cGAMP | +, 3 STING candidates (STING-1, STING-2, and STING-3) | + | - | + | - | - |
+ = present; - = absent; ? = not known/unsure; cUA, cyclic UMP-AMP; cAA, cyclic diAMP.
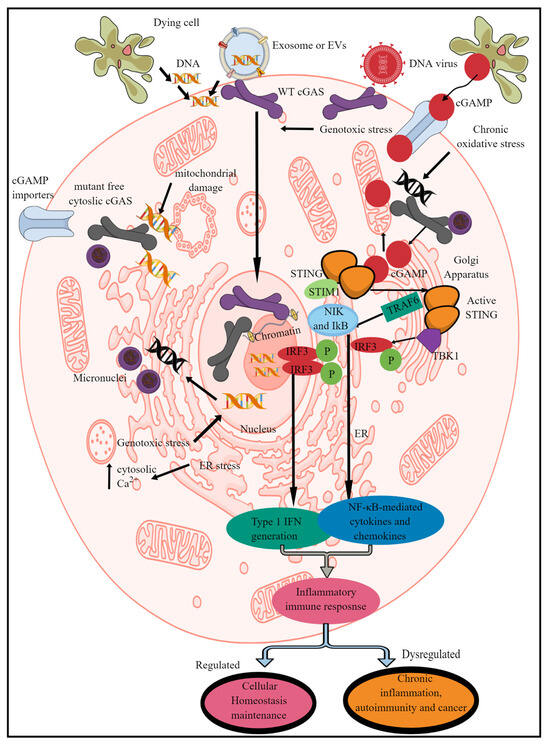
Figure 1. Schematic representation of mammalian cGLR (cGAS/STING signaling) activation and downstream effects. cGAS recognizes cytosolic dsDNA, which can be foreign (pathogen-derived) or self- or adjacent-cell-generated (Mt-DNA and micronuclei generated due to genotoxic or oxidative stress) as a PAMP/DAMP. The enzymatic activity of cGAS cleaves cytosolic dsDNA into CDNs called cGAMP. STING recognizes cGAMP, which induces its transfer to ERGIC and the Golgi apparatus. For example, in the non-activated stage, it remains in the ER via binding to STIM-1 to retain its position in the ER membranes. The STING translocation to the Golgi apparatus phosphorylates (denoted by “P” in figure),TBK1 which induces IRF3 phosphorylation and induction of IRF3-dependent type 1 IFN genes. Active STING also phosphorylates TRAF6, which induces downstream signaling molecules (NIK and IκB) to initiate the transcription of NF-κB-dependent pro-inflammatory genes. Cells such as immune cells also express cGAMP importers, which import cGAMP generated by distant cells to initiate STING signaling without the involvement of cGAS itself. Notably, cGAS located in the nucleus is unable to recognize and bind to nuclear DNA to initiate the pro-inflammatory immune response. Kindly see the text for details.