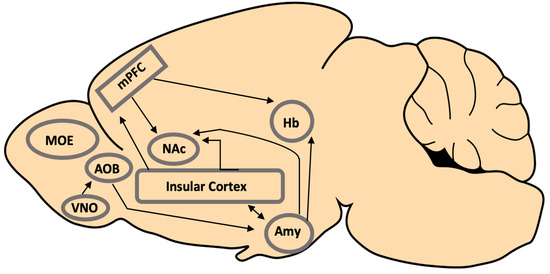
Figure 12. Regions and projections in the mouse brain that are proposed to be involved in the processing and mediation of the behavioural response to the pathogen risk. It is likely that additional systems involved in social information processing are also involved. MOE (main olfactory epithelium); VNO (vomeronasal organ); AOB (accessory olfactory bulb); mPFC (medial prefrontal cortex); NAc (nucleus accumbens); Hb (habenula); Amy (amygdala).
2.1. Medial Amygdala
The medial amygdala (MeA) is a critical region for the decoding of social stimuli and processing of appropriate behavioural responses. In rodents, both the main and accessory olfactory bulbs have projections directly to the MeA
[16][17][18][95,96,97]. The majority of the olfactory projections to the MeA come from the accessory olfactory bulb (and indirectly from the VNO), suggesting that the MeA is involved in the processing of non-volatile chemical sensory information primarily detected by the accessory olfactory bulb
[19][20][98,99]. In turn, the MeA is functionally connected to a broad network of limbic regions and the hypothalamus, suggesting that the MeA is involved in the transformation of sensory information into specific behavioural responses
[19][21][22][98,100,101].
The MeA is differentially implicated in both appetitive and aversive/avoidance behaviour, with it and its projections mediating appetitive responses to reproductive stimuli, as well as the processing and avoidance of aversive odours
[22][23][24][101,102,103]. The integration of olfactory signal and social stimuli processing in the MeA likely makes the region integral in both the detection of, and the behavioural response to, parasitic risk.
In male rats, odour from conspecifics treated with LPS has been shown to induce avoidance response in healthy rats. Arakawa et al. (2010)
[25][104] found that the MeA and multiple brain regions that it projects to were activated (including the bed nucleus of the stria terminalis (BNST) and the paraventricular nucleus of the hypothalamus (PVN)) in rats exposed to the odour of a sick LPS-treated conspecific, exemplifying the role of the MeA as a hub of social information processing. In this regard, whether the responses were elicited by exposure to sickness, which elicits a constellation of responses, or to LPS treatment per se remains to be determined.
Together, these findings indicate that the MeA serves as one of the first processing points of conspecific chemosignals, facilitating the detection of pathogenic threats and eliciting subsequent behavioural responses, such as approach and mate choice. As such, the MeA is crucial to the detection of social signals that are indicative of pathogenic threats and likely plays a key role in triggering behavioural avoidance and disgust responses through its projections to other brain regions. Which specific brain regions, and particularly how these may be integrated to facilitate behavioural responses, remains to be determined.
2.2. Insular Cortex
The insular cortex is a region heavily implicated in the disgust response and the mediation of various aversive/avoidance behaviours through connections with both social brain regions and reward regions
[26][27][28][29][91,106,107,108]. The insular cortex can be divided into two distinct regions: the posterior insular cortex and the anterior insular cortex. The posterior insular cortex projects to the sensory and limbic brain regions, while the anterior insular cortex mainly projects to the social brain network, including the medial prefrontal cortex, the nucleus accumbens, and the amygdala
[29][30][108,109].
The insular cortex is directly involved in pathogen avoidance, with inactivation of the posterior insular cortex eliminating the avoidance response to conspecifics treated with a viral mimetic in both female and male rats
[31][110]. A recent study by Min et al. (2023)
[30][109] demonstrated that lesions on the anterior insular cortex of male mice resulted in an inability to distinguish between a cage mate and a novel mouse. Due to the role of the anterior insular cortex in processing information leading to social recognition, this region may also be involved in the processing of infection cues and the recognition, and avoidance, of parasitized individuals.
A study investigating the correlation between the stereotyped facial expression and the affective state in mice found that exposure to a toxin (lithium chloride) elicited facial expressions that were associated with disgust and corresponded to increased activity in the anterior insular cortex
[32][111]. It should be noted that to date, there are no reports that exposure to the odours of infected individuals elicits disgust-like facial expressions in rodents. More detailed machine-learning assessments of facial responses to infection cues are needed.
In mice, the insular cortex also receives monosynaptic input from the MeA, with this anatomical connection being considered to be important in valence processing and emotional regulation
[30][33][34][109,112,113]. Additionally, the expression of a disgust-like response to toxins in mice involves neural pathways from the anterior insula to the medial and basolateral amygdala
[35][114]. These neural projections between the insular cortex and the MeA denote a functional link between the two regions in the processing of aversive social stimuli and disgust-like responses.
2.3. Habenula
The habenula is a highly conserved limbic brain region implicated in divergent motivational states and cognition
[36][115]. Subnuclei of the habenula are thought to be involved in the processing of avoidance responses to various stimuli. The lateral habenula is central to the brain’s reward system and inhibits dopamine release in the midbrain to signal negative valence
[37][116]. Studies using fibre-photometry found that the lateral habenula of mice shows increased neuronal activity when the mouse was presented with a bitter taste, pain, and aggression from a conspecific
[38][117]. In addition to this, the lateral habenula is also involved in the regulation of social behaviour, with chemogenetic activation of the region resulting in a reduction in social behaviour and social interactions
[14][39][93,118]. Wang et al. (2017)
[38][117] suggested that aversion processing in the lateral habenula is highly influenced by learning, as a prolonged positive reward inhibited lateral habenula activity in mice that were conditioned to an aversive stimulus. This suggests that lateral habenula activity in response to, and processing of, aversive stimuli is experience dependent.
Weiss et al. (2023)
[40][87] further implicated the lateral habenula in the detection and avoidance of the urine of LPS-treated conspecifics. Exposure to LPS-treated conspecific urine resulted in lateral habenula activation four times stronger than the activity seen during exposure to control urine. Although the habenula is often described as the region associated with hedonia and anhedonia, more consideration should be focused on this region when studying disgust and aversion to pathogens, as positive and negative affect likely heavily influence adaptive responses to stimuli that evoke disgust.
2.4. Nucleus Accumbens
Recent research has explored the role of the nucleus accumbens (NAc) in the avoidance of aversive stimuli. The NAc is integral in the detection of aversive stimuli and translating motivation into action. The NAc is a convergence point of both reward and aversion circuitry, receiving projections from the ventral tegmental area, the medial prefrontal cortex, and the basolateral amygdala
[41][42][43][44][119,120,121,122]. Specifically, dopamine signalling from the ventral tegmental area has been linked to reward and aversion processing in the NAc, while glutamatergic signalling from the thalamic paraventricular nucleus to the NAc is implicated in aversion
[45][46][123,124]. He et al. (2023)
[41][119] found that Tac1 neurons in the medial shell of the NAc are specifically implicated in the mediation of the avoidance response to aversive olfactory stimuli (formaldehyde) by receiving excitatory glutamatergic input from the medial prefrontal cortex and sending inhibitory signals to the lateral hypothalamic area.
The NAc is also involved in social reward, sexual behaviour, and mating
[47][48][49][125,126,127]. In both male and female mice, approach and investigation of an opposite sex conspecific result in dopamine activation in the NAc, while dopamine signalling in the NAc is sexually dimorphic during sexual behaviours
[50][128]. Additionally, the role of the neuropeptide oxytocin (which is highly implicated in social behaviour) in the NAc has been shown to regulate social preference and other aspects of social behaviour related to sexual behaviour
[51][52][53][129,130,131].
Given the role of the NAc in both sexual behaviour and approach/avoidance, it is likely that it is an important region in mate assessment and avoidance of parasitic risk. The NAc and other regions of the mesolimbic reward network are associated with the determination of the incentive salience and reward value of a potential mate
[54][55][56][38,132,133]. As such, how the cues of infected individuals influence the functioning of the reward network components (specifically dopamine levels) and subsequent approach/avoidance regarding mate choice remains to be determined.
2.5. Concluding Statement on Brain Regions
This short discussion summarizes the brain regions and their interconnections (circuits) shown to be involved in the mediation of responses to infection threats in rodents. Whether these regions and circuits are similarly involved in the initiation and maintenance of avoidance responses remains to be determined. In addition, a range of other brain regions, including those in the social brain network (e.g., medial prefrontal cortex, basal ganglia, ventral tegmental area), are implicated in the processing of defensive responses to a variety of threats, likely including those to the pathogen threat (for a discussion, see Tseng et al., 2023
[57][134]). Notably, the prefrontal cortex has been recently shown to encode both general and specific threat representations
[58][135]. In this regard, studies with humans showed that pictures of sick LPS-treated individuals led to decreased activity in the ventromedial prefrontal cortex and the initiation of avoidance
[59][136].
2.6. Immune–Brain Interactions
The roles of peripheral systems (e.g., immune systems) and their interactions with central systems in the modulation of the behavioural avoidance of the pathogen threat need consideration. Infection- and disease-associated contexts have been shown to trigger immune responses
[60][61][62][63,137,138]. It is likely that social cues of infection can alter immune responses in uninfected individuals. Indeed, recent work has suggested a co-evolution of social behaviour and immune responses. A zinc finger transcription factor, ZFP189, was shown to modulate social behaviour by controlling transposable elements and immune responses in the prefrontal cortex of mice
[63][139]. This was based on earlier work suggesting a co-evolutionary link between social behaviour and anti-pathogen immune responses driven by interferon-γ signalling
[64][140]. ZFP182 upregulated immune genes that are involved in anti-pathogen responses, while also promoting social behaviour. Moreover, these responses were suggested to be responsive to environmental stressors, including possibly that of the parasite/infection threat. It was further speculated that other transcription factors may regulate social behaviour and immune responses. These findings provide an exciting new direction for the elucidation of the neurobiological regulation of pathogen avoidance.
2.7. Microbiome–Immune–Brain Interactions
Animals have diverse populations of bacteria and viruses both internally and externally
[65][66][67][68][141,142,143,144]. There is now substantial evidence that the gastrointestinal microbiota (microbiome) can signal to the brain (microbiota–gut–brain axis) through a range of pathways, including immune activation; production of microbial metabolites and peptides; alterations in neurotransmitters and hormonal levels; neuromodulators and activation of the vagus nerve
[69][145]. These actions can influence various brain regions, including those of the social brain, with marked implications for pathogen avoidance and mate choice.
The microbiome plays a role in influencing the immune response and, subsequently, social behaviour and pathogen avoidance. The microbiome can influence social behaviour through the production of, and effect on, chemosignals
[65][70][71][141,146,147]. Microbial metabolism can release volatile compounds that are detectable by conspecifics and may influence olfactory social signalling
[72][148]. Studies in fruit flies (
Drosophila melanogaster) have shown that the gut microbiota composition mediates olfactory-based mate preference
[73][149]. More pertinent to this review, Li et al. (2013)
[74][150] demonstrated that trimethylamine (TMA), which is a volatile compound involved in the signalling of species identity, is influenced by the microbiota in mice. TMA is produced by gut bacteria during choline metabolism and is converted into an ‘odorless’ compound by enzymes in the liver. However, these liver enzymes are downregulated in sexually mature male mice, allowing TMA and its odour to be detectable in the urine. When male mice were administered antibiotics, they produced significant quantities of TMA, suggesting the direct implication of the microbiome in producing this chemosignal.
Additional studies have shown that the sexual attractiveness of female mice may be impacted by dysbiosis of the gut microbiome
[75][151]. Yi and Cha (2022)
[75][151] found that male mice showed more interaction with control females in a three-chamber social test in comparison to females that had a gut microbiome manipulated by antibiotic treatment. The authors suggested that the decrease in sexual attractiveness in antibiotic-treated females may be due to alterations to the immune system
[75][151]. It is also possible that dysbiosis of females may be influencing the chemosignals through hormonal effects as well. Some research has coined the term ‘Estrogen–Gut Microbiome Axis’, highlighting the influence that the microbiome has on the estrogenic system
[76][77][152,153]. Multiple studies have shown that alterations to the circulating estrogen and androgen levels impact gut microbiota composition
[78][79][80][81][82][83][154,155,156,157,158,159]. Given that estrogens are heavily implicated in olfactory and pheromonal cues, it is likely that reduced female attraction caused by dysbiosis may be a result of altered estrogen levels. Altered estrogen levels may influence the production of other pheromones and ultimately influence the attractiveness of the female
[84][85][160,161].
Additionally, some interesting research has directly investigated the influence of parasitism on the microbiome. These interactions are complex and are mediated by the environment, genetics, and the infection burden
[86][162]. Zhao et al. (2019)
[87][163] demonstrated that female mice infected with the helminth parasite
Schistosoma japonicum had a significant influence on the gut microbiome composition, likely caused by the parasitic egg granulomas in the intestinal tissue. Preliminary data show that the gut microbiome composition is affected by tick parasitism in mice
[88][164]. A number of additional studies in rodents have shown an interaction between parasitic infection and the host microbiome
[87][89][90][91][163,165,166,167].
To date, much of the work on the effect of the microbiome composition and how it influences mate choice has been conducted in non-rodent species
[73][92][93][149,168,169]. This leaves an exciting avenue for rodent researchers to investigate and establish how the microbiome composition (including both bacterial and viral elements), and the impact of parasitism on this composition, may influence mate choice and social behaviour. Integral to this is also determining how the microbiome influences the expression of affective states such as disgust, fear, and anxiety.
3. Neuropeptides and Hormones Involved in Pathogen Avoidance
Many of the brain regions, and likely the microbiome components, considered above involve the actions of various neuromodulatory systems: various neuropeptide systems, neurotransmitter systems, sex steroid hormones and other steroids. These are briefly considered below.
3.1. Oxytocin
The neuropeptide oxytocin is implicated in various aspects of social behaviour, such as the processing of social information and social memory in mice and various other rodent species
[94][95][170,171]. In addition to mediating social recognition and peer affiliation, oxytocin is involved in the olfactory-mediated recognition, and avoidance of, infected individuals. In rodents, oxytocin receptors modulate the ‘social salience network’, a collection of interconnected brain nuclei involved in the processing and encoding of social and sensory cues and involved in the processing of cues associated with the expression of disgust
[95][96][97][98][171,172,173,174].
Female mice with both genetic and pharmacological impairment to oxytocin receptor function displayed decreased aversion and increased choice toward parasitized male conspecific odour compared to control females in a mate-choice paradigm
[99][175]. Additionally in female mice, the preference for parasitized and non-parasitized conspecific male odours was affected by the administration of an oxytocin receptor antagonist, suggesting that oxytocin is involved in the rapid decision-making associated with the approach or avoidance of a parasitic threat
[100][48]. These findings suggest that oxytocin may play a central role in the recognition and subsequent avoidance of parasitized males by females.
A study on conditioned disgust responses in male rats found that oxytocin modulates the expression of socially mediated disgust. When male rats with lithium chloride (LiCl)-induced sickness were conditioned with a social partner, they showed an increased mouth-gaping response (indicative of disgust and malaise) when re-exposed to the conditioned partner in comparison to exposure to a novel conspecific. Boulet et al. (2016)
[101][176] demonstrated that when given an oxytocin receptor antagonist, rats that were conditioned with LiCl showed significantly less disgust (mouth gaping) when re-exposed to the conditioned partner, suggesting that functional oxytocin receptors are required to develop an associative disgust response toward a conspecific. As such, oxytocin may modulate both disgust-associated social salience and social recognition, integral components of mate choice.
What oxytocin-associated pathways are involved in mediating these effects remains to be determined. Also, whether these modulatory roles arise from the effects of oxytocin on other neurotransmitter/neuromodulatory pathways remains to be determined. In this regard, oxytocin has interactions with estrogens and dopamine, and likely immune components, in the regulation of social behaviours
[102][177]. Both dopaminergic and estrogenic systems are implicated in the regulation of female mate choice and, as such, pathogen avoidance.
3.2. Vasopressin
The neuropeptide vasopressin has also been shown to influence the avoidance of pathogen risk. Arakawa et al. (2010)
[25][104] demonstrated that vasopressin within the MeA of rats is involved in conspecific infection avoidance. When exposed to the odours of LPS-treated male conspecifics, the expression of vasopressin messenger RNA in the MeA increased. Infusion of the vasopressin receptor (V1A) antagonist to the MeA inhibited the previously demonstrated avoidance response to LPS-treated odours.
Vasopressin is structurally very similar to oxytocin and has a high homology for oxytocin receptors as well as vasopressin (AVP) receptors. Due to this homology, the vasopressin system may likely interact with, and influence, the oxytocin system and the role that it is known to play in pathogen and sickness detection and avoidance. Similarly, as for oxytocin, the roles of other neurotransmitter/neuromodulatory systems remain to be examined regarding the potential interactions with vasopressin.
3.3. Estrogens
Estrogens are a class of steroid hormones that affect multiple physiological functions and behaviours in both males and females. In females, heightened sexual motivation is mainly influenced by estrogens, which in turn influence dopamine signalling in the mesolimbic reward centres of the brain
[103][104][105][59,178,179]. The salience of a potential mate’s cues can influence the sexual preference and decision made. Lynch and Ryan (2020)
[103][59] postulated that estrogens increase female interest in mating, while the estrogenic influence on dopamine facilitates the detection of signals with high incentive versus low incentive and informs mate preference.
There is also evidence that estrogenic mechanisms are involved in the processing and avoidance of conspecific infection cues
[99][106][107][175,180,181]. Kavaliers et al. (2004)
[99][175] showed that female ERα and ERβ receptor knock-out mice had impairments in the discrimination and avoidance of urine odour in both males that were either treated with LPS or infected with a nematode parasite. This suggests that estrogenic mechanisms are involved in the recognition of, and response to, sick and infected conspecifics. Male ERα and ERβ receptor knock-out mice showed similar impairment in their response to the urine odours of sick and infected males
[108][71]. The inability of ERα and ERβ receptor knock-out mice to recognize cues of infection was shown to not be the result of impaired olfactory processing, as other olfactory-based paradigms were not impaired (such as the ability to distinguish the sexual status of conspecifics and aversion to predator odour).
Additionally, all three estrogen receptors (ERα, ERβ, GPER) are expressed in the MeA, where they are involved in enhancing social recognition and interact with oxytocin receptors, suggesting that estrogens are implicated in the MeA’s role in the detection of and behavioural response to parasitized conspecifics
[106][109][110][180,182,183].
Given the roles that estrogenic mechanisms play in olfactory-mediated social recognition, response to pathogenic threat, influence on immune function, and sexual motivation/behaviour, estrogens are integral to the processing of salient social information that informs the adaptive behavioural response to pathogenic risk
[106][111][112][113][114][180,184,185,186,187]. More research investigating how estrogenic mechanisms interact and potentially link these physiological, cognitive, and behavioural processes is required.
3.4. Progesterone
In human research, the compensatory prophylaxis hypothesis states that the luteal phase of the menstrual cycle (characterized by high progesterone levels) suppresses various mechanisms of immune response and thus should result in a heightened disgust response (reviewed in Fleischman and Fessler, 2011
[115][188]). Recent evidence suggests that progesterone plays a part in the rodent disgust response and pathogen avoidance. In a re-analysis of the results of Kavaliers et al. (2021)
[116][189] conducted by Bressan and Kramer (2022)
[117][190], estrous female mice administered exogenous progesterone had an enhanced avoidance response to the odours of a nematode-infected male.
The biological mechanisms that progesterone may be acting on to influence a disgust-like response remain to be determined. However, progesterone may influence sensory inputs involved in detecting salient social information, as progesterone was shown to potentially influence social recognition
[116][189]. More research into the potentially enhancing effects of progesterone and its neurosteroid products on pathogen detection and disgust-like response needs to be conducted before further speculation.
3.5. Concluding Statement on Neuropeptides and Hormones
The researchers have highlighted the neuromodulatory mechanisms that have been directly implicated in the modulation of pathogen detection and avoidance as they relate to mate choice. There are a range of other regulatory systems that need consideration (e.g., immune system components, opioid systems, neurotransmitters (e.g., 5-HT, endocannabinoids) and steroid and non-steroid hormones). For example, there is evidence that avoidance of infection risk is driven by physiological responses associated with stress and anxiety. Lopes (2023)
[118][191] speculated that after the initial detection of parasite risk, the sympathetic nervous system, adrenomedullary response and adrenocortical response are triggered to facilitate avoidance. This reinforces the need to further consider the roles of, and links between, disgust, fear, and anxiety in modulating pathogen detection and avoidance.