Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Bing Wang and Version 2 by Peter Tang.
A periodontal pathogen, namely, Porphyromonas gingivalis, was found to be associated with all three types of cancers: oral squamous cell carcinoma (OSCC), colorectal cancer (CRC), and pancreatic ductal adenocarcinoma (PDAC).
- periodontitis
- oral squamous cell carcinomas
- colorectal cancers
- pancreatic ductal carcinomas
- bateria–host interactions
- microbial interaction
1. Introduction
The human microbiome consists of a diverse range of bacteria that play a vital role in maintaining the equilibrium between health and diseases [1][2][1,2]. Among them, the oral microbiome stands as the second most diverse and intricate ecosystem [3] residing in the oral cavity, which is the primary site of entrance into both the digestive and respiratory systems. The enlarged Human Oral Microbiome Database reports the presence of over 700 bacterial species involved in dynamic and intricate microbial interactions [4]. The oral microbiome has garnered increasing attention in recent years due to its potential implication for various health/disease conditions, not only inside the oral cavity but also at distant body sites [5][6][5,6].
The significant advancements in next-generation sequencing technology and bioinformatic tools have facilitated the exploration of the harmonious equilibrium among the microbiome, the host, and the environment. It has been revealed that the microbial community as a whole, rather than a few single microbes, maintains this equilibrium. In the healthy state, the microbiome and host establish a symbiotic relationship; whereas in the disease state, a dysbiosis environment promotes the prevalence of pathogenic species in a microbiome, ultimately contributing to the development of illnesses [7]. It was shown that the dysbiotic oral microbiome not only leads to oral infectious diseases, such as caries and periodontitis, but also plays an important role in the development of multiple systemic diseases, including cardiovascular disease, rheumatoid arthritis, Alzheimer’s disease, pulmonary disease, and cancer [8][9][10][11][12][8,9,10,11,12].
Cancer, which is characterized by uncontrolled cell growth and potential metastasis, remains a leading cause of mortality globally [13]. Traditionally, the etiological factors of cancer have been attributed to intrinsic factors, like genetic, environmental, and lifestyle components [14]. Recent discoveries in cancer research revealed that tumor growth might be affected by the dynamic interactions between all constituents inside the tumor microenvironment (TME) as well [15][16][15,16]. Within the TME, there are not only intricate signaling contacts between cellular and non-cellular factors but also reciprocal interactions involving microbial components [16][17][16,17]. It was believed that the microbiome could be operated as a powerful regulator inside the TME, thereby influencing the host (immune) responses [7][18][7,18].
Oral squamous cell carcinoma (OSCC) and gastrointestinal cancers, including colorectal cancer (CRC) and pancreatic ductal adenocarcinoma (PDAC), are among the malignancies that have been linked to the oral microbiome in an intriguing way [19]. It was hypothesized that oral microbes reach distant body sites via the circulatory system after processes such as mastication and routine oral hygiene practices, like teeth brushing and flossing [20]. The concept of the “oral-gut axis” was proposed to illustrate this connection [21]. For example, Fusobacterium nucleatum, which is a well-known Gram-negative pathogen for periodontal infections, has been implicated in the development of systemic disorders, including premature birth, inflammatory bowel disease, and CRC. Another periodontal pathogen, namely, Porphyromonas gingivalis, was found to be associated with all three types of cancers: OSCC, CRC, and PDAC [22][23][22,23]. Until now, ample studies have explored the role of oral microbes in the onset and development of cancers [6][19][24][6,19,24]. However, it is noteworthy that most studies focused on the role of a single bacterial species. It has been acknowledged that tumor tissues do not harbor a single bacterial species, but a multi-species microbial community that accommodates active bacterial interactions [25][26][25,26]. These bacterial interactions could change the formation of tumors. For example, Pustelny et al. demonstrated in a murine tumor model where mice were coinfected with the cystic fibrosis pathogen Pseudomonas aeruginosa and a strictly anaerobic bacterium Veillonella parvula had worse survival rates due to higher P. aeruginosa loads in tumor tissues compared with those coinfected with either bacterial species alone [27]. Lertpirlyapong et al. showed that gastric colonization with multi-species microbiota and the carcinogenic pathogen Helicobacter pylori in male mice leads to more invasive gastrointestinal intraepithelial neoplasia than colonization with H. pylori alone [28]. Although both studies were conducted in mice, the experimental evidence hints at the potential importance of microbial interaction in cancer development. Thus, the existing knowledge on the contribution of a single bacterial species to cancer progression is insufficient without considering the complex dynamics arising from interactions within the microbial community.
The oral pathogen F. nucleatum is known as a bridging bacterium that is able to coaggregate with various bacterial species, such as P. gingivalis, Treponema denticola, and Prevotella intermedia [29][30][29,30]. Similarly, the fimbriae of P. gingivalis can mediate the coaggregation with Streptococcus gordonii, Veillonella sp., and Aggregatibacter actinomycetemcomitans [31][32][31,32]. It was shown that F. nucleatum can enhance the invasion of human gingival epithelial cells by P. gingivalis, which might increase the transmission of P. gingivalis to other body sites in periodontitis patients [33].
2. The Prevalence of P. gingivalis in OSCC, CRC, and PDAC
P. gingivalis is a well-known periodontal pathogen. It is a Gram-negative anaerobic bacterium associated with the onset and progression of periodontitis [34][44]. Previous studies showed that it can colonize malignant tissues in an oral cavity, such as OSCC, ESCC, and gingival carcinoma [35][45]. Sayehmiri et al. conducted a meta-analysis that revealed the presence of P. gingivalis is associated with a risk increase of more than 1.36-fold in the development of OSCC [36][46]. An excessive amount of P. gingivalis was identified as a potential risk factor for OSCC [37][38][39][47,48,49]. In addition to oral cancers, P. gingivalis was frequently related to cancers at other body sites, including esophageal cancer, lung cancer, CRC, and PDAC [40][41][42][43][50,51,52,53]. Ample clinical studies found high abundances of P. gingivalis in both tumor tissue and fecal samples of CRC patients, which were correlated with the onset of CRC and poor prognosis in patients. For example, in a cross-sectional study, Kerdreux et al. examined 247 CRC patients and 89 controls (stages I–IV). They found a significant increase in P. gingivalis levels in fecal samples of CRC patients compared with the healthy controls. P. gingivalis could be identified in the fecal samples of 2.6–5.3% of CRC patients [22]. In another cohort study, P. gingivalis was detectable in 10 out of 31 CRC tissue samples using quantitative polymerase chain reaction (qPCR). A higher prevalence of P. gingivalis was found in individuals with the latter phases of colonic carcinogenesis [44][35]. Similarly, a high prevalence of P. gingivalis in PDAC patients has been reported. A recent prospective cohort study of 361 individuals diagnosed with PDAC found that the presence of P. gingivalis was associated with a 59% rise in PDAC development. Results also show an imbalance in oral microbial composition occurred before the onset of the cancer [45][54]. Another prospective cohort study examined 405 individuals diagnosed with pancreatic cancer, together with 410 control subjects, and found that the individuals with high levels of antibodies against P. gingivalis had a greater than twofold increased risk of developing PDAC [46][55]. Overall, there is clear clinical evidence that a high level of P. gingivalis, either in an oral cavity, fecal samples, or tumor tissues, is associated with the development of all three types of cancers.3. Mechanisms of P. gingivalis in Cancer Development
As a known periodontal pathogen, P. gingivalis employs many strategies to compromise tissue integrity and impair the host immune response. These strategies include the prevention of cell apoptosis, stimulation of cell proliferation, initiation of chronic inflammation, and generation of oncometabolites [47][56]. Previous studies indicate that P. gingivalis can promote tumorigenesis by influencing various signaling pathways (Figure 1): (1) Upon infection of the host by P. gingivalis, the B7-H1 receptor can be activated, facilitating the apoptosis of activated T cells. The increased expression of B7-H1 receptors in host cells may impact the persistence of inflammatory illnesses [48][57]. (2) Nucleoside Diphosphate Kinase (NDK), which is the effector protein produced by intracellular P. gingivalis, can block the signaling of extracellular adenosine triphosphate (ATP)/purinergic receptor (P2X7) on macrophages by consuming ATP. This prevents inflammasome activation and the secretion of interleukin-1β (IL-1β), consequently facilitating the process of tumorigenesis [49][58]. The NDK enzyme is also known to phosphorylate heat shock protein 27 (HSP27), and it is capable of triggering antiapoptotic processes upon phosphorylation [50][59]. (3) P. gingivalis activates antiapoptotic pathways, such as Janus kinase 1 (JAK1)/signal transducer and activator of transcription 3 (STAT3) and phosphoinositide 3-kinase PI3K/protein kinase B (Akt) in oral epithelial cells, thus promoting OSCC [51][60]. Besides inhibiting the intrinsic apoptosis of the invaded epithelial cells, P. gingivalis can also enhance the progression of the S phase of the cell cycle by inhibiting the p53 tumor suppressor gene (TSG) through the FimA adhesin [52][61]. Expression of the aforementioned B7-H1 receptor can inhibit the effector T cells by inducing regulatory T cells, which is beneficial for the invaded cell survival [48][53][57,62]. Due to the induction of regulatory T cells by the B7-H1 receptor, the immune system is (partly) evaded. (4) The activation of extracellular signal-regulated kinase 1/2 (ERK1/2)-protein ETS1, p38/HSP27, and PAR2 (protease-activated receptor 2)/nuclear factor kappa B (NF-κB) pathways was observed in response to P gingivalis infection, leading to the induction of pro-matrix metalloproteinase-9 (pro-MMP-9) expression, hence the increasing levels of MMP-9 and enhanced cellular invasion [54][55][56][63,64,65].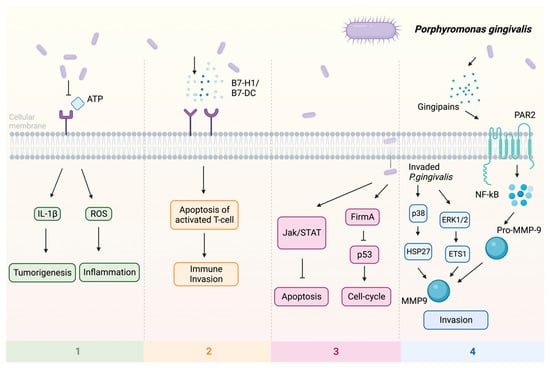
Figure 1. Multiple pathways employed by P. gingivalis in tumor induction: (1) P2X7 activation via ATP is blocked, leading to the stimulation of IL-1β, which promotes tumorigenesis, and the induction of ROS, which fosters a pro-inflammatory microenvironment. (2) Facilitation of immune evasion occurs through the activation of B7-H1 and B7-DC receptors, contributing to a (partial) circumvention of the immune system. Immune evasion is facilitated through the activation of B7-H1 and B7-DC receptors, contributing to a (partial) evasion of the immune system. (3) Activation of FimA results in the downregulation of p53, enhancing the host cell’s cell cycle while simultaneously suppressing apoptosis. The JAK/STAT axis is also implicated in the downregulation of apoptosis. (4) Additionally, P. gingivalis stimulates invasion through PAR2 activation via gingipains, activating NF-κB signaling, which leads to the formation of MMP-9, thereby enhancing P. gingivalis invasion. Upon invasion, pro-MMP-9 undergoes upregulation facilitated by ERK1/2 and ETS1, along with activation of p38 and HSP27. Abbreviations: ATP—adenosine triphosphate; ERK1/2—extracellular signal-regulated kinase 1/2; ETS1—protein; FimA—protein; HSP27—heat shock protein 27; IL-1β—interleukin-1β; JAK—Janus kinase; MMP-9—matrix metalloproteinase-9; NF-κB—nuclear factor kappa B; P2X7—purinergic receptor; pro-MMP-9—pro-matrix metalloproteinase-9; p38, p53—protein; ROS—reaction oxygen species; STAT—signal transducer and activator of transcription.