Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Nafees Khan and Version 2 by Lindsay Dong.
Ethylene, a gaseous phytohormone, is emerging as a central player in the intricate web of plant developmental processes from germination to senescence under optimal and stressed conditions. The presence of ethylene has been noted in different plant parts, including the stems, leaves, flowers, roots, seeds, and fruits.
- ethylene
- plant
- cell division and elongation
- flower and fruit development
- chloroplast development
- photosynthesis
- senescence and abscission
1. Introduction
Plant development is influenced by several intrinsic and extrinsic factors that coordinate to regulate all processes in the life cycles of plants. Plant growth is one carefully regulated process that occurs throughout the vegetative phase. Plant growth regulators act as regulators of plant developmental processes and are crucial for the resilience of responses under stress. Ethylene (ET) is a versatile hormone involved in development, metabolism, and stress responses in plants. Researchers have shown manifold roles of this hormone in the development of plants, as a signaling agent, in leaf development, in senescence, in fruit ripening, in the promotion of germination, etc. ET is the second most fundamental unsaturated hydrocarbon. It exhibits peculiar dose-dependent actions under ideal and external disruptions for all its suppression or promotion responses like germination, ripening, growth, and senescence. For instance, a low concentration of ET facilitates the activation of defense signaling in plants; its high concentration seems to inhibit development in Triticum aestivum and Cucumis sativus [1][2][3][1,2,3]. Plants growing in compacted soil experience excessive accumulation and concentration of ET in root tissues, resulting in root growth inhibition [4]. Increasing evidence confirms that ET is crucial in plant architecture. In Zea mays, ACS7 (1-aminocyclopropane-1-carboxylic acid synthases 7) mutation brought about a dwarf phenotype with a larger leaf angle on Sdw3 (semi-dwarf3) due to increased ET synthesis [5].
ET plays a central role in the growth and development process, starting from seed germination to senescence. However, complex crosstalk between phytohormones like abscisic acid (ABA), gibberellin (GA), cytokinin, and auxin regulates the responses sometimes antagonistically or synergistically [6][7][6,7]. ET biosynthesis or signaling mutant showed differential responses to hormonal sensitivity and plant developmental processes. For example, ethylene overproduction 3 (eto3) and constitutive triple response 1 (ctr1) mutants correspond to ABA insensitivity, whereas ethylene receptor 1 (etr1), ethylene-insensitive 2 (ein2), and ethylene-insensitive 6 (ein6) show enhancement of ABA sensitivity [8]. The faulty hook formation in ET-insensitive mutant ein2 signifies the prominence of ET-mediated auxin biosynthesis for hook development and regulation [9]. Moreover, EIN3 synchronizes the expression of chlorophyll biosynthesis genes PORA/B (PROTOCHLOROPHYLLIDE OXIDOREDUCTASE A/B) [10]: the pigment-binding proteins LHC which are essential for photosynthesis initiation. EIN3/EIL1 chiefly regulates ethylene signaling responses, thereby tuning the range of transcriptional regulation depending on spatiotemporal and environmental conditions [11]. Exogenous application of ET has been found to increase the number of lateral roots, root fresh weight, and mineral content in the root and shoot system, along with the upregulation of auxin biosynthesis and transportation genes [12]. In addition, ET has a significant effect on flower development and sex differentiation. ACS gene family has been widely studied and characterized for its potential function in sex determination [13] Fruit ripening is another ET-dependent process associated with multiple biological events like respiration, pigment accumulation, ET production, change in texture, and overall building up of fruit quality traits. ET activity participates in gene expression responsible for the aforementioned changes during ripening [14].
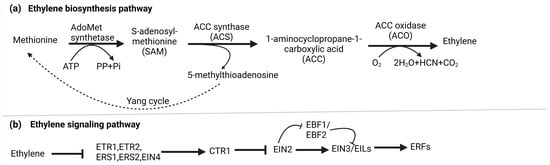
2. Ethylene Biosynthesis and Signaling
Ethylene biosynthesis involves a two-step enzymatic pathway starting with methionine, which is converted into S-adenosyl-L-methionine (SAM or S-AdoMet) by S-AdoMet synthetase. This transformation utilizes one ATP molecule. Subsequently, S-AdoMet is enzymatically converted into 1-aminocyclopropane-1-carboxylic acid (ACC) and 5′-methylthioadenosine (MTA) by ACC synthase (ACS) [15][25]. Lastly, ET is generated through the oxidation of ACC, facilitated by ACC oxidase (ACO). In parallel, the Yang cycle operates to convert MTA, a byproduct of the second stage, back into methionine, ensuring an optimal methionine pool [16][26]. Under normal basal ET production levels, ACS is believed to control the rate-limiting step in biosynthesis [15][25]. Nevertheless, specific circumstances can lead to ACO becoming the limiting factor [17][27]. Figure 1a illustrates the biosynthesis of ET.Figure 1. (a) Biosynthesis of ethylene: In higher plants, ethylene is produced through the Methionine/Yang cycle. This involves converting methionine into 1-aminocyclopropane-1-carboxylic acid (ACC) using ACC synthase (ACS), and then ACC is further converted into ethylene by ACC oxidase (ACO). The Yang cycle recycles 5-methylthioadenosine to regenerate methionine, which is crucial for this process. (b) Signaling pathway of ethylene: Ethylene receptors (ETR1, ERS1, EIN4, ETR2, and ERS2) on the endoplasmic reticulum (ER) activate CTR1 kinase, which phosphorylates and degrades EIN2s C-terminal part, preventing its nucleus entry. Simultaneously, EBF1/2 transcripts produce F-box proteins that target EIN3/EIL transcription factors for degradation, blocking ethylene response. Ethylene binding deactivates CTR1, leading to EIN2 dephosphorylation. A protease cleaves EIN2s C-terminal part (EIN2-CEND) in the nucleus and cytoplasm. EIN2-CEND then enters the nucleus and binds to EIN3/EIL transcription factors, initiating gene expression for ethylene response.
The canonical ethylene signaling pathway begins when ethylene binds to ER-localized receptor proteins, such as ETHYLENE RESPONSE 1 (ETR1), ETHYLENE RESPONSE 2 (ETR2), ETHYLENE-INSENSITIVE 4 (EIN4), ETHYLENE RESPONSE SENSOR 1 (ERS1), and ERS2 in Arabidopsis, organized into two subfamilies defined by their ET binding and histidine kinase domains. Without ET, these receptors activate CONSTITUTIVE TRIPLE RESPONSE (CTR1), a Ser-Thr protein kinase on the ER membrane, which phosphorylates ETHYLENE-INSENSITIVE 2 (EIN2) C-terminal end (EIN2-CEND). F-box proteins ETP1 and ETP2 target phosphorylated EIN2 for degradation by the 26 S proteasome, preventing its nuclear translocation [18][28].
3. Snapshot of Regulation of Ethylene Biosynthesis
Since ACS and ACO are the exclusive enzymes involved in ET biosynthesis, most regulation concerning overall ET production revolves around modulating these pivotal enzymes’ transcription, translation, and protein stability. However, these enzymes are also affected by phytohormones and other stimuli like light and stress. In the early stages of research on the transcriptional regulation of ET biosynthesis, scientists uncovered the presence of a multigene family known as ACS genes, which exhibited distinct expression patterns in plants. Notably, four specific ACS genes in S. lycopersicum were pivotal in orchestrating the shift from autoinhibitory to autocatalytic ET production during fruit ripening [19][32]. Leading the charge as the initial regulator of ACS expression was the MADS-box transcription factor SlRIN, which directly bolstered the expression of selected S. lycopersicum ACS genes [20][33]. Over the years, numerous other transcription factors have emerged as key players, promoters, and inhibitors of ACS gene expression, influencing diverse growth processes. However, there is also supporting information suggesting that ET itself can directly influence ACS transcription. For instance, the ET response factor SlERF2/TERF2 of S. lycopersicum interacts with the promoter of NtACS3, thereby stimulating its expression [21][34]. The regulation of ET synthesis is further complicated by its interaction with light, phytohormones, and various biotic or abiotic stresses. The impact of light on ACS gene expression varies, contingent on developmental stages and light conditions. Notably, different ACS isozymes seem to have specific functions, with ACS5, 6, 8, and 9 primarily influencing ET production in dark-grown seedlings, while ACS2 and ACS4 assume control over ET production in well-lit conditions [22][35]. Mutations in the phytochrome genes PHYA and PHYB have been found to affect ET biosynthesis, with phyA mutants displaying a more pronounced increase in ET production.
Much like their ACS counterparts, ACO genes are subject to transcriptional regulation. Notably, the S. lycopersicum HD-ZIP transcription factor SlHB-1 and the ripening regulator RIN directly stimulate ACO gene expression. Across various species, different classes of transcription factors have also been shown to regulate ACO. Interestingly, ET can directly control ACO expression through intricate feedback mechanisms mediated by ERF (ethylene response factor) proteins [23][36]. In S. lycopersicum, ACO1 experiences upregulation in response to white light pulses. This phenomenon propels ET production and serves as a reporting mechanism for ET responses, thus establishing a positive feedback loop. Additionally, ACO transcript levels surge after ACC treatment of light-grown seedlings. During phases of ripening or stress, when ACS activity is maximum, ACO activity becomes rate-limiting, prompting an upswing in ACO expression. This orchestrated feed-forward mechanism is pivotal in eliminating excess ACC when ACO activity imposes restrictions on ET production [22][35].
Transcriptional regulation of ET biosynthesis is complemented by posttranslational modifications of ACS proteins. ACS proteins share a conserved N terminus and catalytic core, there’s notable variability in the C terminus among different ACS isoforms. This variability leads to the classification of ACS proteins into three major groups in Arabidopsis: type 1 (ACS1, 2 and 6), type 2 (ACS4, 5, 8, 9, and 11), and type 3 (ACS7). Type 1 has a phosphorylation site for mitogen-activated protein kinases (MAPKs) and calcium-dependent protein kinases (CDPKs). Type 2 has a site for CDPKs and ETO1 (Ethylene Overproducer1)/ETO1-LIKE1 (EOL), an E3 ligase leading to degradation. Type 3 has no target site. Phosphorylation stabilizes type 1 ACS, promoting ET production. Conversely, dephosphorylation can lead to ACS degradation, although this effect depends on the type of ACS [23][36]. Moreover, phosphorylation can destabilize type 2 ACS, as exemplified by the phosphorylation of ACS5 (type 2) by Casein Kinase 1.8 (CK1.8), resulting in E3 ubiquitin ligases mediated degradation [24][37]. Light-triggered posttranslational regulation of type-2 ACSs, especially ACS5, regulating hypocotyl elongation during the dark-to-light shift [25][29]. PIF3 plays a key role, with ET stabilizing PIF3 in light. This light-stimulated stabilization of ACS enzymes increases ET production, potentially contributing to PIF3 stability and the subsequent promotion of ET-driven hypocotyl elongation under light exposure [26][38]. PP2A, a regulatory component, plays a role in posttranslational ACS stability regulation. PP2A-mediated dephosphorylation negatively affects ACS6 protein stability in the dark [27][39]. Posttranslational regulation of ACS also involves ubiquitination, with type 3 ACS7 being targeted for degradation through ubiquitination mediated by XBAT32 (XB3 orthologue 2 in Arabidopsis (Figure 2a). Protein Phosphatase 2C family members (PP2C’s) also play a role in stabilizing ACS7 [23][36].
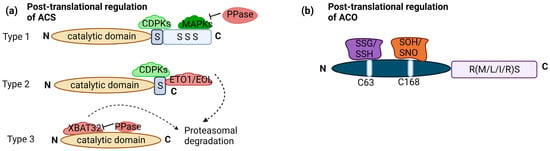
Figure 2. Posttranslational regulation of ACS and ACO: (a) ACS enzymes can be categorized into three groups based on their phosphorylation sites: type 1 with both CDPK (calcium-dependent protein kinase) and MAPK (Mitogen-activated protein kinase) sites, type 2 with only CDPK sites, and type 3 with no C-terminal regulatory sites. Type 1 ACS is positively regulated by MAPKs and CDPKs, significantly boosting its activity and ethylene production. Phosphatases (PPase) are involved in dephosphorylation, negatively impacting protein stability. Type 2 ACSs are regulated by the ETO1-containing E3 ligase, recognizing the TOE (Target of ETO1) domain in their C-termini. CDPK phosphorylation likely plays a role in regulating this ubiquitination process and, thus, the stability of Type 2 ACS protein. XBAT32 directly binds to type 3 ACS, leading to protein degradation, while PPase plays a role in stabilizing. (b) ACO enzymes fall into three related groups based on amino acids in the RXS motif: type 1 with RMS intermediate residue, type 2 with R-L/I-S intermediate residue, and type 3 with RRS intermediate residue. Type 1 ACOs can undergo modifications at C63, including S-glutathionylation (SSG) and S-sulfhydration (SSH), as well as at C168, including S-sulfenylation (SOH) and S-nitrosylation (SNO). No such modifications have been observed in type 2 and type 3 ACOs. The figure is based on and modified from [23][36]. Dotted arrow—multistep pathway; solid arrow—promotion; flat-head arrow—inhibition.
While much is known about the posttranslational regulation of ACS, research on the posttranslational regulation of ACO has been comparatively limited. It is worth mentioning that the three categories of ACO can be distinguished based on the specific intermediate amino acid present within the conserved RXS motif. Type 1 ACOs contain an RMS intermediate residue, type 2 ACOs possess an R-L/I-S intermediate residue, and type 3 ACOs feature an RRS intermediate residue. ACO proteins undergo redox-specific posttranslational modifications. These modifications involve cysteine residues and include S-glutathionylation (SSG), S-sulfhydration (SSH) at cysteine (C63), S-sulfenylation (SOH), and S-nitrosylation (SNO) at C168. These modifications are recorded only in type 1 ACO (Figure 2b).
4. Involvement of Ethylene in Plant Developmental Processes
4.1. Cell Division and Elongation
The role of ET in cell division is complex. Ethylene affects cell division depending on the specific tissue type and the internal and external signals at play. In certain situations, ET acts as a stimulator of cell division. For example, during the development of the apical hook and during the early stages of apical hook development, ethylene appears to play a synergistic role with auxins in stimulating cell division within the subepidermal layers. This collaborative effect is crucial for the bending of the apical hook.
However, the role of ET in cell division is not uniform across all plant tissues. Ethylene’s effects on cell division in the root system are somewhat contradictory. Studies have shown that ET does not significantly alter the expression pattern of CYCLIN-DEPENDENT PROTEIN KINASE B1;1 (CYCB1;1), pointing out that it does not directly affect mitotic activity in the root. However, it is important to note that ET modulates cell division within the quiescent center, developing additional columella layers in the root cap [28][29][44,45].
The impact of ET on leaf cell division is context-dependent. Under environmental stress, particularly when plants experience less than 10 h of osmotic stress, ET mediates transient and reversible cell cycle cessation. This effect is thought to involve the phosphorylation-mediated inactivation of CYCLIN-DEPENDENT KINASE A (CDKA), possibly via the MPK3/6 pathway. Notably, this cell cycle arrest operates independently of the EIN3 transcriptional control [30][49]. Cell cycle inhibition by ET in leaves is multifaceted; ET accumulation activates BOLITA, an ERF, activating type II TCP (TEOSINTE BRANCHED 1/CYCLOIDEA/PCF) genes [31][50]. These TCP proteins then bind to the promoter of RBR1 (RETINOBLASTOMA RELATED 1), phosphorylating E2Fa and repressing E2F target gene transcription, thereby impeding progression into the S-phase and cell division [32][51].
Cellular growth relies on key processes like the rearrangement of the cytoskeleton, the modification of the cell wall facilitated by cell-wall-remodeling enzymes, and water uptake via aquaporins. Cell elongation involves rearranging cortical microtubules (CMTs) perpendicular to the growth axis [33][54]. Ethylene rapidly alters CMT orientation in Arabidopsis roots and hypocotyls, inhibiting cell elongation and promoting radial swelling within 10 min [33][54].