Selenoproteins (SePs) from Se-enriched agricultural foods have attracted increasing attention due to their bioactivities, indicating that Se-containing foods have great potential to be used as natural functional materials for dietary Se supplements. Selenoproteins account for a significant portion of the total Se content in various Se-enriched foods. It can be obtained from plant-based, animal-based sources, and also fungi and yeast sources, which not only provide essential amino acids but also possess physicochemical properties of both Se and proteins. Additionally, Se-containing peptides (SePPs) have also been prepared from Se-enriched plants, such as rice, green tea, soybean, and tuna to explore their potential health benefits. In addition, a variety of factors, including components, amino acid species and sequences, molecular weight, Se status, and structure can significantly affect the bioactivities and functional applications. As several of the SePs identified in mammals are critical selenoenzymes in cells, animal foods that are rich in these SePs are of particular importance. Although Se is not deemed as a crucial element for higher plants, some plants can still integrate it into SePs. Several SePs have been identified in higher plants, including those found in mammalian cells such as GPxs, TrxRs, and selenocysteine methyltransferases. These proteins are also involved in various plant physiology processes, such as antioxidant defense, redox regulation, and Se metabolism.
- selenium
- selenoprotein
- bioaccessibility
- bioactivity
- food resources
1. The Route from Selenoamino Acids to Selenoproteins
2. Functional Properties of Selenoproteins in the Human Body
| Selenoprotein | Name | Specific/Rich in | Main Function(s) | Reference |
|---|---|---|---|---|
| Glutathione Peroxidase (GPx) | GPx1 (or CGPx) |
Nearly all Mammalian Tissues | A family of peroxidases that reduces H2O2, lipid peroxides and organic peroxides from the cytoplasm, cell membrane, and extracellular space of human cells using glutathione as the e− donor in reducing peroxide-induced oxidative stress. | [92,117][24][50] |
| [ | ||||
| 123 | ||||
| , | ||||
| 124 |
| Source | Se Content (μg/g) | Major Se Species | Identification Method | Study Model | Biological Effects | Reference | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Se-enriched Cardamine violifolia | 2450 ± 80 | SeCys, SeMet, MeSeCys | HPLC-AFS LC-MS/MS |
In vitro: antioxidant activity assessment assays (DPPH/OH/O2−·scavenging capacity test) In vivo: ICR mice (male, four-week-old, SPF grade). |
Antioxidant activity and anti-fatigue activity (increase in SOD level, GSH level, and HG level, promotes GPxs activity, suppress MDA and protein carbonyl levels, decrease in BLA and BUN levels). | [76][6] | |||||
| GPx2 (or GI-GPx) |
Gastrointestinal Epithelial Cells (Cytoplasm, Extracellular) | ||||||||||
| Se-enriched Cardamine violifolia | 215–735 | SeMet, MeSeCys, SeCys | HPLC-AFS Nano LC-MS/MS Preparative HPLC |
In vitro: antioxidant activity assessment assays (DPPH/OH/O2−/ABTS+·scavenging capacity test). | Antioxidant activity. | [143][80] | GPx3 (or PGPx) |
Plasma/Intestine (Extracellular, Plasma) | |||
| Se-enriched soybean | 0.33925 | SeMet, SeCys | AFS | In vitro: Caco-2 cell In vivo: BALB/c mice (female, six to eight-week-old,18.0 ± 2.0 g body weight). |
Antioxidant activity: the presence of SeP from soybean inhibited oxidative stress through upregulating the expression of antioxidant enzymes (GPx, SOD) via modulating the NRF-2/HO-1 signaling pathway. Additionally, the administration of soybean SeP to mice improved the activity of GPx and SOD. | [71][81 | GPx4 (or PHGPx) | Sperm (Biological Membrane/Cytomembrane (Phospholipid)) | |||
| ] | |||||||||||
| Se-enriched soybean | 6.35–11.47 | SeMe, SeCys, SeMeCys | AFS, HPLC ICP-MS, FT-IR SEM | / | / | [84][15] | GPx6 | Embryonic Tissue/Epithelial Tissue of Olfactory Organs | |||
| Se-enriched soybean | ~40 | SeMet, SeCys | AFS Q Exactive Orbitrap MS HPLC-MS/MS |
In vitro: Caco-2, HepG2 and Endothelial EA. Hy926 cells. In vivo: ICR mice (female, six-week-old) (D-galactose-induced aging mice). |
Antioxidant activity: protected cells by suppressing the form of TNF-α inflammatory factors and down-regulating the expression levels of cellular adhesion factors. Anti-inflammation and anti-aging: the administration of SeP enhanced SOD and GPx-1, reduced aspartate aminotransferase, amine aminotransferase, and NF-κB, and alleviated brain oxidative damage via modulating MAPK/NF-κB pathway in D-galactose-induced aging in mice. |
[146][84] | Thioredoxin Reductase (TrxR) | TrxR1 | |||
| Se-enriched soybean | Soybean protein isolate: 13.79 ± 0.11 | Extracellular Matrix | Soybean peptides: 21.78 ± 0.17 | Reduces thioredoxin, the e− donor for peroxiredoxin reduction of H | SeCys | 2O2 and other peroxides. Using flavin adenine dinucleotide (FAD) as a coenzyme, it catalyzes the reduction of thioredoxin (Trx) by NADPH. It participates in various cellular processes, including DNA synthesis, protein folding, and cell signaling, and is implicated in several diseases, including cancer and neurodegenerative disorders. | [100,101,118][32][33][51] | ||||
| HPLC-ESI-MS/MS | In vivo: Sprague Dawley rats (male). | Hepatoprotective effects (alleviated liver fibrosis caused by CCL | 4 | by promoting GPxs synthesis and increasing MMP9 mRNA expression). | [ | 73] | TrxR2 | Mitochondria | |||
| Se-MeSeCys, | SeMet | AAS MRM HPLC-ESI-MS/MS |
/ | / | [ | TrxR3 | Testis | ||||
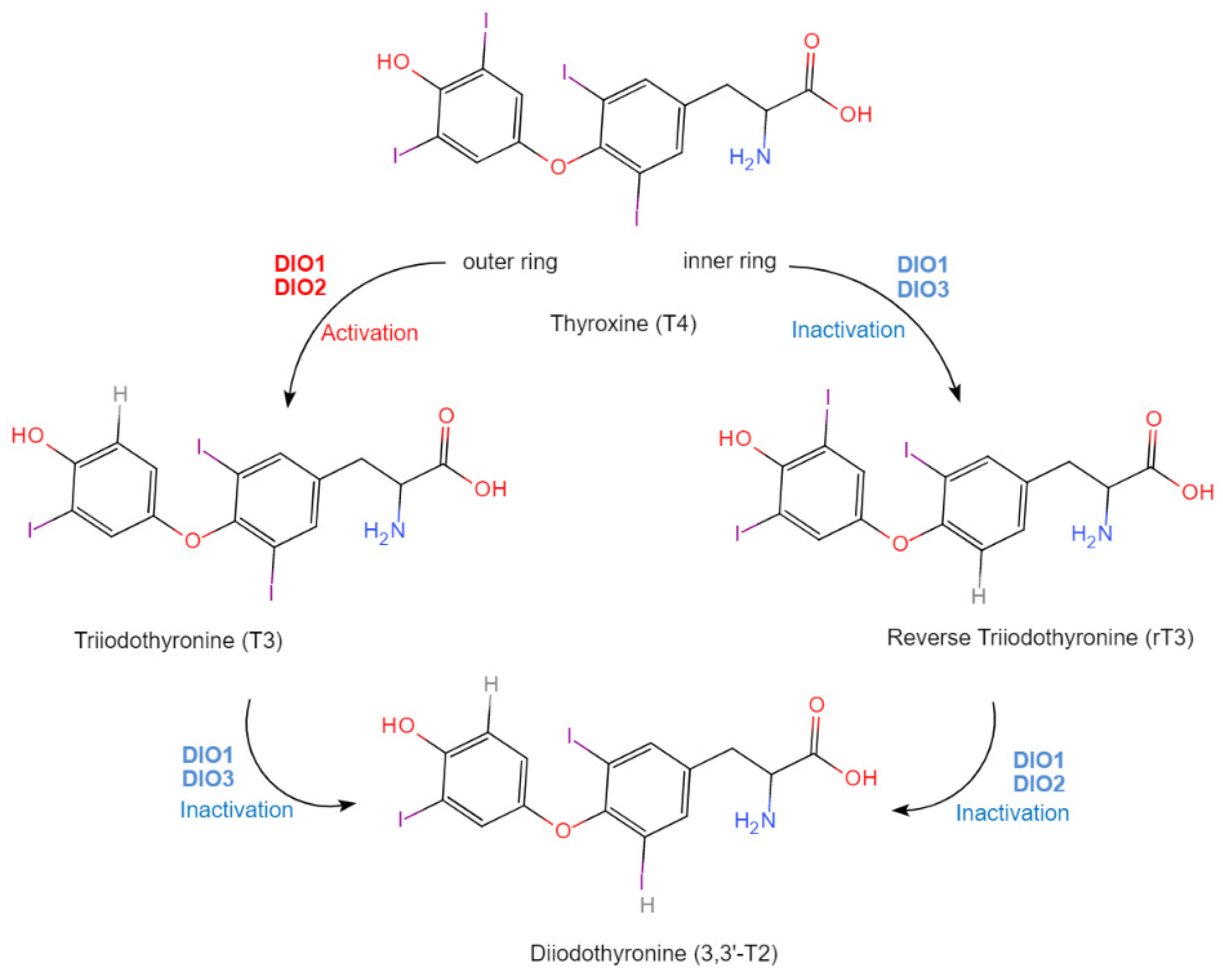
| 145 | ] | [ | Iodothyronine Deiodinase (DIO) | DIO1 | Liver/Kidney | Regulation of thyroid gland secretion, thyroid hormone metabolism, and neuron health. | [53,119][36][52] | ||||
| DIO2 | Pituitary Gland/Skeletal Muscle/Thyroid/Heart/Fat/CNS | ||||||||||
| DIO3 | Brain/Fetal Tissue/Placenta | ||||||||||
| [ | 1 | ] | 83 | ] | |||||||
| Se-enriched soybean | 75 ± 5 | SeMet, SeCys | ICP-MS 2D HPLC-ICP-MS; HPLC-Chip-ESI-ITMS |
/ | / | [144][82] | |||||
| Se-biofortified corn (Zea mays Lin) | 32.37 | SeCys, SeMet, MeSeCys | AFS HPLC-ESI-MS/MS |
In vitro: antioxidant activity assessment assays (DPPH/OH/O2−·scavenging capacity test, inhibition of linoleic acid peroxidation) In vivo: BALB/c mice (male, SPF grade) |
Antioxidant, hepatoprotective (suppressed MDA, improved SOD and GPxs activities, decreased oxidative stress, inhibited hepatic injury). | [75][5] | Selenophosphate synthetase 2 | SEPHS2 | Testes/Liver/Kidney/Brain | Catalyzing the synthesis of selenophosphate from selenide and adenosine triphosphate (ATP), it serves as the selenium donor for selenoprotein and helps maintain proper functioning of selenoproteins. | |
| Se-enriched rice | Water-soluble SeP: 22.01 ± 0.34; alkali-soluble SeP: 8.26 ± 0.40; salt-soluble SeP: 1.67 ± 0.07; alcohol-soluble SeP: 0.073 ± 0.13 | / | AFS | In vitro: antioxidant activity assessment assays (DPPH/OH) In vivo: Kunming mice (male, four-week-old, 20–25 g body weight) |
Antioxidant activity: high free radical (DPPH, OH) scavenging effect. The administration of rice SeP (25 μg/kg/day) enhanced the activities of T-AOC, GPx, SOD, reduced MDA levels, reduces adipocytes, alleviates body weight, liver damage, and the abnormal decrease of the liver coefficient in aging mice. However, the high dose of SeP administration was found to cause hypertoxicity. | [149][87] | |||||
| Se-enriched rice | 12.84 ± 0.05 | SeMet | ICP-MS, RP-UPLC-Triple-TOF MS/MS | In vitro: RAW264.7 cell study | Immunomodulatory activity: the SeP hydrolysate enhanced phagocytosis and proliferation of RAW 264.7 cell and suppressed NO production. However, phagocytosis rate declined when the SeP hydrolysate concentration exceeded 100 μg/mL. | [83][14] | [107,120,121][40][53][54] | ||||
| Selenoprotein methionine sulfoxide reductase B1 | |||||||||||
| Se-enriched brown rice | SeP hydro lysates: 0.156–1.79 | SeMet | AFS, Scide Triple TOF-LC-MS/MS | In vitro: RAW264.7 cell study | Anti-inflammatory: suppressed the production of NO, PGE2, IL-6, IL-1β and TNF-α; inhibited the expression of iNOS and COX-2. | [18][2] | SelR/MsrB1 | Cell Nucleus/Cytoplasm | Maintains cellular redox balance, repairs oxidative damage to proteins, regulates cellular signaling pathways, and regulates cell proliferation and apoptosis. | [112,113][ | |
| Se-enriched rice | / | SeMet | SEC-HPLC HPLC-ICP-MS | 45][46] | |||||||
| In vitro: PC12 cell and RAW264.7 cell study | Protected against Pb | 2 | + | induced apoptosis. | [ | 81][12] | 15-kDa Selenoprotein | Sep15 | Various Tissues and Organs | Is involved in oxidative stress regulation, protein folding, thyroid hormone metabolism, and immune function. | [ |
| Se-enriched brown rice | 6.26 | SeCys, MeSeCys, | 122 | ][55] | |||||||
| SeMet | 2D-LC, HPLC-ICP-MS, ESI FT-ICR MS | In vitro: antioxidant activity determination (DPPH/ABTS + scavenging capacity test, ORAC value, chromium VI-reducing activity, and inhibition activity of linoleic acid emulsion peroxidation) | Antioxidant activity: SeP isolated from brown rice possessed higher ORAC values and free radical scavenging activity than native protein. | [ | 70,148][3][86] | Selenoprotein H | SelH | Brain/Nervous System | Cell cycle regulation. Regulates the activity of the nuclear kernel oxidative enzyme and exhibits potential in cancer prevention.][56 | ||
| Se-fertilized maize, cowpea and groundnut | / | SeMet, SeMeSeCys, SeCys | ] | [57] | |||||||
| Selenoprotein I | |||||||||||
| Se-enriched soybean | 1.118 | ICP-MS, HPLC-ICP-MS | / | / | [ | 158][96] | SelI | Testes | Phospholipid biosynthesis. | [53][36] | |
| Se-enriched peanut | 9.71 | SeMet, SeCys, MeSeCys | ICP-MS, HPLC-ICP-MS | In vitro: AML-12 cell. In vivo: ICR mice (four-week-old) |
Exhibited antioxidant activity: peanut SeP suppressed oxidative stress, reversed cell viability and cell death, inhibited ethanol-induced cytochrome P4502E1 activation, and restored GPx enzyme levels. Ameliorated alcohol-induced liver damage: the administration of peanut SeP reduced oxidative stress through modulating MAPK/NF-κB pathway, regulated lipid metabolism, and minimized liver damage. |
[82][13] | Selenoprotein K | SelK | Endoplasmic Reticulum Membrane | Regulates oxidative stress and endoplasmic reticulum stress. Immunity, inflammation and calcium ion adjustment. Regulates endoplasmic reticulum homeostasis and protein folding. Protects skeletal muscles from damage and is required for satellite cells-mediated myogenic differentiation. | [125,126,127 |
| Se-containing Spirulina platensis | , | 128 | ] | [58][59][ | 0.67–1.99 | ICP-MS60][61] | |||||
| In vitro: antioxidant assessment assay (ABTS+); RAW264.7 cell study | Exhibited antioxidant and anti-inflammatory activities (suppressed inflammatory cytokines, including IL-6, TNF-α, MDA, and IL-1β; decreased the production of NO but promoted the activities of SOD and GPxs). | [ | 151 | ] | [ | 89] | Selenoprotein M | SelM | Brain/Heart/Liver/Kidney/Skeletal Muscle | Maintenance of Ca2+ ions, protein folding, promotion of hypothalamic leptin signaling, and thioredoxin antioxidant activity; overexpression of Sel M; activates Parkin-mediated mitophagy to reduce mitochondrial apoptosis and remove HFD-damaged mitochondria. | [129,130][62 |
| Se-containing Spirulina platensis | ] | [ | 63 | ] | |||||||
| / | / | ICP-AES | In vitro: MC3T3-E1 mouse preosteoblast cells | Prevented mitochondrial dysfunction: balanced the expression of the Bcl-2 family while controlling the opening of the mitochondrial permeability transition pore (MPTP). Additionally, recovered oxidative damage induced by cisplatin. This effect was achieved by inhibiting the excessive generation of reactive oxygen species (ROS) and superoxide anions. Consequently, the process reversed both early and late apoptosis triggered by cisplatin, as it inhibited the cleavage of PARP and the activation of caspases. | [ | 152][90] | Selenoprotein N | SelN | Skeletal Muscle | Growth and development of muscles and protein folding. | [131] |
| Se-enriched Chlorella vulgaris | [ | 64 | / | SeMet, SeCys, MeSeCys | ICP-MS, GC-APCI-HRMS, HPLC-ICP-MS GC, MS] | ||||||
| / | The SeP from | Chlorella vulgaris | has higher bioaccessibility (∼49%) as compared to Se-supplements (∼32%), Se-yeast (∼21%), and Se-foods. | [ | 155 | ][93] | Selenoprotein O | SelO | Brain/Liver/Kidney/Testes | Regulation of redox reactions. | [ |
| Se-containing monkeypot nut seeds (Lecythis minor) | 132 | ] | 4480 ± 22 | SeMet | ICP-MS, ESI-Q-TOF LC–MS/MS[65] | ||||||
| / | / | [ | 74 | ] | [ | 4] | Selenoprotein P | SelP | |||
| Se-enriched mushroom ( | Liver/Plasma | Se carrier. Transportation of Se to brain and other tissues of the body. Protein folding. Prevention of ferroptosis-like cell death and stress-induced nascent granule degradation. | Agaricus blazei) | [133,134,135][66][67][68] | |||||||
| 8.2–26.1 | SeCys, MeSeCys, SeMet | HG-AFS HPLC-MS/MS | / | / | [ | 157][95] | Selenoprotein S | SelS | Plasma/Endoplasmic Reticulum/Immune cells | ||
| Se-enriched mushroom ( | Regulation of inflammation and redox reactions. | Agaricus bisporus) | [ | 136,137][69][70 | SeCys | LC-ESI-MS] | |||||
| In vivo: Sprague Dawley rats (male, 9-week-old). | Antioxidant activities and protection against colorectal cancer (promoted the gene expression of GPx-1 and GPx-2 and enzyme activity of GPx-1 in rat colon). | [ | 78 | , | [ | 8156]][94] | Selenoprotein T | SelT | Brain | Regulation of neuronal function and protection against oxidative stress; regulation of a variety of cellular processes, including calcium signaling, endoplasmic reticulum (ER) stress response, and regulation of protein synthesis. | [138][71] |
| Selenoprotein V | SelV | Testes | Expression of taste, regulation of redox homeostasis, and protection against oxidative stress. | [53][36] | |||||||
| Selenoprotein W | SelW | Mitochondria/Skeletal Muscle/Heart/Brain/Liver/Testes | Oxidative stress regulation, bone remolding and muscle growth. Ensures physiological bone remodeling by preventing hyperactivity of osteoclasts. | [124,[139,57140]][72][73] |
3. Dietary Sources of Selenoproteins
3.1. Preparation and Characterization of Selenoproteins from Foods
3.2. Biological Activity of Selenoproteins from Food
References
- Liu, W.; Hou, T.; Shi, W.; Guo, D.; He, H. Hepatoprotective effects of selenium-biofortified soybean peptides on liver fibrosis induced by tetrachloromethane. J. Funct. Foods 2018, 50, 183–191.
- Feng, M.; Wang, X.; Xiong, H.; Qiu, T.; Sun, Y. Anti-inflammatory effects of three selenium-enriched brown rice protein hydrolysates in LPS-induced RAW264.7 macrophages via NF-κB/MAPKs signaling pathways. J. Funct. Foods 2021, 76, 104320.
- Liu, K.; Du, R.; Chen, F. Antioxidant activities of Se-MPS: A selenopeptide identified from selenized brown rice protein hydrolysates. LWT 2019, 111, 555–560.
- Németh, A.; Dernovics, M. Effective selenium detoxification in the seed proteins of a hyperaccumulator plant: The analysis of selenium-containing proteins of monkeypot nut (Lecythis minor) seeds. JBIC J. Biol. Inorg. Chem. 2015, 20, 23–33.
- Guo, D.; Zhang, Y.; Zhao, J.; He, H.; Hou, T. Selenium-biofortified corn peptides: Attenuating concanavalin A—Induced liver injury and structure characterization. J. Trace Elem. Med. Biol. 2019, 51, 57–64.
- Zhu, S.; Yang, W.; Lin, Y.; Du, C.; Huang, D.; Chen, S.; Yu, T.; Cong, X. Antioxidant and anti-fatigue activities of selenium-enriched peptides isolated from Cardamine violifolia protein hydrolysate. J. Funct. Foods 2021, 79, 104412.
- Ježek, P.; Hlušek, J.; Lošák, T.; Jůzl, M.; Elzner, P.; Kráčmar, S.; Buňka, F.; Martensson, A. Effect of foliar application of selenium on the content of selected amino acids in potato tubers (Solanum tuberosum L.). Plant Soil. Environ. 2011, 57, 315–320.
- Maseko, T.; Callahan, D.L.; Dunshea, F.R.; Doronila, A.; Kolev, S.D.; Ng, K. Chemical characterisation and speciation of organic selenium in cultivated selenium-enriched Agaricus bisporus. Food Chem. 2013, 141, 3681–3687.
- Liu, J.; Luo, G.; Mu, Y.; Hoac, T.; Lundh, T.; Önning, G.; Åkesson, B. Selenoproteins and Selenium Speciation in Food. Selenoproteins Mimics 2012, 183–206.
- White, P.J. Selenium metabolism in plants. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2018, 1862, 2333–2342.
- Thiry, C.; Schneider, Y.-J.; Pussemier, L.; De Temmerman, L.; Ruttens, A. Selenium bioaccessibility and bioavailability in Se-enriched food supplements. Biol. Trace Elem. Res. 2013, 152, 152–160.
- Fang, Y.; Xu, Z.; Shi, Y.; Pei, F.; Yang, W.; Ma, N.; Kimatu, B.M.; Liu, K.; Qiu, W.; Hu, Q. Protection mechanism of Se-containing protein hydrolysates from Se-enriched rice on Pb2+-induced apoptosis in PC12 and RAW264.7 cells. Food Chem. 2017, 219, 391–398.
- Gao, L.; Yuan, J.; Cheng, Y.; Chen, M.; Zhang, G.; Wu, J. Selenomethionine-dominated selenium-enriched peanut protein ameliorates alcohol-induced liver disease in mice by suppressing oxidative stress. Foods 2021, 10, 2979.
- Fang, Y.; Pan, X.; Zhao, E.; Shi, Y.; Shen, X.; Wu, J.; Pei, F.; Hu, Q.; Qiu, W. Isolation and identification of immunomodulatory selenium-containing peptides from selenium-enriched rice protein hydrolysates. Food Chem. 2019, 275, 696–702.
- Deng, X.; Liao, J.; Zhao, Z.; Qin, Y.; Liu, X. Distribution and speciation of selenium in soybean proteins and its effect on protein structure and functionality. Food Chem. 2022, 370, 130982.
- Jin, W.; Yoon, C.; Johnston, T.V.; Ku, S.; Ji, G.E. Production of selenomethionine-enriched Bifidobacterium bifidum BGN4 via sodium selenite biocatalysis. Molecules 2018, 23, 2860.
- Qamar, N.; John, P.; Bhatti, A. Emerging role of selenium in treatment of rheumatoid arthritis: An insight on its antioxidant properties. J. Trace Elem. Med. Biol. 2021, 66, 126737.
- Fairweather-Tait, S.J.; Collings, R.; Hurst, R. Selenium bioavailability: Current knowledge and future research requirements. Am. J. Clin. Nutr. 2010, 91, 1484S–1491S.
- Commans, S.; Böck, A. Selenocysteine inserting tRNAs: An overview. FEMS Microbiol. Rev. 1999, 23, 335–351.
- Bo, A.; Forchhammer, K.; Heider, J.; Baron, C. Selenoprotein synthesis: An expansion of the genetic code. Trends Biochem. Sci. 1991, 16, 463–467.
- Alves, C.S.; Vicentini, R.; Duarte, G.T.; Pinoti, V.F.; Vincentz, M.; Nogueira, F.T. Genome-wide identification and characterization of tRNA-derived RNA fragments in land plants. Plant Mol. Biol. 2017, 93, 35–48.
- Kieliszek, M. Selenium–fascinating microelement, properties and sources in food. Molecules 2019, 24, 1298.
- Esworthy, R.S.; Doroshow, J.H.; Chu, F.-F. The beginning of GPX2 and 30 years later. Free Radic. Biol. Med. 2022, 188, 419–433.
- Pei, J.; Pan, X.; Wei, G.; Hua, Y. Research progress of glutathione peroxidase family (GPX) in redoxidation. Front. Pharmacol. 2023, 14, 1147414.
- Caruso, G.; Grasso, M.; Fidilio, A.; Torrisi, S.A.; Musso, N.; Geraci, F.; Tropea, M.R.; Privitera, A.; Tascedda, F.; Puzzo, D. Antioxidant activity of fluoxetine and vortioxetine in a non-transgenic animal model of Alzheimer’s disease. Front. Pharmacol. 2021, 12, 809541.
- Carducci, F.; Ardiccioni, C.; Fiorini, R.; Vignini, A.; Di Paolo, A.; Alia, S.; Barucca, M.; Biscotti, M.A. The ALA5/ALA6/ALA7 repeat polymorphisms of the glutathione peroxidase-1 (GPx1) gene and autism spectrum disorder. Autism Res. 2022, 15, 215–221.
- Wang, Y.; Cao, P.; Alshwmi, M.; Jiang, N.; Xiao, Z.; Jiang, F.; Gu, J.; Wang, X.; Sun, X.; Li, S. GPX2 suppression of H2O2 stress regulates cervical cancer metastasis and apoptosis via activation of the β-catenin-WNT pathway. OncoTargets Ther. 2019, 12, 6639.
- Nirgude, S.; Choudhary, B. Insights into the role of GPX3, a highly efficient plasma antioxidant, in cancer. Biochem. Pharmacol. 2021, 184, 114365.
- Chang, C.; Worley, B.L.; Phaëton, R.; Hempel, N. Extracellular glutathione peroxidase GPx3 and its role in cancer. Cancers 2020, 12, 2197.
- Forcina, G.C.; Dixon, S.J. GPX4 at the crossroads of lipid homeostasis and ferroptosis. Proteomics 2019, 19, 1800311.
- Ursini, F.; Maiorino, M. Lipid peroxidation and ferroptosis: The role of GSH and GPx4. Free Radic. Biol. Med. 2020, 152, 175–185.
- Jia, J.-J.; Geng, W.-S.; Wang, Z.-Q.; Chen, L.; Zeng, X.-S. The role of thioredoxin system in cancer: Strategy for cancer therapy. Cancer Chemother. Pharmacol. 2019, 84, 453–470.
- Tinkov, A.A.; Bjørklund, G.; Skalny, A.V.; Holmgren, A.; Skalnaya, M.G.; Chirumbolo, S.; Aaseth, J. The role of the thioredoxin/thioredoxin reductase system in the metabolic syndrome: Towards a possible prognostic marker? Cell. Mol. Life Sci. 2018, 75, 1567–1586.
- Valverde-R, C.; Croteau, W.; LaFleur, G.J., Jr.; Orozco, A.; St. Germain, D.L. Cloning and expression of a 5′-iodothyronine deiodinase from the liver of Fundulus heteroclitus. Endocrinology 1997, 138, 642–648.
- Gereben, B.; Zavacki, A.M.; Ribich, S.; Kim, B.W.; Huang, S.A.; Simonides, W.S.; Zeold, A.; Bianco, A.C. Cellular and molecular basis of deiodinase-regulated thyroid hormone signaling. Endocr. Rev. 2008, 29, 898–938.
- Kieliszek, M.; Bano, I. Selenium as an important factor in various disease states: A review. EXCLI J. 2022, 21, 948–966.
- Köhrle, J. The trace element selenium and the thyroid gland. Biochimie 1999, 81, 527–533.
- Köhrle, J.; Gärtner, R. Selenium and thyroid. Best. Pract. Res. Clin. Endocrinol. Metab. 2009, 23, 815–827.
- Bianco, A.C.; Kim, B.W. Deiodinases: Implications of the local control of thyroid hormone action. J. Clin. Investig. 2006, 116, 2571–2579.
- Kryukov, G.V.; Castellano, S.; Novoselov, S.V.; Lobanov, A.V.; Zehtab, O.; Guigó, R.; Gladyshev, V.N. Characterization of mammalian selenoproteomes. Science 2003, 300, 1439–1443.
- Schweizer, U.; Fradejas-Villar, N. Why 21? The significance of selenoproteins for human health revealed by inborn errors of metabolism. FASEB J. 2016, 30, 3669–3681.
- Bang, J.; Kang, D.; Jung, J.; Yoo, T.-J.; Shim, M.S.; Gladyshev, V.N.; Tsuji, P.A.; Hatfield, D.L.; Kim, J.-H.; Lee, B.J. SEPHS1: Its evolution, function and roles in development and diseases. Arch. Biochem. Biophys. 2022, 730, 109426.
- Na, J.; Jung, J.; Bang, J.; Lu, Q.; Carlson, B.A.; Guo, X.; Gladyshev, V.N.; Kim, J.; Hatfield, D.L.; Lee, B.J. Selenophosphate synthetase 1 and its role in redox homeostasis, defense and proliferation. Free Radic. Biol. Med. 2018, 127, 190–197.
- Minich, W.B. Selenium metabolism and biosynthesis of selenoproteins in the human body. Biochemistry 2022, 87, S168–S177.
- Lee, B.C.; Lee, S.-G.; Choo, M.-K.; Kim, J.H.; Lee, H.M.; Kim, S.; Fomenko, D.E.; Kim, H.-Y.; Park, J.M.; Gladyshev, V.N. Selenoprotein MsrB1 promotes anti-inflammatory cytokine gene expression in macrophages and controls immune response in vivo. Sci. Rep. 2017, 7, 5119.
- Tarrago, L.; Kaya, A.; Kim, H.-Y.; Manta, B.; Lee, B.-C.; Gladyshev, V.N. The selenoprotein methionine sulfoxide reductase B1 (MSRB1). Free Radic. Biol. Med. 2022, 191, 228–240.
- Brown, K.M.; Arthur, J. Selenium, selenoproteins and human health: A review. Public Health Nutr. 2001, 4, 593–599.
- Raschke, S.; Ebert, F.; Kipp, A.P.; Kopp, J.; Schwerdtle, T. Selenium homeostasis in human brain cells: Effects of copper (II) and Se species. J. Trace Elem. Med. Biol. 2023, 78, 127149.
- Schomburg, L.; Orho-Melander, M.; Struck, J.; Bergmann, A.; Melander, O. Selenoprotein-P deficiency predicts cardiovascular disease and death. Nutrients 2019, 11, 1852.
- Andrade, I.G.A.; Suano-Souza, F.I.; Fonseca, F.L.A.; Lago, C.S.A.; Sarni, R.O.S. Selenium levels and glutathione peroxidase activity in patients with ataxia-telangiectasia: Association with oxidative stress and lipid status biomarkers. Orphanet J. Rare Dis. 2021, 16, 83.
- Zhang, J.; Zheng, Z.-Q.; Xu, Q.; Li, Y.; Gao, K.; Fang, J. Onopordopicrin from the new genus Shangwua as a novel thioredoxin reductase inhibitor to induce oxidative stress-mediated tumor cell apoptosis. J. Enzym. Inhib. Med. Chem. 2021, 36, 790–801.
- Ogawa-Wong, A.N.; Berry, M.J.; Seale, L.A. Selenium and metabolic disorders: An emphasis on type 2 diabetes risk. Nutrients 2016, 8, 80.
- Manta, B.; Makarova, N.; Mariotti, M. The selenophosphate synthetase family: A review. Free Radic. Biol. Med. 2022, 192, 63–76.
- Kang, D.; Lee, J.; Jung, J.; Carlson, B.A.; Chang, M.J.; Chang, C.B.; Kang, S.-B.; Lee, B.C.; Gladyshev, V.N.; Hatfield, D.L. Selenophosphate synthetase 1 deficiency exacerbates osteoarthritis by dysregulating redox homeostasis. Nat. Commun. 2022, 13, 779.
- Peeler, J.C.; Weerapana, E. Chemical biology approaches to interrogate the selenoproteome. Acc. Chem. Res. 2019, 52, 2832–2840.
- Bertz, M.; Kühn, K.; Koeberle, S.C.; Müller, M.F.; Hoelzer, D.; Thies, K.; Deubel, S.; Thierbach, R.; Kipp, A.P. Selenoprotein H controls cell cycle progression and proliferation of human colorectal cancer cells. Free Radic. Biol. Med. 2018, 127, 98–107.
- Ding, W.; Wang, S.; Gu, J.; Yu, L. Selenium and human nervous system. Chin. Chem. Lett. 2022, 34, 108043.
- Verma, S.; Hoffmann, F.W.; Kumar, M.; Huang, Z.; Roe, K.; Nguyen-Wu, E.; Hashimoto, A.S.; Hoffmann, P.R. Selenoprotein K knockout mice exhibit deficient calcium flux in immune cells and impaired immune responses. J. Immunol. 2011, 186, 2127–2137.
- Wang, S.; Zhao, X.; Liu, Q.; Wang, Y.; Li, S.; Xu, S. Selenoprotein K protects skeletal muscle from damage and is required for satellite cells-mediated myogenic differentiation. Redox Biol. 2022, 50, 102255.
- Li, S.; Kuo, H.-C.D.; Yin, R.; Wu, R.; Liu, X.; Wang, L.; Hudlikar, R.; Peter, R.M.; Kong, A.-N. Epigenetics/epigenomics of triterpenoids in cancer prevention and in health. Biochem. Pharmacol. 2020, 175, 113890.
- Zhang, W.; Sun, X.; Lei, Y.; Liu, X.; Zhang, Y.; Wang, Y.; Lin, H. Roles of selenoprotein K in oxidative stress and endoplasmic reticulum stress under selenium deficiency in chicken liver. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2023, 264, 109504.
- Gong, T.; Hashimoto, A.C.; Sasuclark, A.R.; Khadka, V.S.; Gurary, A.; Pitts, M.W. Selenoprotein M promotes hypothalamic leptin signaling and thioredoxin antioxidant activity. Antioxid. Redox Signal. 2021, 35, 775–787.
- Cai, J.; Huang, J.; Yang, J.; Chen, X.; Zhang, H.; Zhu, Y.; Liu, Q.; Zhang, Z. The protective effect of selenoprotein M on non-alcoholic fatty liver disease: The role of the AMPKα1–MFN2 pathway and Parkin mitophagy. Cell. Mol. Life Sci. 2022, 79, 354.
- Negro, R. Selenium and thyroid autoimmunity. Biol. Targets Ther. 2008, 2, 265.
- Mangiapane, E.; Pessione, A.; Pessione, E. Selenium and selenoproteins: An overview on differrent biological systems. Curr. Protein Pept. Sci. 2018, 19, 725.
- Saito, Y. Selenoprotein P as a significant regulator of pancreatic β cell function. J. Biochem. 2020, 167, 119–124.
- Dominiak, A.; Wilkaniec, A.; Wroczyński, P.; Adamczyk, A. Selenium in the Therapy of Neurological Diseases. Where is it Going? Curr. Neuropharmacol. 2016, 14, 282–299.
- Kitabayashi, N.; Nakao, S.; Mita, Y.; Arisawa, K.; Hoshi, T.; Toyama, T.; Ishii, K.-A.; Takamura, T.; Noguchi, N.; Saito, Y. Role of selenoprotein P expression in the function of pancreatic β cells: Prevention of ferroptosis-like cell death and stress-induced nascent granule degradation. Free Radic. Biol. Med. 2022, 183, 89–103.
- Yang, R.; Liu, Y. Structure, Function, and Nutrition of Selenium-Containing Proteins from Foodstuffs. In Mineral Containing Proteins; Springer: Berlin/Heidelberg, Germany, 2017; pp. 89–116.
- Chi, Q.; Zhang, Q.; Lu, Y.; Zhang, Y.; Xu, S.; Li, S. Roles of selenoprotein S in reactive oxygen species-dependent neutrophil extracellular trap formation induced by selenium-deficient arteritis. Redox Biol. 2021, 44, 102003.
- Pothion, H.; Jehan, C.; Tostivint, H.; Cartier, D.; Bucharles, C.; Falluel-Morel, A.; Boukhzar, L.; Anouar, Y.; Lihrmann, I. Selenoprotein T: An essential oxidoreductase serving as a guardian of endoplasmic reticulum homeostasis. Antioxid. Redox Signal. 2020, 33, 1257–1275.
- Kim, H.; Lee, K.; Kim, J.M.; Kim, M.Y.; Kim, J.-R.; Lee, H.-W.; Chung, Y.W.; Shin, H.-I.; Kim, T.; Park, E.-S. Selenoprotein W ensures physiological bone remodeling by preventing hyperactivity of osteoclasts. Nat. Commun. 2021, 12, 2258.
- Misra, S.; Lee, T.-J.; Sebastian, A.; McGuigan, J.; Liao, C.; Koo, I.; Patterson, A.D.; Rossi, R.M.; Hall, M.A.; Albert, I. Loss of selenoprotein W in murine macrophages alters the hierarchy of selenoprotein expression, redox tone, and mitochondrial functions during inflammation. Redox Biol. 2023, 59, 102571.
- Liang, X.; Xue, Z.; Zheng, Y.; Li, S.; Zhou, L.; Cao, L.; Zou, Y. Selenium supplementation enhanced the expression of selenoproteins in hippocampus and played a neuroprotective role in LPS-induced neuroinflammation. Int. J. Biol. Macromol. 2023, 234, 123740.
- Huertas-Abril, P.V.; Prieto-Álamo, M.-J.; Jurado, J.; García-Barrera, T.; Abril, N. A selenium-enriched diet helps to recover liver function after antibiotic administration in mice. Food Chem. Toxicol. 2023, 171, 113519.
- Gandin, V.; Khalkar, P.; Braude, J.; Fernandes, A.P. Organic selenium compounds as potential chemotherapeutic agents for improved cancer treatment. Free Radic. Biol. Med. 2018, 127, 80–97.
- Jiang, W.; He, S.; Su, D.; Ye, M.; Zeng, Q.; Yuan, Y. Synthesis, characterization of tuna polypeptide selenium nanoparticle, and its immunomodulatory and antioxidant effects in vivo. Food Chem. 2022, 383, 132405.
- Rua, R.M.; Nogales, F.; Carreras, O.; Ojeda, M.L. Selenium, selenoproteins and cancer of the thyroid. J. Trace Elem. Med. Biol. 2023, 76, 127115.
- Jiang, Z.; Chi, J.; Li, H.; Wang, Y.; Liu, W.; Han, B. Effect of chitosan oligosaccharide-conjugated selenium on improving immune function and blocking gastric cancer growth. Eur. J. Pharmacol. 2021, 891, 173673.
- Zhu, S.; Du, C.; Yu, T.; Cong, X.; Liu, Y.; Chen, S.; Li, Y. Antioxidant activity of selenium-enriched peptides from the protein hydrolysate of Cardamine violifolia. J. Food Sci. 2019, 84, 3504–3511.
- Zhao, X.; Gao, J.; Hogenkamp, A.; Knippels, L.M.; Garssen, J.; Bai, J.; Yang, A.; Wu, Y.; Chen, H. Selenium-enriched soy protein has antioxidant potential via modulation of the NRF2-HO1 signaling pathway. Foods 2021, 10, 2542.
- Chan, Q.; Caruso, J.A. A metallomics approach discovers selenium-containing proteins in selenium-enriched soybean. Anal. Bioanal. Chem. 2012, 403, 1311–1321.
- Tie, M.; Li, B.; Zhuang, X.; Han, J.; Liu, L.; Hu, Y.; Li, H. Selenium speciation in soybean by high performance liquid chromatography coupled to electrospray ionization–tandem mass spectrometry (HPLC–ESI–MS/MS). Microchem. J. 2015, 123, 70–75.
- Zhang, X.; He, H.; Xiang, J.; Li, B.; Zhao, M.; Hou, T. Selenium-containing soybean antioxidant peptides: Preparation and comprehensive comparison of different selenium supplements. Food Chem. 2021, 358, 129888.
- Liu, K.; Chen, F.; Zhao, Y.; Gu, Z.; Yang, H. Selenium accumulation in protein fractions during germination of Se-enriched brown rice and molecular weights distribution of Se-containing proteins. Food Chem. 2011, 127, 1526–1531.
- Liu, K.; Zhao, Y.; Chen, F.; Fang, Y. Purification and identification of Se-containing antioxidative peptides from enzymatic hydrolysates of Se-enriched brown rice protein. Food Chem. 2015, 187, 424–430.
- Zeng, R.; Farooq, M.U.; Zhang, G.; Tang, Z.; Zheng, T.; Su, Y.; Hussain, S.; Liang, Y.; Ye, X.; Jia, X. Dissecting the potential of selenoproteins extracted from selenium-enriched rice on physiological, biochemical and anti-ageing effects in vivo. Biol. Trace Elem. Res. 2020, 196, 119–130.
- de Lima, A.B.; de Andrade Vilalta, T.; de Lima Lessa, J.H.; Lopes, G.; Guilherme, L.R.G.; Guerra, M.B.B. Selenium bioaccessibility in rice grains biofortified via soil or foliar application of inorganic Se. J. Food Compos. Anal. 2023, 124, 105652.
- Jiang, P.; Meng, J.; Zhang, L.; Huang, L.; Wei, L.; Bai, Y.; Liu, X.; Li, S. Purification and anti-inflammatory effect of selenium-containing protein fraction from selenium-enriched Spirulina platensis. Food Biosci. 2022, 45, 101469.
- Sun, J.-Y.; Hou, Y.-J.; Fu, X.-Y.; Fu, X.-T.; Ma, J.-K.; Yang, M.-F.; Sun, B.-L.; Fan, C.-D.; Oh, J. Selenium-containing protein from selenium-enriched Spirulina platensis attenuates cisplatin-induced apoptosis in MC3T3-E1 mouse preosteoblast by inhibiting mitochondrial dysfunction and ROS-mediated oxidative damage. Front. Physiol. 2019, 9, 1907.
- Zhang, H.; Chen, T.; Jiang, J.; Wong, Y.-S.; Yang, F.; Zheng, W. Selenium-containing allophycocyanin purified from selenium-enriched Spirulina platensis attenuates AAPH-induced oxidative stress in human erythrocytes through inhibition of ROS generation. J. Agric. Food Chem. 2011, 59, 8683–8690.
- Chen, T.; Wong, Y.-S. In vitro antioxidant and antiproliferative activities of selenium-containing phycocyanin from selenium-enriched Spirulina platensis. J. Agric. Food Chem. 2008, 56, 4352–4358.
- Saurav, K.; Mylenko, M.; Ranglová, K.; Kuta, J.; Ewe, D.; Masojídek, J.; Hrouzek, P. In vitro bioaccessibility of selenoamino acids from selenium (Se)-enriched Chlorella vulgaris biomass in comparison to selenized yeast; a Se-enriched food supplement; and Se-rich foods. Food Chem. 2019, 279, 12–19.
- Maseko, T.; Howell, K.; Dunshea, F.R.; Ng, K. Selenium-enriched Agaricus bisporus increases expression and activity of glutathione peroxidase-1 and expression of glutathione peroxidase-2 in rat colon. Food Chem. 2014, 146, 327–333.
- Hu, Z.; Yao, Y.; Lv, M.; Zhang, Y.; Zhang, L.; Yuan, Y.; Yue, T. Isolation and identification of three water-soluble selenoproteins in Se-enriched Agaricus blazei Murrill. Food Chem. 2021, 344, 128691.
- Muleya, M.; Young, S.D.; Reina, S.V.; Ligowe, I.S.; Broadley, M.R.; Joy, E.J.; Chopera, P.; Bailey, E.H. Selenium speciation and bioaccessibility in Se-fertilised crops of dietary importance in Malawi. J. Food Compos. Anal. 2021, 98, 103841.
- Wu, G.; Liu, F.; Sun, X.; Lin, X.; Zhan, F.; Fu, Z. Preparation of selenium-enriched yeast by re-using discarded Saccharomyces cerevisiae from the beer industry for Se-supplemented fodder applications. Appl. Sci. 2019, 9, 3777.
- Suhajda, A.; Hegoczki, J.; Janzso, B.; Pais, I.; Vereczkey, G. Preparation of selenium yeasts I. Preparation of selenium-enriched Saccharomyces cerevisiae. J. Trace Elem. Med. Biol. 2000, 14, 43–47.
- Zhao, X.; Zhao, Q.; Chen, H.; Xiong, H. Distribution and effects of natural selenium in soybean proteins and its protective role in soybean β-conglycinin (7S globulins) under AAPH-induced oxidative stress. Food Chem. 2019, 272, 201–209.
- Hilal, T.; Killam, B.Y.; Grozdanović, M.; Dobosz-Bartoszek, M.; Loerke, J.; Bürger, J.; Mielke, T.; Copeland, P.R.; Simonović, M.; Spahn, C.M. Structure of the mammalian ribosome as it decodes the selenocysteine UGA codon. Science 2022, 376, 1338–1343.
- Zhang, X.; Yu, Y.; Lei, H.; Cai, Y.; Shen, J.; Zhu, P.; He, Q.; Zhao, M. The Nrf-2/HO-1 signaling axis: A ray of hope in cardiovascular diseases. Cardiol. Res. Pract. 2020, 2020, 5695723.
- Wang, X.; Shen, Z.; Wang, C.; Li, E.; Qin, J.G.; Chen, L. Dietary supplementation of selenium yeast enhances the antioxidant capacity and immune response of juvenile Eriocheir Sinensis under nitrite stress. Fish. Shellfish. Immunol. 2019, 87, 22–31.
- Khalil, H.S.; Mansour, A.T.; Goda, A.M.A.; Omar, E.A. Effect of selenium yeast supplementation on growth performance, feed utilization, lipid profile, liver and intestine histological changes, and economic benefit in meagre, Argyrosomus regius, fingerlings. Aquaculture 2019, 501, 135–143.
- Wang, Y.; Chen, H.; Chang, W.; Chen, R.; Xu, S.; Tao, D. Protective effects of selenium yeast against cadmium-induced necroptosis via inhibition of oxidative stress and MAPK pathway in chicken liver. Ecotoxicol. Environ. Saf. 2020, 206, 111329.