Infection by Brucella species in pregnant animals and humans is associated with an increased risk of abortion, preterm birth, and transmission of the infection to the offspring. The pathogen has a marked tropism for the placenta and the pregnant uterus and has the ability to invade and replicate within cells of the maternal–fetal unit, including trophoblasts and decidual cells. Placentitis is a common finding in infected pregnant animals. Several proinflammatory factors have been found to be increased in both the placenta of Brucella-infected animals and in trophoblasts or decidual cells infected in vitro. As normal pregnancies require an anti-inflammatory placental environment during most of the gestational period, Brucella-induced placentitis is thought to be associated with the obstetric complications of brucellosis.
- Brucella
- placentitis
- abortion
- vertical transmission
- inflammation
1. Introduction
2. Epidemiology of Brucella-Induced Pregnancy Complications
In their preferential hosts, Brucella spp. mainly affect the reproductive tract. In bovine and caprine brucellosis, abortion is one of the most characteristic clinical signs, which occurs in the middle to late stages of gestation [9] at a rate that varies from 30 to 80% in susceptible herds. Also, in dogs, the classic symptom of B. canis infection is late abortion, between 30 and 57 days of gestation, with a higher frequency noted between days 45 and 55 [10]. On the other hand, in swine brucellosis, abortion is generally a minor disease presentation under field conditions, as infection could result in small fetuses expelled in placental tissues that are rarely detected by farmers. The first evidence of early abortions may be a return to estrus at 40–45 days after natural breeding [11]. Regarding B. ovis infection, it rarely causes abortions in ewes and rarely extends from one pregnancy to the next [12][13] (Table 1).| Brucella Species | Hosts | Gestational Manifestations | Vertical Transmission | Contagion Source |
|---|---|---|---|---|
| B. melitensis | Small ruminants | Abortion, weak offspring, reduced milk yields | + | Contaminated placenta or aborted fetus. Milk |
| B. abortus | Bovines | Abortion, weak offspring, reduced milk yields | + | Contaminated placenta or aborted fetus. Milk |
| B. suis (biovars 1, 2, 3) | Swine | Abortion, weak offspring | + | Contaminated placenta or aborted fetus. Milk. Contaminated semen |
| B. canis | Canines | Abortion, weak offspring | + | Contaminated placenta or aborted fetus. Milk. Contaminated semen |
| B. ovis | Sheep | Abortion, weak offspring (rare) | Not reported | Close contact or mating with infected rams. |
| B. melitensis, B. abortus, B. suis | Humans | Abortion, preterm birth, intrauterine fetal death, neonatal or maternal death | + | Contaminated milk and dairy products. Tissues or secretions from infected animals. Contaminated aerosols. |
Although abortion in animals caused by Brucella infection is well known, for several years, there has been controversy about the relationship between human brucellosis and pregnancy outcomes. In pregnant women, the seroprevalence of brucellosis varies from 1.3 to 12.2% [14][15][16][17][18]. Such variability depends on livestock contact, as pregnant women without animal-related occupations show a lower seroprevalence of brucellosis [19] than women from agro-pastoral communities, who do most of the work associated with the care and harvest of livestock products [20]. In the last decades, there have been more reports of adverse outcomes in Brucella-infected pregnant women [15][21]. Spontaneous miscarriage rates range from 18.6 to 73.3% [14][22]. Potential factors affecting the rate of miscarriage include the infecting species (B. melitensis is usually regarded as more virulent), the infection route (food versus other sources), and the median age of the mothers. Interestingly, most of the cases are documented to occur during the first and second trimesters of gestation and differ from the time of abortion occurrence in animals, commonly manifested at later gestational stages [23].
3. Brucella Vaccines and Gestational Complications in Animals
In many developing countries, brucellosis leads to substantial economic losses due to abortions and infertility in pregnant livestock. To prevent infections and abortions, which not only result in economic setbacks but also contribute to the spread of the bacterium within herds and pose a risk of human infection, it is recommended to vaccinate animals alongside implementing testing and slaughter measures. Currently, licensed available vaccines include live attenuated strains, such as B. abortus S19 (S19) and B. abortus RB51 (RB51) for bovines and B. melitensis Rev.1 (Rev.1) for small ruminants. There are no vaccines available for dogs and pigs. The Rev. 1 strain is widely used worldwide. When administered to sexually immature females, the vaccine is safe and induces long-lasting protection against B. melitensis infection and abortion [24]. However, administering Rev. 1 during gestation results in a variable abortion rate of 40% to 80%, which can spread disease within the herd and pose a risk to individuals handling aborted placentas and fetuses, as this vaccine strain can cause disease in humans [24][25][26][27]. B. abortus S19 is a naturally attenuated strain with a deletion in the erythritol catabolic genes [28], whereas RB51 is a rough mutant strain derived from B. abortus 2308, which lacks the wboA gene encoding a glycosyl transferase necessary for O-side chain synthesis [29]. Although numerous studies have shown that S19 and RB51 vaccinations protect approximately 65–75% of cows against abortion and infection [30], their administration during pregnancy can cause abortion in cows. Additionally, despite being attenuated in animals, both vaccine strains are infectious to humans [31]. Due to the drawbacks associated with the vaccines mentioned above, several studies have been directed toward the development of new vaccines that are both safe and effective. One strategy involves the creation of subunit vaccines derived from Brucella (including lipopolysaccharides, proteins, DNA, and outer membrane vesicles). Another approach includes the use of new live attenuated mutant strains capable of protecting against virulent Brucella infection, yet without the adverse effects associated with commercial attenuated vaccines [27].4. Pathological Findings in the Infected Placenta
Brucella is known to invade and colonize the placenta of both wildlife and domestic animals, with similar pathological findings in all cases [32][33][34][35][36][37]. Since the early studies by Payne [38][39] until the present, different animal models have been tested to explain the pathology of abortion linked to Brucella infections, which allowed for reaching a consensus regarding the histopathological findings in the placentas of the different animal species. The most frequent lesions include placentitis, inflammatory infiltrates (including polymorphonuclear cells, lymphocytes, and macrophages), vasculitis, necrosis, and ulcerated or compromised chorioallantoic membrane [40][41][42][43][44][45][46][47][48]. Other more sporadic findings are placental calcifications (associated with chronicity), purulent or fibrinous exudates, granulomas, and placental edema. Placental lesions found in animal models of Brucella infection are in line with those found in natural infections. However, the inflammation induced by B. abortus in the murine model is much weaker than the severe inflammation seen in the natural host [49]. In contrast, B. melitensis produces the same type of lesions in mice, sheep, and goats [50]. Whatever the case, the mouse is widely used as a model of abortion and placentitis induced by different Brucella strains [51].5. Brucella Infection and Replication in Placental Cells
Placental infection can originate via two routes: one via sexual transmission, where the pathogen ascends through the genital tract to the placenta, and the other via maternal blood. In humans, there are two points of contact between mother and fetus that could allow transmission of infection: (a) the maternal decidua cells (immune and stroma cells) that come into contact with the extravillous trophoblast (EVT) at the site of implantation and (b) the maternal blood surrounding the syncytiotrophoblast (SYN) [52]. In the face of pathogen entry, there are four main barriers that prevent fetal infection: (1) immune cells present at the maternal–fetal interface, originating from maternal blood, (2) the SYN, a monolayer of multinucleated fused trophoblasts that have intrinsic resistance to infection by certain pathogens, (3) the EVT, which has innate defense mechanisms against pathogen invasion, although it is more susceptible than the SYN, and (4) the basal membrane beneath the trophoblast, representing the last barrier preventing colonization of fetal stroma [52]. The intracellular replication capability of Brucella is a fundamental determinant of its pathogenicity both in general and in gestational complications in particular. Within both phagocytic and non-phagocytic cells, the Brucella-containing vacuole (BCV) engages in transient interactions with early endosomes, late endosomes, and lysosomes [53]. During this phase, BCVs are identified by lysosomal membrane-associated protein 1 (LAMP1). The acidification of BCVs is essential as it promotes the intracellular expression of genes responsible for encoding the VirB type IV secretion system (T4SS). Subsequently, Brucella orchestrates fusion with endoplasmic reticulum (ER) membranes in a VirB T4SS-dependent manner and replicates within ER-derived compartments in both professional and non-professional phagocytes [54][55]. The three main zoonotic species, B. melitensis, B. abortus, and B. suis, are able to infect and replicate in human cell lines of cytotrophoblast (CTB) (BeWo and JAR) and EVT (HTR8/SVNeo, JEG-3 and Swan-71) in vitro [7][50][51]. In CTB, B. abortus and B. suis replicate through the formation of their conventional BCVs. In contrast, in the EVT cell line JEG-3, both strains replicate by forming inclusions in a different vacuole (LAMP1+ and calnexin-). Such replication is not fully T4SS-dependent, whereas B. melitensis replicates in these cells in conventional BCVs in a VirB-dependent manner. These findings are consistent with those observed in trophoblasts isolated from human placentas at term where B. abortus is able to replicate both in CTB (in conventional BCVs) and EVT (also forming inclusions) [56]. Hormone secretion is essential for placental development. In vitro infection with B. abortus and B. melitensis does not affect human chorionic gonadotrophin secretion in JEG-3 cells. However, B. melitensis infection decreases progesterone and estradiol production in these cells [56][57]. Consistent with these results, B. abortus infection has been shown to suppress placental progesterone production in the mouse pregnancy model [8]. In hemochorial placentation (humans, mice), the invasion of the maternal endometrium by EVT typically occurs in the early stages of pregnancy and is a critical step in anchoring the placenta. Furthermore, as the transition from the first to the second trimester occurs, EVT plays an additional role in remodeling uterine arteries, facilitating maternal blood flow into the placental intervillous space. This, in turn, ensures the delivery of essential nutrients and oxygen to the developing fetus. It has been observed that infection with B. melitensis, but not B. suis or B. abortus, diminishes the invasiveness of JEG-3 cells [56]. This reduction in invasiveness could potentially impact implantation and the adequate supply of nutrients and oxygen to the developing fetus. The failure of trophoblast functionality is not due to a cytotoxic effect of Brucella since infection with virulent species did not affect human trophoblast viability [7][56][57]. Of note, however, B. melitensis infection of JEG-3 cells increased the expression levels of CD98hc, a protein involved in the regulation of integrin-mediated signaling, and the authors hypothesized that this change may have a role in the reduced invasiveness of the infected EVT [57].6. Brucella-Induced Inflammatory Responses in Trophoblasts and Other Cells from the Maternal—Fetal Unit
As mentioned above, several studies have shown that Brucella infection induces placentitis in pregnant animals. In addition, histological analyses indicate that trophoblasts are a central target of the pathogen, and studies performed with trophoblastic cell lines revealed the ability of Brucella to manipulate cellular mechanisms to promote its survival and replication in these cells. Of note, several studies have suggested that infected trophoblasts may have an important role in the Brucella-induced inflammatory phenomena in the placenta. Fernandez et al. [7] showed that B. abortus infection significantly increases the production of interleukin 8 (IL-8), monocyte chemotactic protein 1 (MCP-1), GM-CSF, and IL-6 in the human trophoblastic cell line Swan-71. Taking into account that, during Brucella infections, placental trophoblasts could interact with decidual macrophages or with monocytes and neutrophils attracted to the infection site by chemokines, the authors also analyzed the production of proinflammatory factors in the context of the interaction of trophoblasts with infected phagocytes. Of note, the stimulation of Swan-71 cells with conditioned medium (CM) from B. abortus-infected human monocytes (THP-1 cells), macrophages, or neutrophils induced a significant increase of IL-8, MCP-1, and IL-6 compared to stimulation with CM from non-infected cells. Neutralization studies showed that IL-1β is involved in the stimulating effects of CM from infected phagocytes on the production of the three cytokines by Swan-71 cells, whereas TNF-α is also involved in the induction of MCP-1. A similar proinflammatory response has been shown for canine trophoblasts [58]. Primary canine trophoblasts isolated from the placenta of healthy pregnant bitches responded to B. canis infection with increased levels of IL-8 and RANTES (CCL5). Similar to the situation with human trophoblasts, the stimulation of canine trophoblasts with CM from B. canis-infected monocytes and neutrophils also induced a significant increase of IL-8, IL-6, and RANTES secretion compared to stimulation with control CM. While not formally tested in this study, the fact that TNF-α levels were significantly increased in CM from B. canis-infected canine neutrophils and monocytes suggests that this factor may be involved in the stimulating effect of phagocytes on trophoblast cytokines. As IL-8 is a chemoattractant for neutrophils and RANTES is a chemoattractant for a variety of leukocytes in inflammatory sites, these results suggest that trophoblasts-derived chemokines may be involved in the development of the neutrophilic and histiocytic infiltrates usually observed in the placentas of B. canis-induced canine abortions [36][37][59]. Bovine trophoblasts also respond to Brucella infection with an increased production of proinflammatory cytokines. When explants of chroioallantoic membranes obtained from healthy cows were infected on their trophoblastic side with B. abortus, a reduced expression of some proinflammatory genes was observed at 4 h post-inoculation by microarray and RT-PCR [44]. However, this seemed to be a transient phenomenon, as the expression of CXCL8 and CXCL6 was significantly increased at 12 h post-infection (last time measured). A few additional studies have been performed to identify bacterial or host factors involved in the inflammatory response of trophoblasts to Brucella infection. A study by Liu et al. [60] investigated the role of high-mobility group box 1 (HMGB1) in regulating the inflammatory response of primary murine trophoblasts to B. melitensis infection. HMGB1, which is present in all cell types, is a damage-associated molecular pattern (DAMP) and is a known mediator of inflammatory response during sterile and infection-associated diseases. Brucella outer membrane protein Omp25 has been claimed to induce cytokine responses in HPT-8 cells. However, the differences in the levels of the three evaluated cytokines (TNF-α, IL-1β, and IL-10) between cells infected with B. abortus S2308 or its ΔOmp25 mutant were quite small. Nevertheless, the mutant induced a markedly weaker stimulation of p38, ERK1/2, and JNK kinases [61]. Besides trophoblasts, endometrial cells may also be relevant for the induction of inflammatory responses in the maternal–fetal unit. When the blastocyst initiates its implantation in the uterus, trophoblasts begin to invade the endometrial epithelium and the underlying stroma. Stromal cells respond by producing the decidual reaction (epithelial transformation of fibroblasts with glycogen and lipids storage), and this endometrial region transforms into the decidua. The decidual stromal cells secrete prolactin, insulin growth factor-binding protein, and several cytokines that regulate innate immunity [62]. The maternal decidua may be the initial site of placental colonization for Brucella, as has been described for several microorganisms that reach the placenta by the hematogenous route [52][63]. A study by Zavattieri et al.[64] evaluated the ability of B. abortus to invade and establish a replicative niche in non-decidualized and decidualized human endometrial stromal cells (T-HESC cell line). B. abortus was able to infect and replicate in T-HESC cells in both conditions. The production of prolactin by infected decidualized T-HESC did not differ from that of uninfected controls, showing that B. abortus infection does not affect the decidualization status of the cells. Both decidualized and non-decidualized cells increased their production of CXCL-8 (IL-8) and MCP-1 in response to infection. In the context of Brucella infection in the pregnant uterus, endometrial cells may be stimulated by factors secreted by adjacent infected macrophages. Stimulation with conditioned media from B. abortus-infected macrophages induced a significant production of IL-6, MCP-1 and IL-8 by decidualized T-HESC cells. Globally, the results suggest that during B. abortus infection in pregnant females, endometrial cells may produce proinflammatory factors not only in response to bacterial antigens but also to stimulation by factors produced by adjacent Brucella-infected macrophages. These proinflammatory responses and cellular interactions may be long-lasting due to the ability of Brucella to survive and replicate in macrophages and endometrial cells, and may contribute to the gestational complications of brucellosis.7. Conclusions
Abortions associated with Brucella infections have been well documented, not only in domestic animals but also in wildlife, and there is currently a wide consensus regarding the relationship between brucellosis and gestational complications in humans. In many cases, the association with human abortions derives from serological studies in cohorts of pregnant women, but in selected cases, the pathogen has been isolated from placental and fetal tissues, thus confirming the link between the infection and the obstetric complications. While B. melitensis is known to be responsible for the majority of human cases of abortion linked to brucellosis, B. abortus has also been identified as the causative agent in some cases. Placental inflammation has been a common finding in affected animals and probably explains most of the pathology in Brucella-induced abortion and preterm birth, as a successful gestation requires the maintenance of an anti-inflammatory environment in the maternal–fetal unit during most of the pregnancy period. Several studies have shown the ability of different Brucella strains to invade and replicate in human and animal trophoblasts, and in vitro studies have shown that these cells produce a wide array of proinflammatory factors in response to the infection, including TNF-α, IL-6, RANTES, MCP-1, and IL-8. These cytokines may mediate several processes that are deleterious for pregnancy (Figure 1), including the infiltration of neutrophils and macrophages (with increased production of reactive oxygen species, proteases, and other harmful products), the induction of matrix metalloproteinases, and the alteration of the hormonal balance required to support gestation (e.g., decreased production of prolactin and hCG).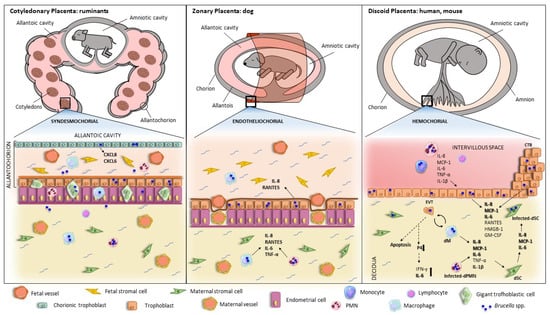
References
- Pappas, G.; Papadimitriou, P.; Akritidis, N.; Christou, L.; Tsianos, E.V. The New Global Map of Human Brucellosis. Lancet Infect. Dis. 2006, 6, 91–99.
- Laine, C.G.; Johnson, V.E.; Scott, H.M.; Arenas-Gamboa, A.M. Global Estimate of Human Brucellosis Incidence. Emerg. Infect. Dis. 2023, 29, 1789–1797.
- Bosilkovski, M.; Arapović, J.; Keramat, F. Human Brucellosis in Pregnancy—An Overview. Bosn. J. Basic Med. Sci. 2020, 20, 415–422.
- Mor, G.; Aldo, P.; Alvero, A.B. The Unique Immunological and Microbial Aspects of Pregnancy. Nat. Rev. Immunol. 2017, 17, 469–482.
- Krishnan, L.; Nguyen, T.; Mccomb, S. From Mice to Women: The Conundrum of Immunity to Infection during Pregnancy. J. Reprod. Immunol. 2013, 97, 62–73.
- Tsai, A.Y.; Byndloss, M.X.; Seyffert, N.; Winter, M.G.; Young, B.M.; Tsolis, R.M. Tumor Necrosis Factor Alpha Contributes to Inflammatory Pathology in the Placenta during Brucella abortus Infection. Infect. Immun. 2022, 90, e00013-22.
- Fernández, A.G.; Ferrero, M.C.; Hielpos, M.S.; Fossati, C.A.; Baldi, P.C. Proinflammatory Response of Human Trophoblastic Cells to Brucella abortus Infection and upon Interactions with Infected Phagocytes. Biol. Reprod. 2016, 94, 131706.
- Ren, J.; Hou, H.; Zhao, W.; Wang, J.; Peng, Q. Administration of Exogenous Progesterone Protects Against Brucella abortus Infection-Induced Inflammation in Pregnant Mice. J. Infect. Dis. 2021, 224, 532–543.
- Rossetti, C.A.; Maurizio, E.; Rossi, U.A. Comparative Review of Brucellosis in Small Domestic Ruminants. Front. Vet. Sci. 2022, 9, 7671.
- Wanke, M.M. Canine Brucellosis. Anim. Reprod. Sci. 2004, 82–83, 195–207.
- Olsen, S.; Tatum, F. Swine Brucellosis: Current Perspectives. Vet. Med. 2016, 8, 1–12.
- Grilló, M.J.; Marín, C.M.; Barberán, M.; Blasco, J.M. Experimental Brucella ovis Infection in Pregnant Ewes. Vet. Rec. 1999, 144, 555–558.
- Ris, D.R. The Bacteriology and Serology of Ewes Inoculated with Viable Brucella ovis Organisms. N. Z. Vet. J. 1970, 18, 2–7.
- Khan, M.Y.; Mah, M.W.; Memish, Z.A. Brucellosis in Pregnant Women. Clin. Infect. Dis. 2001, 32, 1172–1177.
- Elshamy, M.; Ahmed, A.I. The Effects of Maternal Brucellosis on Pregnancy Outcome. J. Infect. Dev. Ctries. 2008, 2, 230–234.
- Kurdoglu, M.; Adali, E.; Kurdoglu, Z.; Karahocagil, M.K.; Kolusari, A.; Yildizhan, R.; Kucukaydin, Z.; Sahin, H.G.; Kamaci, M.; Akdeniz, H. Brucellosis in Pregnancy: A 6-Year Clinical Analysis. Arch. Gynecol. Obstet. 2010, 281, 201–206.
- Makala, R.; Majigo, M.V.; Bwire, G.M.; Kibwana, U.; Mirambo, M.M.; Joachim, A. Seroprevalence of Brucella Infection and Associated Factors among Pregnant Women Receiving Antenatal Care around Human, Wildlife and Livestock Interface in Ngorongoro Ecosystem, Northern Tanzania. A Cross-Sectional Study. BMC Infect. Dis. 2020, 20, 152.
- Ali, S.; Akhter, S.; Neubauer, H.; Scherag, A.; Kesselmeier, M.; Melzer, F.; Khan, I.; El-Adawy, H.; Azam, A.; Qadeer, S.; et al. Brucellosis in Pregnant Women from Pakistan: An Observational Study. BMC Infect. Dis. 2016, 16, 468.
- Kledmanee, K.; Liabsuetrakul, T.; Sretrirutchai, S. Seropositivities against Brucellosis, Coxiellosis, and Toxoplasmosis and Associated Factors in Pregnant Women with Adverse Pregnancy Outcomes: A Cross-Sectional Study. PLoS ONE 2019, 14, e0216652.
- Te-Chaniyom, T.; Geater, A.F.; Kongkaew, W.; Chethanond, U.; Chongsuvivatwong, V. Goat Farm Management and Brucella Serological Test among Goat Keepers and Livestock Officers, 2011–2012, Nakhon Si Thammarat Province, Southern Thailand. One Health 2016, 2, 126–130.
- Vilchez, G.; Espinoza, M.; D’Onadio, G.; Saona, P.; Gotuzzo, E. Brucellosis in Pregnancy: Clinical Aspects and Obstetric Outcomes. Int. J. Infect. Dis. 2015, 38, 95–100.
- Al-Tawfiq, A.J.; Memish, A.Z. Pregnancy Associated Brucellosis. Recent Pat. Anti-Infect. Drug Discov. 2013, 8, 47–50.
- Poester, F.P.; Samartino, L.E.; Santos, R.I. Pathogenesis and Pathobiology of Brucellosis in Livestock. Rev. Sci. Tech. 2013, 32, 105–115.
- Alton, G.G. Control of Brucella melitensis Infection in Sheep and Goats-a Review. Trop. Anim. Health Prod. 1987, 19, 65–74.
- Zundel, E.; Verger, J.M.; Grayon, M.; Michel, R. Conjunctival Vaccination of Pregnant Ewes and Goats with Brucella melitensis Rev 1 Vaccine: Safety and Serological Responses. Ann. Rech. Vet. 1992, 23, 177–188.
- Jiménez de Bagués, M.P.; Marin, C.M.; Barberán, M.; Blasco, J.M. Responses of Ewes to B. Melitensis Rev1 Vaccine Administered by Subcutaneous or Conjunctival Routes at Different Stages of Pregnancy. Ann. Rech. Vet. 1989, 20, 205–213.
- Hensel, M.E.; Garcia-Gonzalez, D.G.; Chaki, S.P.; Hartwig, A.; Gordy, P.W.; Bowen, R.; Ficht, T.A.; Arenas-Gamboa, A.M. Vaccine Candidate Brucella melitensis 16M ΔvjbR Is Safe in a Pregnant Sheep Model and Confers Protection. mSphere 2020, 5, e00120-20.
- Sangari, F.J.; García-Lobo, J.M.; Agüero, J. The Brucella abortus Vaccine Strain B19 Carries a Deletion in the Erythritol Catabolic Genes. FEMS Microbiol. Lett. 1994, 121, 337–342.
- Schurig, G.G.; Roop, R.M.; Bagchi, T.; Boyle, S.; Buhrman, D.; Sriranganathan, N. Biological Properties of RB51; A Stable Rough Strain of Brucella abortus. Vet. Microbiol. 1991, 28, 171–188.
- de Oliveira, M.M.; Pereira, C.R.; de Oliveira, I.R.C.; Godfroid, J.; Lage, A.P.; Dorneles, E.M.S. Efficacy of Brucella abortus S19 and RB51 Vaccine Strains: A Systematic Review and Meta-Analysis. Transbound. Emerg. Dis. 2022, 69, e32–e51.
- Schurig, G.G.; Sriranganathan, N.; Corbel, M.J. Brucellosis Vaccines: Past, Present and Future. Vet. Microbiol. 2002, 90, 479–496.
- Kudi, A.C.; Kalla, D.J.U.; Kudi, M.C.; Kapio, G.I. Brucellosis in Camels. J. Arid Environ. 1997, 37, 413–417.
- Miller, W.G.; Adams, L.G.; Ficht, T.A.; Cheville, N.F.; Payeur, J.P.; Harley, D.R.; House, C.; Ridgway, S.H. Brucella-Induced Abortions and Infection in Bottlenose Dolphins (Tursiops truncatus). J. Zoo Wildl. Med. 1999, 30, 100–110.
- Mackie, J.T.; Blyde, D.; Harris, L.; Roe, W.D.; Keyburn, A.L. Brucellosis Associated with Stillbirth in a Bottlenose Dolphin in Australia. Aust. Vet. J. 2020, 98, 92–95.
- Rebollada-Merino, A.; García-Seco, T.; Pérez-Sancho, M.; Domínguez, L.; Rodríguez-Bertos, A. Histopathologic and Immunohistochemical Findings in the Placentas and Fetuses of Domestic Swine Naturally Infected with Brucella suis Biovar 2. J. Vet. Diagn. Investig. 2023, 35, 258–265.
- Carmichael, L.E.; Kenney, R.M. Canine Abortion Caused by Brucella canis. J. Am. Vet. Med. Assoc. 1968, 152, 605–616.
- Gyuranecz, M.; Szeredi, L.; Rónai, Z.; Dénes, B.; Dencso, L.; Dán, Á.; Pálmai, N.; Hauser, Z.; Lami, E.; Makrai, L.; et al. Detection of Brucella canis-Induced Reproductive Diseases in a Kennel. J. Vet. Diagn. Investig. 2011, 23, 143–147.
- Payne, J.M. Changes in the Rat Placenta and Foetus Following Experimental Infection with Various Species of Bacteria. J. Pathol. Bacteriol. 1958, 75, 367–385.
- Payne, J.M. The Pathogenesis of Experimental Brucellosis in the Pregnant Cow. J. Pathol. Bacteriol. 1959, 78, 447–463.
- Anderson, T.D.; Meador, V.P.; Cheville, N.F. Pathogenesis of Placentitis in the Goat Inoculated with Brucella abortus. I. Gross and Histologic Lesions. Vet. Pathol. 1986, 23, 219–226.
- Siddiqur, R.M.; Kirl, B.B. Clinical and Pathological Findings in Experimental Brucellosis in Pregnant Rats. J. Infect. Dev. Ctries. 2008, 2, 226–229.
- Anderson, T.D.; Cheville, N.F. Ultrastructural Morphometric Analysis of Brucella abortus-Infected Trophoblasts in Experimental Placentitis. Bacterial Replication Occurs in Rough Endoplasmic Reticulum. Am. J. Pathol. 1986, 124, 226.
- Meador, V.P.; Deyoe, B.L. Intracellular Localization of Brucella abortus in Bovine Placenta. Vet. Pathol. 1989, 26, 513–515.
- Carvalho Neta, A.V.; Stynen, A.P.R.; Paixão, T.A.; Miranda, K.L.; Silva, F.L.; Roux, C.M.; Tsolis, R.M.; Everts, R.E.; Lewin, H.A.; Adams, L.G.; et al. Modulation of the Bovine Trophoblastic Innate Immune Response by Brucella abortus. Infect. Immun. 2008, 76, 1897–1907.
- Carvalho Neta, A.V.; Mol, J.P.S.; Xavier, M.N.; Paixão, T.A.; Lage, A.P.; Santos, R.L. Pathogenesis of Bovine Brucellosis. Vet. J. 2010, 184, 146–155.
- Bosseray, N. Brucella Infection and Immunity in Placenta. Ann. l’Institut Pasteur. Microbiol. 1987, 138, 110–113.
- Bosseray, N. Colonization of Mouse Placentas by Brucella abortus Inoculated during Pregnancy. Br. J. Exp. Pathol. 1980, 61, 361–368.
- Poveda-Urkixo, I.; Ramírez, G.A.; Grilló, M.J. Kinetics of Placental Infection by Different Smooth Brucella Strains in Mice. Pathogens 2022, 11, 279.
- Keestra-Gounder, A.M.; Byndloss, M.X.; Seyffert, N.; Young, B.M.; Chávez-Arroyo, A.; Tsai, A.Y.; Cevallos, S.A.; Winter, M.G.; Pham, O.H.; Tiffany, C.R.; et al. NOD1 and NOD2 Signalling Links ER Stress with Inflammation. Nature 2016, 532, 394–397.
- Deng, Y.; Liu, X.; Duan, K.; Peng, Q. Research Progress on Brucellosis. Curr. Med. Chem. 2018, 26, 5598–5608.
- Santos, R.L.; Silva, T.M.A.; Costa, E.A.; Paixo, T.A.; Tsolis, R.M. Laboratory Animal Models for Brucellosis Research. J. Biomed. Biotechnol. 2011, 2011, 518323.
- Robbins, J.R.; Bakardjiev, A.I. Pathogens and the Placental Fortress. Curr. Opin. Microbiol. 2012, 15, 36–43.
- Starr, T.; Ng, T.W.; Wehrly, T.D.; Knodler, L.A.; Celli, J. Brucella Intracellular Replication Requires Trafficking through the Late Endosomal/Lysosomal Compartment. Traffic 2008, 9, 678–694.
- Comerci, D.J.; Martínez-Lorenzo, M.J.; Sieira, R.; Gorvel, J.P.; Ugalde, R.A. Essential Role of the VirB Machinery in the Maturation of the Brucella abortus-Containing Vacuole. Cell. Microbiol. 2001, 3, 159–168.
- Delrue, R.M.; Martinez-Lorenzo, M.; Lestrate, P.; Danese, I.; Bielarz, V.; Mertens, P.; De Bolle, X.; Tibor, A.; Gorvel, J.P.; Letesson, J.J. Identification of Brucella Spp. Genes Involved in Intracellular Trafficking. Cell. Microbiol. 2001, 3, 487–497.
- Salcedo, S.P.; Chevrier, N.; Lacerda, T.L.S.; Ben Amara, A.; Gerart, S.; Gorvel, V.A.; De Chastellier, C.; Blasco, J.M.; Mege, J.L.; Gorvel, J.P. Pathogenic Brucellae Replicate in Human Trophoblasts. J. Infect. Dis. 2013, 207, 1075–1083.
- García-Méndez, K.B.; Hielpos, S.M.; Soler-Llorens, P.F.; Arce-Gorvel, V.; Hale, C.; Gorvel, J.P.; O’Callaghan, D.; Keriel, A. Infection by Brucella melitensis or Brucella papionis Modifies Essential Physiological Functions of Human Trophoblasts. Cell. Microbiol. 2019, 21, 13019.
- Fernández, A.G.; Hielpos, M.S.; Ferrero, M.C.; Fossati, C.A.; Baldi, P.C. Proinflammatory Response of Canine Trophoblasts to Brucella canis Infection. PLoS ONE 2017, 12, e0186561.
- Brennan, S.J.; Ngeleka, M.; Philibert, H.M.; Forbes, L.B.; Allen, A.L. Canine Brucellosis in a Saskatchewan Kennel. Can. Vet. J. 2008, 49, 703–708.
- Liu, X.; Zhou, M.; Wu, J.; Wang, J.; Peng, Q. HMGB1 Release from Trophoblasts Contributes to Inflammation during Brucella melitensis Infection. Cell. Microbiol. 2019, 21, 13080.
- Zhang, J.; Zhang, Y.; Li, Z.; Liu, J.; Shao, X.; Wu, C.; Wang, Y.; Wang, K.; Li, T.; Liu, L.; et al. Outer Membrane Protein 25 of Brucella Activates Mitogen-Activated Protein Kinase Signal Pathway in Human Trophoblast Cells. Front. Vet. Sci. 2017, 4, 197.
- Dunn, C.L.; Kelly, R.W.; Critchley, H.O.D. Decidualization of the Human Endometrial Stromal Cell: An Enigmatic Transformation. Reprod. Biomed. Online 2003, 7, 151–161.
- Vigliani, M.B.; Bakardjiev, A.I. Intracellular Organisms as Placental Invaders. Fetal Matern. Med. Rev. 2014, 25, 332–338.
- Zavattieri, L.; Ferrero, M.C.; Alonso Paiva, I.M.; Sotelo, A.D.; Canellada, A.M.; Baldi, P.C. Brucella Abortus Pro-liferates in Decidualized and Non-Decidualized Human Endometrial Cells Inducing a Proinflammatory Response. Pathogens 2020, 9, 369.