Oilseed crops are rich in plant lipids that not only provide essential fatty acids for the human diet but also play important roles as major sources of biofuels and indispensable raw materials for the chemical industry. The regulation of lipid metabolism genes is a major factor affecting oil production.
- oilseed crops
- lipid metabolism
- seed oil content
1. Introduction
Function of Lipids
Lipids play many roles in plant cells. For example, polar glycerides are the major components of cell membranes, where they act as protective barriers against external damage and initiate signaling [1][6]. Lipid composition varies widely among different species and tissues [2][7]. For example, plants accumulate large amounts of lipids in their seeds or fruits to provide the energy needed for germinating seeds, maintaining moisture, and preventing cold cracking and frostbite in the seeds. Lipids play essential roles in cell signal transduction between flowers, leaves, stems, and other tissues [3][8] and are deposited as waxes on the plant surface to decrease non-stomatal plant water loss or ultraviolet (UV) damage, thus increasing plant tolerance to abiotic stress [4][9].Classification of Plant Lipids
Lipids are widely distributed in all plant tissues and can be divided into eight types according to their chemical composition: FAs, glycerol lipids, glycerophospholipids, sphingolipids, sterol lipids, allyl alcohol lipids, glycolipids, and polyketides [5][6][11,12] (Figure 1). FAs are the building blocks of many of the more complex lipids. About 40 FAs serve as the major components of natural lipids, among over 10,000 known fatty acyl molecules (https://www.lipidmaps.org) accessed on 22 September 2018 [5][7][11,13]. When supplied with sufficient oxygen, FAs can be oxidized and broken down into CO2 and H2O, releasing large amounts of energy, thus making them a major source of energy. FAs can be further divided into saturated fatty acids (SFAs) and unsaturated fatty acids (UFAs) based on the absence or presence of double bonds. In most plants, the major SFAs are palmitic acid (16:0, 16 is the number of carbon atoms, 0 represents the number of double bonds) and stearic acid (18:0), and the major UFAs are monounsaturated FAs (MUFAs), such as oleic acid (18:1), and polyunsaturated FAs (PUFAs), such as linoleic acid (LA, 18:2) and α-linolenic acid (α-ALA, 18:3) [8][14]. These five FAs account for approximately 90% of the lipids in commercial vegetable oils on the market [9][15].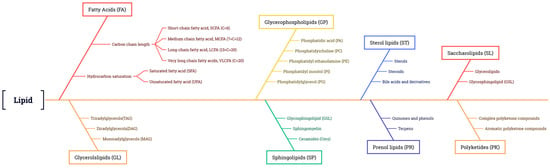
Lipid Accumulation and Storage in Plants
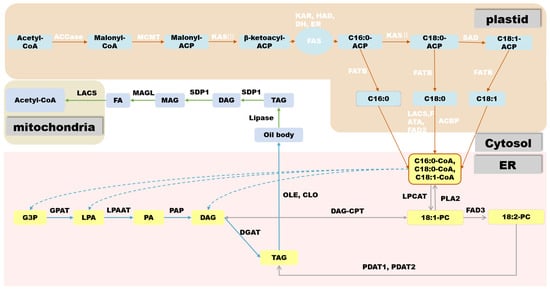
In oilseed crops, the first step in FA biosynthesis is the conversion of acetyl-CoA produced via sucrose glycolysis to malonyl-CoA by acetyl-CoA carboxylase (ACCase). ACCase is a type I biotin-containing enzyme and there are two main forms of ACCase in plants: Heteromeric ACCaseI and Homomeric ACCaseII. ACCase is composed of four subunits: biotin carboxylase (BC), biotin carboxyl carrier protein (BCCP), α-carboxyltransferase (α-CT), and β-carboxyltransferase (β-CT). It has three functional domains, namely the BC functional domain, the BCCP functional domain, and the CT functional domain, respectively [10][11]. After its biosynthesis, malonyl-CoA is transferred to the ACP component of the fatty acid synthase (FAS) complex by malonyl-CoA:ACP malonyltransacylase (MCMT). In plants, FAS is a multi-component type-II enzyme located in the plastids, consisting of 3-β-ketoacyl-ACP synthase III (KASIII), β-ketoacyl-ACP synthase (KASI), ketoacyl-ACP reductase (KAR), hydroxyacyl-ACP reductase (HAD), and enoyl-ACP reductase (ENR). FAS uses acetyl-CoA as the starting unit for a condensation reaction. Each elongation cycle is supplied with a two-carbon unit by malonyl-ACP to produce 16:0-ACP and 18:0-ACP after seven or eight cycles, respectively, at which point 18:0-ACP passes through Δ9 stearoyl-ACP desaturase to form 18:1-ACP, making phosphatidic acid (PA) (16:0) and oleic acid (18:1) the major products of FA biosynthesis in most plant plastids [12].
In most oilseed crops, such as soybean (
Lipid Accumulation and Storage in Plants
In oilseed crops, the first step in FA biosynthesis is the conversion of acetyl-CoA produced via sucrose glycolysis to malonyl-CoA by acetyl-CoA carboxylase (ACCase). ACCase is a type I biotin-containing enzyme and there are two main forms of ACCase in plants: Heteromeric ACCaseI and Homomeric ACCaseII. ACCase is composed of four subunits: biotin carboxylase (BC), biotin carboxyl carrier protein (BCCP), α-carboxyltransferase (α-CT), and β-carboxyltransferase (β-CT). It has three functional domains, namely the BC functional domain, the BCCP functional domain, and the CT functional domain, respectively [26,27]. After its biosynthesis, malonyl-CoA is transferred to the ACP component of the fatty acid synthase (FAS) complex by malonyl-CoA:ACP malonyltransacylase (MCMT). In plants, FAS is a multi-component type-II enzyme located in the plastids, consisting of 3-β-ketoacyl-ACP synthase III (KASIII), β-ketoacyl-ACP synthase (KASI), ketoacyl-ACP reductase (KAR), hydroxyacyl-ACP reductase (HAD), and enoyl-ACP reductase (ENR). FAS uses acetyl-CoA as the starting unit for a condensation reaction. Each elongation cycle is supplied with a two-carbon unit by malonyl-ACP to produce 16:0-ACP and 18:0-ACP after seven or eight cycles, respectively, at which point 18:0-ACP passes through Δ9 stearoyl-ACP desaturase to form 18:1-ACP, making phosphatidic acid (PA) (16:0) and oleic acid (18:1) the major products of FA biosynthesis in most plant plastids [28].
In most oilseed crops, such as soybean (
Glycine max
) and rapeseed (
Brassica napus), TAGs are stored in seeds as an energy source for germination and aid in seed dispersal [13]. TAG consists of a glycerol backbone and three FA molecules chemically linked by ester bonds, providing a carbon skeleton and energy source. TAG biosynthesis and accumulation occur through a complex network of reactions taking place in the plastid, cytoplasm, and ER. Depending on the plant species, TAG can accumulate in different organs, mainly in embryonic tissues (rapeseed) or endosperm tissues (castor bean). In oilseeds that store oil in the embryo, the main storage tissue is the cotyledons, but substantial seed oil can also accumulate in the hypocotyl, radicle, and surrounding endosperm/aleurone layers [14]. After acyl-CoA is transported from the plastid to the ER, the most common pathway for TAG biosynthesis is the acyl-CoA-dependent Kennedy pathway [15]. In this pathway, acyl-CoA is incorporated into glycerol-3-phosphate (G3P) by acyl-CoA:glycerol-3-phosphate acyltransferase (GPAT) and lysophosphatidic acid acyltransferase (LPAT) at the sn-1 and sn-2 positions of G3P, respectively, to form PA.
Cytoplasmic lipid droplets (LDs) are organelles that store non-polar lipids such as TAGs and sterol esters [16]. In mature seeds, LDs are distributed in the central region of storage cells and are mostly oval or irregular in shape [17]. The primary LD structure consists of a phospholipid monolayer coated with various proteins. The current general model of LD biogenesis is that non-poplar lipids such as TAGs are first produced by membrane-associated enzymes in the ER and then accumulate in the form of a lens between lobes of the ER membrane, culminating with LD formation on the cytoplasmic side of the ER membrane [18]. Two important proteins have recently been shown to be involved in LD formation: SEIPIN [19][20] and lipid-droplet-associated protein in Arabidopsis (
), TAGs are stored in seeds as an energy source for germination and aid in seed dispersal [29]. TAG consists of a glycerol backbone and three FA molecules chemically linked by ester bonds, providing a carbon skeleton and energy source. TAG biosynthesis and accumulation occur through a complex network of reactions taking place in the plastid, cytoplasm, and ER. Depending on the plant species, TAG can accumulate in different organs, mainly in embryonic tissues (rapeseed) or endosperm tissues (castor bean). In oilseeds that store oil in the embryo, the main storage tissue is the cotyledons, but substantial seed oil can also accumulate in the hypocotyl, radicle, and surrounding endosperm/aleurone layers [30]. After acyl-CoA is transported from the plastid to the ER, the most common pathway for TAG biosynthesis is the acyl-CoA-dependent Kennedy pathway [31]. In this pathway, acyl-CoA is incorporated into glycerol-3-phosphate (G3P) by acyl-CoA:glycerol-3-phosphate acyltransferase (GPAT) and lysophosphatidic acid acyltransferase (LPAT) at the sn-1 and sn-2 positions of G3P, respectively, to form PA.
Cytoplasmic lipid droplets (LDs) are organelles that store non-polar lipids such as TAGs and sterol esters [33]. In mature seeds, LDs are distributed in the central region of storage cells and are mostly oval or irregular in shape [34]. The primary LD structure consists of a phospholipid monolayer coated with various proteins. The current general model of LD biogenesis is that non-poplar lipids such as TAGs are first produced by membrane-associated enzymes in the ER and then accumulate in the form of a lens between lobes of the ER membrane, culminating with LD formation on the cytoplasmic side of the ER membrane [35]. Two important proteins have recently been shown to be involved in LD formation: SEIPIN [36,37] and lipid-droplet-associated protein in Arabidopsis (
Arabidopsis thaliana) [21].
) [38].
2. Progress in the Identification of Key Genes behind Lipid Metabolism in Plants
2.1. Identification and Functional Characterization of Key Genes
3.1. Identification and Functional Characterization of Key Genes
As mentioned above, FATB is a key enzyme in FA biosynthesis that catalyzes the removal of ACP from 16:0-ACP and 18:1-ACP to generate NEFAs. The knockdown of GmFATB1 expression decreases UFA content in soybean seeds [27][45]. Similarly, the expression of BnFATB in rapeseed appears to be highly and positively correlated with seed oil content (SOC), as evidenced by an analysis integrating quantitative trait loci (QTLs) from nine different populations [28][46]. LPAT is one of the key enzymes in the Kennedy pathway that converts acyl-CoA to PA. The heterologous expression of a yeast (Saccharomyces cerevisiae) sn-2 acyltransferase gene in rapeseed resulted in increased FA content by 8–22% [29][47]. Cloning and spatiotemporal expression of AhLPAT2 from peanut (Arachis hypogaea) showed that increased expression of this gene was closely related to seed oil content; moreover, heterologous expression of AhLPAT2 in Arabidopsis from a seed-specific promoter significantly increased SOC [30][48]. DGAT is a key enzyme that acylates DAG at the sn-3 position. Seed-specific overexpression of Arabidopsis DGAT in wild-type plants resulted in an increase in SOC and seed weight from 29% to 35%, along with an increase in the average thousand-seed weight from 19 mg in non-transgenic plants to 23 mg in the overexpression lines [31][49], which was consistent with the results of heterologous expression of DGAT1 from sesame (Sesamum indicum) in Arabidopsis [32][50]. GDSL esterase is a type of hydrolase that is widely involved in various physiological activities in plants and is important in oil metabolism. The name of the enzyme derives from its conserved domain (GDSL), where G, D, and S represent glycine (Gly), aspartate (Asp), and serine (Ser), respectively [33][52]. Arabidopsis has five GDSL-like lipase genes (also called SEED FATTY ACID REDUCER [SFAR] genes) that decrease SOC by acting downstream of the gibberellin signaling (GA) pathway since GA may affect the seed FA content via SFARs [34][53]. The overexpression or knockout of each SFAR gene significantly lowered or increased SOC, respectively, and altered FA composition. The heterologous expression of a GDSL gene from oil palm (Elaeis guineensis) in Arabidopsis resulted in a 9.5% increase in total FA content compared to the wild type. In addition to these important pathway enzymes and their encoding genes directly involved in de novo FA biosynthesis and TAG assembly, recent studies have identified several genes annotated as participating in carbon source provisioning and photosynthetic pathways that may also be involved in regulating SOC [35][55]. For instance, trehalose-6-phosphate (T6P), a metabolic precursor of sucrose and a key signaling molecule for plants to respond to carbon availability and regulate growth and development, also regulates FA biosynthesis by inhibiting sucrose non-fermenting 1-related kinase 1 (SnRK1). Indeed, incubation of rapeseed suspension cells in T6P-containing medium or heterologous expression of T6P synthase from Escherichia coli in Nicotiana benthamiana significantly increased FA biosynthesis rates [36][56]. CALCINEURIN B-LIKE PROTEIN-INTERACTING PROTEIN KINASEs (CIPKs) are a family of energy-signaling protein kinases in plants. Relative to non-transgenic plants, the overexpression of BnCIPK9 during seed development in transgenic rapeseed lowered oil biosynthesis [37][16].2.2. Transcription Factors Involved in Regulation
3.2. Transcription Factors Involved in Regulation
Looking for transcription factors (TFs) that regulate the expression of multiple genes is also an effective method to increase SOC [38][57]. Currently, well-studied TFs in plants mainly include WRINKLED1 (WRI1), LEAFY COTYLEDON1 (LEC1), LEC2, LEC1-LIKE (LIL), FUSCA3 (FUS3), and ABSCISIC ACID INSENSITIVE3 (ABI3). WRI1 is an APETALA2 (AP2)/ETHYLENE-RESPONSIVE ELEMENT BINDING PROTEIN (EREBP) that regulates genes involved in glycolysis and sucrose entry into TAG [39][58]. WRI1 was discovered in Arabidopsis in 1998 as a mutant affecting seed storage accumulation. Compared with the wild type, the mutant was unable to convert glucose and sucrose into precursors for fatty acid synthesis during seed development and reduced the activity of several glycolytic enzymes such as hexokinase and phosphofructokinase, resulting in an 80% reduction in SOC [40][59]. The oil content of N. benthamiana leaves expressing WRI1 from castor bean showed a 4.3–4.8 times increase compared to the control group. The Arabidopsis wri1 loss-of-function mutant was almost completely rescued by the strong expression of castor bean WRI1 from a seed-specific promoter, resulting in a total FA content close to that of non-transgenic seeds [41][60]. LEC1 is a member of the nuclear factor-YB (NF-YB) family of TFs, and the individual overexpression of Arabidopsis LEC1 or L1L, or the overexpression of their homologous genes BnLEC1 or BnL1L from rapeseed, resulted in significantly increased FA levels in transgenic Arabidopsis [42][43][61,62]. FUS3, another member of the plant-specific B3 domain family, plays an important role in recognizing and binding to the RY element CATGCA, which is found in the promoters of many genes. fus3 mutants in rapeseed showed a lower total SOC than the wild type [44][64]. Inducible expression of Arabidopsis FUS3 increased the oil content of Arabidopsis seedlings to 6% of dry weight, which was more than 50-fold higher than that of non-transgenic seeds. Similarly, the inducible expression of FUS3 in Nicotiana tabacum L. cv Bright Yellow2 (BY2) cells increased TAG accumulation, and the co-expression of FUS3 and DGAT1 further increased the TAG levels to 4% of the dry weight. ABI3 is also a member of the B3 domain TF family. The heterologous expression of GmABI3 from soybean in Arabidopsis significantly increased TAG content and altered the FA composition of seeds. In addition, GmABI3 expression successfully complemented the phenotype and oil content of Arabidopsis abi3 mutant seeds [45][66]. In another study, tobacco leaf protoplasts were prepared for transfection with effector constructs overexpressing WRI1 and/or DGAT1 alone or together with one of the master regulators ABI3, FUS3, LEC1, or LEC2. The co-expression of FUS3 and ABI3 with WRI1 and DGAT1 significantly increased the content of total non-polar lipids of the protoplasts. Notably, the expression of ABI3 overexpression alone resulted in the highest accumulation of total non-polar lipid in protoplasts. Furthermore, in contrast to LEC2, the co-expression of ABI3 further increased the lipid content of protoplasts transiently expressing WRI1 and DGAT1 [46][67]. MYB TFs comprise one of the largest gene families in plants and are associated with the regulation of plant growth and development, metabolism, morphology, and cellular patterning [47][70]. MYB92 can directly bind to the promoter of and activate the transcription of Biotin carboxyl carrier protein 2 (BCCP2), which encodes a component of the FA biosynthetic pathway. The overexpression of Arabidopsis MYB92 in N. benthamiana induced the expression of FA biosynthesis genes, resulting in the accumulation of various types of lipids [48][71]. DECREASE WAX BIOSYNTHESIS 2 (DEWAX2) is a member of the AP2-EREBP TF family in Arabidopsis and negatively regulates epidermal wax deposition by directly binding to the promoters of LACS1, LACS2, KCSII, and ECERIFERUM1 (CER1) to inhibit their expression [4][9]. Growth-regulating factor 2a (BnRGF2a) was identified in rapeseed based on its differential expression between two rapeseed lines with differing seed oil production. Arabidopsis lines overexpressing BnGRF2 showed improved seed quality and oil content. In addition, transcriptome analysis revealed that some genes related to cell proliferation, photosynthesis, and oil biosynthesis were upregulated in these overexpression lines, suggesting that cell number and photosynthesis are linked to the observed increase in seed weight and oil content [49][73].2.3. Advances in Multi-Omics Studies of Oilseed Crops
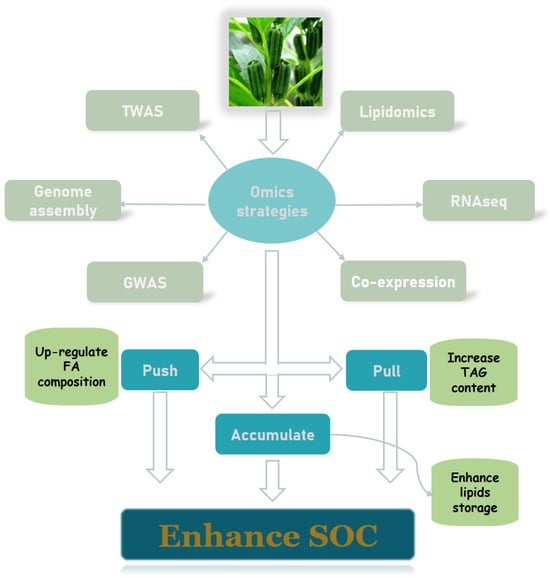
3.3. Advances in Multi-Omics Studies of Oilseed Crops
With the abundance of high-throughput sequencing information, the mining of large amounts of biological data is becoming increasingly popular and feasible. The availability of genomic, transcriptomic, and other multi-omics data provides a powerful avenue to unravel the detailed regulation of lipid metabolism [50][51][77,78]. From an omics perspective, the following aspects of lipid metabolism in oilseed crops are currently of interest (Figure 34).