The first GH3 gene was identified from
Glycine max as a rapid early auxin-responsive gene. Subsequent research has established the widespread distribution of the GH3 gene family across the plant kingdom, encompassing species from
Arabidopsis thaliana to
Oryza sativa,
Zea mays,
Triticum aestivum, and even non-vascular plants like
Physcomitrella patens and
Marchantia polymorpha [8][11][14][15][16][17][18][8,11,15,16,17,18,19]. Based on sequence homology and substrate specificity, the GH3 family in Arabidopsis is categorized into three distinct groups: Group I, II, and III. Group I GH3 genes are known to encode enzymes that synthesize amides from jasmonic acid (JA) or salicylic acid (SA). Group II GH3 enzymes function as IAA-amido synthetases, and Group III has been shown to catalyze the conjugation between amino acids and 4-substituted benzoates or indole-3-butyric acid (IBA)
[8][19][8,20].
2. The Catalytic Mechanisms and Substrate Specificity of GH3 Acyl Acid Amido Synthetase Enzyme
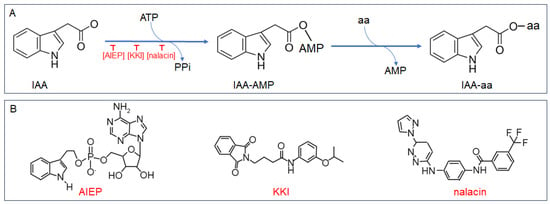
Chen et al. firstly utilized a combination of initial velocity and product inhibition analyses, alongside mass spectrometry, to delineate the kinetic and chemical mechanisms governing OsGH3.8 activity
[20][25]. They discovered that the conjugation of IAA with Asp operates via a ‘Bi Uni Uni Bi Ping Pong’ mechanism, as depicted in
Figure 1A. The process initiates with the binding of IAA and ATP, in the presence of Mg
2+, to the unoccupied enzyme. This interaction results in the formation of an adenylated IAA intermediate (IAA-AMP) and the concurrent release of pyrophosphate (PPi). Following this, Asp attaches to the enzyme•IAA•AMP complex, leading to the displacement of AMP and the establishment of an amide linkage between IAA and Asp. The final reaction products, IAA-Asp and AMP, are then released from the OsGH3.8 enzyme’s active site
[20][25].
Figure 1. Catalytic reaction and inhibitor structures of Group II GH3 amido synthetase. (A) Schematic representation of the total reaction mediated by Group II GH3 amido synthetases. (B) Chemical structures of inhibitors targeting Group II GH3 amido synthetases: AIEP (adenosine-59-[2-(1H-indol-3-yl) ethyl] phosphate), KKI (kakeimide), and nalacin (N-[4-[[6-(1H-pyrazol-1-yl)-3-pyridazinyl] amino] phenyl]-3-(trifluoromethyl)benzamide).
Structural analyses of the GH3 enzyme in Arabidopsis, grape, and rice have illuminated that both monocotyledons and dicotyledons employ a similar mechanism for AMP and IAA binding
[21][26]. The GH3 enzyme exhibits distinct acyl acid binding preferences, with specific residues within its active site conferring selectivity for particular substrates
[21][26]. In OsGH3.8, the amino acids Arg130 and Leu137 play a crucial role in substrate specificity. The mutation of Arg130 to Leu (Arg130-Leu) shifts the enzyme’s substrate preference from IAA to benzoate/SA, while an Arg130-Thr substitution favors JA over IAA.
3. The Modulation of IAA Homeostasis by Small Chemical Molecules via the Inhibition of GH3 Enzyme Activity
3.1. AIEP, the First Chemical Inhibitor of Auxin Conjugation
The GH3-dependent IAA conjugation initiates when the unbound GH3 enzyme interacts with ATP and IAA, leading to the formation of IAA•AMP. Böttcher et al. engineered and synthesized a stable analogue of IAA•AMP, named AIEP. This molecule competes with ATP and IAA for the binding sites on the GH3 enzyme at the onset of catalysis
[22][30]. The competitive inhibitory effect of AIEP on ATP and IAA binding was validated through substrate velocity experiments involving VvGH3.1 and VvGH3.6 from grape.
3.2. KKI, a Specific Inhibitor of IAA-Conjugating GH3 Enzymes
In the quest to identify inhibitors of IAA-conjugating GH3 enzymes, researchers leveraged Arabidopsis
AtGH3.6 overexpression plants as a biological assay system. They screened a synthetic chemical library comprising 10,000 compounds for agents capable of reverting the altered root hair growth phenotype of AtGH3.6-overexpressed lines. This led to the initial identification of compound ‘1’, followed by the synthesis of 25 derivatives of ‘1’. Among these, kakeimide (KKI) emerged as a highly potent inhibitor. (
Figure 1B). KKI functions by directly interacting with the IAA binding site within the GH3•ATP complex, forming a stable GH3•ATP•KKI ternary complex that impedes the synthesis of IAA-amino acid conjugates
[23][32]. Validation experiments demonstrated KKI’s effectiveness in targeting the IAA binding sites of various GH3 enzymes, notably VvGH3.1, AtGH3.5, and OsGH3.8, while sparing the IBA binding site of AtGH3.15. This specificity, coupled with KKI’s lack of interference in jasmonic acid (JA) homeostasis, underscores its role as a specific inhibitor of IAA-conjugating GH3 enzymes
[23][32].
3.3. Nalacin, a Potent Inhibitor Targeting Group II GH3 Enzymes
Nalacin was identified from a chemical screen by observing the auxin-related root phenotypes in the Arabidopsis wild-type Col-0 (
Figure 1B). Subsequent studies have shown that nalacin competitively inhibits substrate acceptance by AtGH3.6 and AtGH3.11 through trifluoromethyl phenyl occupancy of the IAA binding site of AtGH3s, suggesting that nalacin also functions in the first step of the ‘Bi Uni Uni Bi Ping Pong’ reaction of GH3 enzymes. Unlike KKI, which selectively inhibits only class II GH3 enzymes, nalacin also impedes the formation of JA amino acid conjugates mediated by AtGH3.11, albeit through a distinct binding mode
[24][31].
4. The Transcriptional Control of GH3 Enzymes in Plant Growth, Development, and Stress Adaptation
4.1. GH3-Dependent IAA Conjugation Is Involved in Regulating Multiple Developmental Processes
In Arabidopsis, eight Group II
GH3 genes are involved in catalyzing IAA conjugation to amino acids. Due to redundant gene functions, mutations in single genes result in only subtle phenotypic changes and modified sensitivity to exogenous IAA
[25][54]. In contrast, mutants with overexpressed
GH3 genes, obtained through activation tag insertion, provide a more discernible phenotype for study. For instance,
AtGH3.2 and
AtGH3.6 were identified through the screening of their overexpressed mutants,
ydk1-D and
dfl1-D, respectively
[25][26][54,55]. Interestingly, however, despite all overexpressing genes being closely related to
GH3 Group II family members, they still showed inconsistent phenotypes. Under various light conditions, the
dfl1-D mutant displayed shortened hypocotyls exclusively in light, while the
ydk1-D mutant showed this phenotype under both light and dark conditions. Additionally, the
ydk1-D mutant had a shorter primary root but did not exhibit significant difference in susceptibility to auxin-mediated root growth inhibition. In contrast, the
dfl1-D mutant was resistant to IAA-mediated root growth inhibition and did not present a short-root phenotype compared with the wild type
[25][26][54,55].
As early auxin response genes, Group II
GH3 genes play a significant role downstream of auxin response factors (ARFs), which are key elements in the auxin signaling pathway
[27][57]. Research has elucidated the involvement of ARF6, AtARF7, AtARF8, and AtARF17 in the transcriptional regulation of several
AtGH3 genes. AtARF7 and AtARF8 are known to positively regulate the transcription of
AtGH3.2,
AtGH3.5, and
AtGH3.6, influencing hypocotyl elongation under different light conditions
[25][28][54,58]. In contrast, AtARF17 has a negative regulatory role on
AtGH3.5 and
AtGH3.6 transcription, which is essential for proper plant development. The microRNA AtmiR160 directly targets
AtARF17 mRNA, which is crucial for normal leaf and root growth
[29][59]. In terms of adventitious root development, AtARF6 and AtARF8 act as positive regulators, while AtARF17 functions as a negative regulator. They co-regulate the transcription of
AtGH3.3,
AtGH3.5, and
AtGH3.6, impacting JA conjugation with amino acids, but not IAA conjugation
[30][60].
4.2. The Integration of Hormonal Signals in GH3-Dependent IAA Conjugation’s Responses to Abiotic Stresses
4.2.1. Drought Stress
In Arabidopsis,
AtGH3.5 has been observed to respond rapidly to drought conditions, with the
wes1-D mutant, which overexpresses
AtGH3.5, exhibiting enhanced drought resistance
[31][79]. A subsequent study revealed that AtMYB96 modulates the expression of several
AtGH3 genes, including
AtGH3.3,
AtGH3.5, and
AtGH3.6, under drought stress through an abscisic acid (ABA)-dependent pathway
[32][80]. This finding underscores the importance of ABA signaling in modulating
GH3 gene expression during drought response. Additionally, the
gh3oct mutant, with knockouts of all
Group II GH3 genes (
GH3.1,
2,
3,
4,
5,
6,
9,
17), exhibits increased drought tolerance, further highlighting the role of these genes in drought response mechanisms
[33][81]. In rice, the activation of
OsGH3.13 has been linked to a reduction in free IAA levels, leading to a structural adaptation in the leaves, such as thicker blades, which enhance drought tolerance by minimizing water loss
[34][74].
4.2.2
5. Atypical Roles of Group II GH3 and IAA-Amino Acids
. Temperature (Heat/Cold/Freezing) Stress
In Arabidopsis, the transcription of
AtGH3.5 is notably responsive to temperature extremes, showing increased levels under both low (4 °C) and high (37 °C) temperature conditions. The
wes1-D mutant, characterized by the overexpression of
AtGH3.5, shows increased survival after exposure to freezing temperatures (−7 °C). This suggests a broad regulatory role for
AtGH3.5 across a spectrum of temperature stresses
[35][82]. In rice, the overexpression of
OsGH3.2 leads to a reduction in free IAA levels, thereby activating cold-responsive genes and enhancing the plant’s ability to scavenge reactive oxygen species (ROS).
4.2.3. Salt and Osmotic Stress
All root-expressed Group II
GH3 genes in Arabidopsis are upregulated following treatment with NaCl at concentrations of 75 mM and 150 mM. The
Atgh3oct mutant, with combined knockouts of all Group II
GH3 genes, exhibits greater resilience to NaCl stress compared to the wild type
[33][81]. This enhanced tolerance also extends to sorbitol and mannitol exposure, suggesting that Group II GH3s may confer broad osmotic stress resistance, inclusive of salinity stress. Further investigation reveals that NaCl treatment increases
AtACS2 transcription, leading to the accumulation of the ethylene precursor ACC, which in turn downregulates
AtGH3.5 and
AtGH3.9 transcription, maintaining free IAA levels and primary root growth
[36][83].
4.2.4. Ammonium (NH4+) Stress
NH
4+ serves as a vital nitrogen source for plants, but when available in excess, it can be detrimental to growth
[37][38][85,86]. Prior research indicates that high NH
4+ levels lead to a reduction in free IAA
[39][40][41][87,88,89]. In Arabidopsis, elevated NH
4+ conditions trigger the induction of nearly all Group II
GH3 genes, which in turn accelerates the conjugation of free IAA to amino acids
[39][87].
4.2.5. Pathogen Stress
In response to pathogen attacks, plants activate a comprehensive defense strategy: (1) they initiate a hypersensitive response leading to rapid programmed cell death at the infection site alongside other defense responses; (2) they activate systemic-acquired resistance (SAR) in distal tissues; and (3) they activate basal immunity to limit pathogen growth
[42][91]. GH3-dependent IAA conjugation is intricately involved in these plant defense mechanisms. In Arabidopsis, the pathogens
B. cinerea and
P. syringae pv tomato (
Pst) DC3000 significantly upregulate
AtGH3.2 and
AtGH3.3 transcription. Loss-of-function mutations in
AtGH3.2 or
AtGH3.4 enhance resistance to both pathogens, suggesting that
AtGH3.2,
AtGH3.3, and
AtGH3.4 may negatively influence the plant’s response to
B. cinerea and
Pst DC3000
[43][92]. The overexpression of mutant
gh3.5-1D shows a compromised hypersensitive response but retains normal SAR and basal immunity, whereas the
Atgh3.5 mutant exhibits a defective SAR response yet maintains a typical hypersensitive response and basal immunity. In contrast, the
dfl1-D mutant displays altered hypersensitive and basal immune responses
[42][91].
5. Atypical Roles of Group II GH3 and IAA-Amino Acids
5.1. The Roles of Group II GH3 beyond the Catalyzation of IAA-Amino Acid Conjugate Formation
5.1. The Roles of Group II GH3 beyond the Catalyzation of IAA-Amino Acid Conjugate Formation
The capacity of Group II GH3 enzymes extends beyond the synthesis of IAA-amino acid conjugates. Research using recombinant GH3 IAA-amino acid synthetase from pea has revealed the enzyme’s ability to conjugate IAA not only to aspartate but also to proteins in immature seeds. The proposition that IAA conjugation to proteins may serve a regulatory function acting as a prosthetic group and influencing protein activity via posttranslational modifications is a compelling avenue for further exploration
[44][45][97,98].
5.2. The Specialized Functions of IAA-Amino Acid Conjugates beyond Their Role as Auxin Stock
Previous studies found that the exogenous addition of IAA-aa and IAA both rapidly increased content-free IAA levels and exhibited similar high growth factor phenotypes, suggesting that IAA-aa is a storage form of IAA
[46][103]. Subsequent studies found that these IAA-aa, IAA-Leu, and IAA-Ala could be reversibly converted to free IAA by the action of the hydrolases, IAA-LEUCINE RESIS TANT1 (ILR1), ILR1-LIKE proteins (ILLs), and IAA-ALANINE RESISTANT3 (IAR3)
[5][47][48][5,104,105]. IAA-Glu and IAA-Asp, once considered only as degradation intermediates, are now recognized as reversible storage forms
[49][14]. Beyond storage, IAA-aa have been identified as possessing unique biological functions. IAA-Trp, for instance, acts as a ‘super inactivator’ by not only consuming free IAA for its synthesis but also antagonizing the activity of residual IAA, with IAA-Trp significantly mitigating root inhibition effects caused by IAA
[50][24]. IAA-Asp has been reported to have more diverse roles: (1) correcting the temperature sensitivity of henbane (
Hyoscyamus muticus) XIlB2 (temperature-sensitive variant) cells
[51][22]; (2) IAA-Asp directly and specifically enhance the pea (
Pisum sativum) responses to abiotic stress by increasing the antioxidant enzyme activity and then reducing the H
2O
2 concentration
[52][23]; (3) IAA-Asp as a ripening signal in grapes (
Vitis vinifera) can be perceived at a certain stage of fruit development; however, the mechanism of sensing remains unknown
[53][21]; and (4) IAA-Asp promotes pathogen development in plants by regulating the transcription of virulence genes
[43][92]. These insights suggest that IAA-amino acids are not just byproducts of GH3 activity but are biologically active molecules with specific roles.
 Structural analyses of the GH3 enzyme in Arabidopsis, grape, and rice have illuminated that both monocotyledons and dicotyledons employ a similar mechanism for AMP and IAA binding [21][26]. The GH3 enzyme exhibits distinct acyl acid binding preferences, with specific residues within its active site conferring selectivity for particular substrates [21][26]. In OsGH3.8, the amino acids Arg130 and Leu137 play a crucial role in substrate specificity. The mutation of Arg130 to Leu (Arg130-Leu) shifts the enzyme’s substrate preference from IAA to benzoate/SA, while an Arg130-Thr substitution favors JA over IAA.
Structural analyses of the GH3 enzyme in Arabidopsis, grape, and rice have illuminated that both monocotyledons and dicotyledons employ a similar mechanism for AMP and IAA binding [21][26]. The GH3 enzyme exhibits distinct acyl acid binding preferences, with specific residues within its active site conferring selectivity for particular substrates [21][26]. In OsGH3.8, the amino acids Arg130 and Leu137 play a crucial role in substrate specificity. The mutation of Arg130 to Leu (Arg130-Leu) shifts the enzyme’s substrate preference from IAA to benzoate/SA, while an Arg130-Thr substitution favors JA over IAA.