Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Jose M. Mulet and Version 2 by Rita Xu.
Plants, the cornerstone of life on Earth, are constantly struggling with a number of challenges arising from both biotic and abiotic stressors. To overcome these adverse factors, plants have evolved complex defense mechanisms involving both a number of cell signaling pathways and a complex network of interactions with microorganisms. Among these interactions, the relationship between symbiotic arbuscular mycorrhizal fungi (AMF) and strigolactones (SLs) stands as an important interplay that has a significant impact on increased resistance to environmental stresses and improved nutrient uptake and the subsequent enhanced plant growth.
- arbuscular mycorrhizal fungi
- strigolactone
- synergistic interaction
1. Introduction
Plants are pivotal for many ecosystems and thus essential for the survival of virtually all living organisms. They are not only a source of food for humans and animals and the main point of entrance of solar energy and organic carbon in ecosystems, but they also play a critical role in regulating the Earth’s climate and sustaining the planet’s biodiversity. However, plants are constantly under threat from various biotic and abiotic stresses, such as pests, diseases and environmental factors, like drought and salinity [1][2][3][1,2,3]. Under the current context of anthropogenic global warming, forest and cultivated plants must adapt to the novel conditions or become extinct [4][5][4,5]. To overcome these challenges, plants have evolved complex mechanisms that involve a wide range of signaling pathways and interactions with other organisms, including microbes.
The significance of arbuscular mycorrhizal fungi (AMF) and strigolactones (SLs) in plant–microbe interactions lies in their ability to positively influence plant growth, development and overall health [6][7][6,7]. AMF establish mutualistic associations with plants, colonizing their root systems and providing various benefits [8]. These benefits include enhanced nutrient acquisition, such as an increased availability of phosphorus and micronutrients, improved water uptake and protection against biotic and abiotic stresses [2]. AMF can also induce systemic resistance in plants, making them more resistant to pathogens and pests and abiotic stresses [9][10][9,10]. On the other hand, SLs, a class of plant hormones, regulate several critical processes in plants [11]. They are involved in shaping root architecture, promoting the development of lateral roots and stimulating the establishment of beneficial associations with symbiotic microbes, such as AMF [7][11][12][7,11,12]. SLs also play a role in plant defense responses, including the activation of systemic defense mechanisms against pathogens and the induction of plant secondary metabolites [13].
Recent studies have shown that the synergistic interaction between AMF and SLs can have a significant impact on plant–microbe interactions and plant development. The combined effects of these two components result in improved plant growth, an increased resistance to biotic and abiotic stresses and enhanced nutrient uptake [6][7][14][6,7,14]. Therefore, understanding the interplay between these two components is crucial for developing sustainable agricultural practices and improving crop yields.
Mechanistically, AMF have been found to influence the production and release of SLs by plants [15]. They can stimulate the synthesis and secretion of SLs, which act as signaling molecules to attract beneficial microbes and promote symbiotic interactions [16]. In turn, SLs can modulate the colonization and establishment of AMF within the plant root system, facilitating their beneficial effects [6][7][14][6,7,14]. The significance of this synergistic interaction becomes evident in its potential to improve crop productivity, nutrient utilization and plant resistance in the face of environmental challenges [6][14][6,14]. By harnessing the combined effects of AMF and SLs, agricultural practices can be optimized to enhance nutrient acquisition efficiency, leading to increased crop yields.
2. Overview of Arbuscular Mycorrhizal Fungi
Arbuscular mycorrhizal fungi extend their hyphae into the soil, exploring a larger volume and accessing nutrients inaccessible to the root [17][18][17,18]. In addition, AMF spore dynamics are found at higher densities in rhizosphere soil [19]. They can solubilize nutrients from solid soil particles and organic matter, making them available for plant uptake [20]. Furthermore, AMF release enzymes that break down complex organic compounds, releasing nutrients for plant uptake [21]. They can convert inorganic forms of macronutrients, such as phosphorus, into molecules that plants can assimilate [22]. This promotes efficient nutrient utilization by plants. Furthermore, AMF can affect the synthesis, release and signaling pathways of growth-promoting phytohormones, such as SLs, auxins and cytokinins in plants, leading to enhanced plant growth and development [15][23][15,23]. AMF can induce systemic resistance in plants, preparing them for pathogen and pest attacks. For example, AMF activate plant defense mechanisms by triggering the expression of defense-related genes [24]. This leads to the production of defense compounds, such as pathogenesis-related proteins and antimicrobial peptides that protect plants against pathogens and pests [25]. Furthermore, AMF prime the plant’s immune system, enabling a more rapid and effective defense response upon pathogen or pest attacks [25]. This priming improves the plant’s ability to recognize and respond to subsequent challenges, increasing its overall disease resistance.2.1. Molecular Signaling
AMF are beneficial soil microorganisms that form symbiotic relationships with plants, enhancing nutrient uptake and contributing to the health and sustainability of terrestrial ecosystems [2]. In the early stages of AMF symbiosis, molecular signals are exchanged between the plant and the fungus [26][27][26,27]. The plant releases signaling molecules, such as SLs, into the soil in response to nutrient stress [28]. AMF hyphae in the soil sense these signals and initiate a molecular response [7][29][7,29]. This includes the expression of genes related to hyphal growth and colonization. The growth and branching of AMF hyphae towards plant roots are regulated by various signaling pathways [17][18][17,18]. These pathways involve receptor proteins on the fungal hyphae that recognize specific plant signals. The interaction between plant root cells and AMF hyphae involves molecular cross-talk, allowing the hyphae to penetrate root cells [30]. This process involves the exchange of signaling molecules and the activation of genes that facilitate the establishment of arbuscules, specialized structures within root cells where nutrient exchange occurs [31]. Nutrient exchange in AMF symbiosis is highly dependent on molecular processes. Within arbuscules, specific transporters and channels facilitate the movement of nutrients between the plant and the fungus [7][28][7,28]. The plant provides sugars obtained by photosynthesis and organic compounds from the plant’s own metabolic pathways through molecular transporters, while the fungus supplies the plant with essential nutrients, like phosphorus and nitrogen, which the fungi have solubilized from the soil [32][33][32,33]. AMF also influence plant defense and stress responses at the molecular level [9]. They can enhance the plant’s ability to withstand various environmental stresses, such as biotic and abiotic [2][10][2,10]. The molecular mechanisms involved in these responses include the activation of defense-related genes and the modulation of plant hormone signaling pathways [9][34][9,34]. The molecular interactions between plants and AMF are highly intricate and involve the exchange of signaling molecules, gene regulation and the coordination of various molecular processes to establish and maintain this symbiotic relationship [34][35][34,35]. The synergy between the two organisms at the molecular level results in improved nutrient uptake and enhanced plant resilience.2.2. Mycorrhizal Symbiosis Genes
Mycorrhizal symbiosis involves the interaction between plants and mycorrhizal fungi, and several genes in both partners are crucial for the establishment and maintenance of this symbiotic relationship (Figure 1). Some of the key genes and molecular components involved in mycorrhizal symbiosis are established below.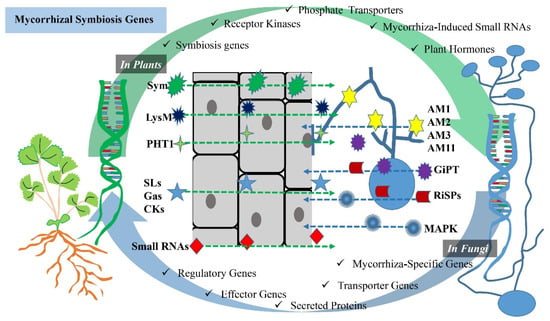
Figure 1. Schematic visualization of genes involved in AM symbiosis. The interaction between plants and AMF involves several genes and molecular components that are crucial for the establishment and maintenance of the symbiotic relationship. While Sym genes, receptor kinases, phosphate (Pi) transporters, mycorrhiza-derived small RNAs and plant hormones in plants are plant-derived genes and components, mycorrhiza-specific genes, transporter genes, secreted proteins, effector genes and regulatory genes are genes of a mycorrhizal fungi origin. These genes and molecular components in both plants and mycorrhizal fungi are essential for the successful establishment and maintenance of mycorrhizal symbiosis, which benefits both partners by improving nutrient exchange and enhancing plant growth.
2.2.1. In Plants
Sym genes (symbiosis genes): these are plant genes that are specifically involved in the establishment and regulation of mycorrhizal symbiosis [36]. They encode various proteins and transcription factors necessary for the recognition of fungal partners, the development of symbiotic structures and the regulation of nutrient exchange [37]. Receptor kinases: plant receptor kinases, such as the LysM receptor kinases, play a role in recognizing fungal signals and initiating the signaling cascade leading to mycorrhizal symbiosis [38][39][38,39]. These receptors are essential for the early recognition of mycorrhizal fungi [38]. Phosphate (Pi) transporters: plants upregulate genes encoding phosphate transporters in response to mycorrhizal colonization [40]. These transporters facilitate the uptake of phosphorus from the fungal partner [41]. Mycorrhiza-induced small RNAs: plants produce small RNAs in response to mycorrhizal colonization, which may play a role in regulating the symbiotic interaction [42]. Plant hormones: in general, plant hormones include activities directed towards the development and branching of plants. For example, strigolactones play an important role in stimulating branching activity in plants, contributing to the development of an extensive root system [43]. The gibberellin hormone promotes root elongation and influences plant growth and development [44]. Cytokinins are also involved in the regulation of plant branching. Cytokinins influence the overall architecture of the plant by promoting the growth of lateral buds [45]. Furthermore, various plant hormones, such as strigolactones, gibberellin and cytokinins, are involved in signaling and regulating mycorrhizal symbiosis [7][46][47][7,46,47]. Strigolactones, for example, are known to promote hyphal growth and root colonization by mycorrhizal fungi [6].2.2.2. In Mycorrhizal Fungi
Mycorrhiza-specific genes: fungi possess genes that are specifically expressed during mycorrhizal symbiosis [48]. These genes are involved in hyphal growth, the establishment of symbiotic structures, like arbuscules, and nutrient exchange [48][49][48,49]. For example, Colard et al. [48] reported that AMF-specific AM1, AM2, AM3 and AM11 genes were activated at the pre-symbiotic stage. Transporter genes: mycorrhizal fungi have transporter genes that code for proteins responsible for the uptake and transfer of nutrients to the plant host [50] (Table 1). For example, phosphate transporters are crucial for delivering phosphorus to the plant [51]. Maldonado-Mendoza et al. [52] revealed that this was thanks to the GiPT gene for Glomus intraradices. Secreted proteins: fungi produce secreted proteins, some of which may be involved in facilitating the interaction with plant roots or modulating plant immune responses [53][54][53,54]. For example, Kamel et al.’s [55] study on Rhizophagus irregularis revealed that this species had a large number of putative secreted proteins (RiSPs), which could be of great importance in establishing symbiosis. Effector genes: some mycorrhizal fungi may produce effector proteins that can manipulate plant host defenses or signaling pathways to promote symbiosis [56][57][56,57]. Regulatory genes: fungi have genes involved in the regulation of their responses to the plant host and environmental cues [58][59][58,59]. These genes help the fungi adapt to different plant partners and environmental conditions [60]. For example, Huang et al. [59] reported that using mitogen-activated protein kinase (MAPK) signaling for the interactions between AMF and apple plant hosts was shown to increase apple drought tolerance. miRNA: there have also been some miRNAs identified as participants in this regulation, such as miR167, miR394 and miR156 [60]. Understanding the genetic and molecular basis of mycorrhizal symbiosis is an active area of research. These genes and molecular components play a critical role in the formation and maintenance of this beneficial mutualistic relationship, contributing to plant nutrient acquisition, stress tolerance and overall ecosystem health (Table 1).Table 1. AMF-derived genes involved in mycorrhizal symbiosis.
| AMF-Induced Genes | References | ||
|---|---|---|---|
| Mycorrhiza-Specific Genes | AM1, AM2, AM3, AM11 | Genes active in the AMF-induced pre-symbiotic stage | [48] |
| AM10, AM14, AM15, AM20, AM24, AM25, AM26, AM29 | Genes active in the AMF-induced early and mature symbiotic stages | [49] | |
| Transporter Genes | GiPT | AMF-induced plant P transporter genes | [52] |
| StPT3 | [61] | ||
| OsPT11 | [62] | ||
| MtPT4 | [63] | ||
| PT11 | [49] | ||
| MtZIP5 | AMF-induced plant Zn transporter gene | [64] | |
| Secreted Proteins | LbMiSSP7 | Secreted proteins regulated by AMF | [65] |
| LjCLE19, LjCLE20 | [66] | ||
| RiSP | [55] | ||
| Effector Genes | RiSLM | AMF-induced effector genes | [67] |
| RirG175680, RirG165580, RirG263220, RirG200050, jgi.p|Gloin1|346360, RirG013260, RirG267270, jgi.p|Gloin1|154898, RirG043250, RirG045350, RirG101100, RirG043650, RirG257590, RirG187640, RirG180400, jgi.p|Gloin1|161262 | [68] | ||
| PvRxLR18, PvAVH52, PvRxLR28, PvRxLR67 | Effector genes against AMF-induced pathogen | [69] | |
| Regulatory Genes | 14-3-3 | Gene regulating AMF-induced ABA-related signaling pathway | [58] |
| MAPK | Genes regulated by AMF to enhance drought tolerance | [59] | |
| miR167, miR394, miR156 | [60] | ||
3. Strigolactones
Strigolactones (SLs), a class of plant hormones, have emerged as key regulators of plant growth, development and interactions with the environment. Striga lutea’s strigol, the first natural SL, was found as a germination stimulant; as a result, these compounds have subsequently been referred to as SLs [70]. They play a crucial role in various plant processes, such as root development, branching and responses to environmental stresses. These hormones are also known to interact with beneficial soil microorganisms, such as mycorrhizal fungi, to promote nutrient uptake and improve plant health. However, SLs secreted by plants cause the seeds of parasitic plants to germinate. This can cause problems in agricultural areas. In particular, the damage caused to agriculture by witchweed in Africa due to the parasitism on SL signaling is a major issue for farmers in developing countries [71].
3.1. Biosynthetic Pathway
According to Matusova et al. [72], SLs are derived from carotenoids, as evidenced by minimal SL accumulation after treatment with the carotenoid biosynthesis inhibitor fluridone in maize plants. The functional role of SLs can be related to their formation and production in response to the needs of the system during evolution [73]. Since it has been discovered that the gene involved in SL production has been reported for many plant species, including algae and bryophytes, it can be hypothesized that these SLs are important molecules that have long persisted in the evolutionary chain [73]. SLs are four-ring (A–D) compounds that change function by the attachment of various groups to the A and B rings [12][15][73][74][12,15,73,74]. SLs were initially thought to be sesquiterpene lactones, but were later shown to be apocarotenoid derivatives of carotenoid cleavage mediated by carotenoid cleavage dioxygenase (CCDs) enzymes [75]. A member of the CCD family is involved in the production of various apocarotenoid compounds, such as cyclohexenone and mycorradicin [76]. Initial biosynthesis occurs in plastids with the help of three plastid-specific enzymes: D (DWARF)27, CCD7 and CCD8. Carotenoid isomerase D27, carotenoid cleavage dioxygenases CCD7 and CCD8 and cytochrome P450 monooxygenases were identified as SL biosynthesis enzymes through genetic screening for shoot-branching mutants [15]. Furthermore, from mutants with excessive shoot branching, SL biosynthesis genes were found and called more axillary growth (MAX) in Arabidopsis thaliana [75], Ramosus (RMS) in Pisum sativum [77], decreased apical dominance (DAD) in petunia [78] and dwarf (D) in Oryza sativa [79][80][79,80].
D27 isomerase converts all-trans-β-carotene to 9-cys-β-carotene, and subsequent processes catalyzed by CCD7 and CCD8 convert 9-cys-β-carotene to carlactone (CL) with A- and D-ring structures [15][73][15,73]. CL is subsequently oxidized to different SL species by the cytochrome P450 monooxygenase MAX1 or other recently discovered enzymes. Briefly, reswearchers show the pathway of SL biosynthesis in Figure 2. Furthermore, not only enzymes downstream of CL but also enzymes upstream of CL may be structurally important for the formation of various SLs in SL biosynthesis. CCD7 and CCD8 carotenoid isomerases convert all-trans-carotene to CL as well as 3-hydroxy-carlactone (3-OH-CL) via zeaxanthin [81]. Although hydroxy-carlactone derivatives are the most common SLs in Arabidopsis [82], their significance for plant growth and development control is unknown.
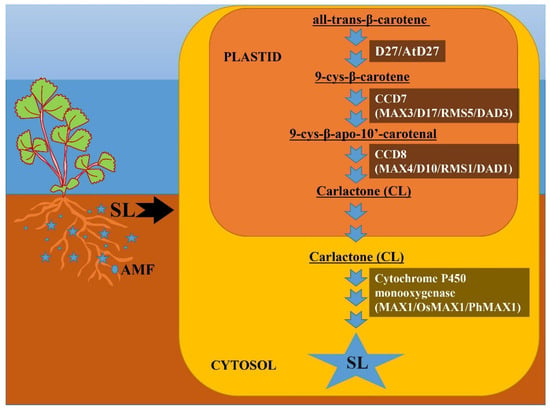
Figure 2. Biosynthetic pathway of strigolactone (SL). This figure depicts the SL biosynthetic route and important enzymes involved in biosynthesis. SL biosynthesis occurs in two distinct compartments: plastid and cytosol. All-trans-β-carotene is converted to carlactone (CL) in plastids via three intermediary stages catalyzed by D27 (At27), CCD7 (MAX3, D17, RMS5, DAD3) and CCD8 (MAX4, D10, RMS1, DAD1), respectively. Carlactone then enters the cytosol, where it is metabolized to several other SLs via cytochrome P450 monooxygenase (MAX1, OsMAX1, PhMAX1) and numerous other unidentified enzymes.
Recent studies have focused on modifying genes in the Strigolactone biosynthesis pathway using CRISPR/Cas9 gene-editing techniques [83][84][85][83,84,85]. Such genetic modifications can affect the biological processes of plants, such as root development, water use and nutrient uptake, and hence increase their interaction with AMF and abiotic stress tolerance. For example, the carotenoid cleavage dioxygenase 8 (CCD8) genes (SbCCD8a and SbCCD8b), which have been shown to be involved in strigolactone biosynthesis in sorghum, were manipulated by two CRISPR/Cas9-mediated genes and were found to enhance weed control and the activity of beneficial microorganisms [85].
3.2. Physiological Functions
Symbiotic interactions: SLs are involved in the establishment of symbiotic associations, particularly with arbuscular mycorrhizal fungi [6][12][14][6,12,14]. They act as rhizosphere signaling molecules, attracting AMF hyphae towards the plant roots and stimulating the branching of fungal hyphae in the soil. Akiyama et al. [86] demonstrated that sesquiterpenes, 5-deoxy-strigol, sorgolactone and strigol extracted from Lotus japonicus exudates promoted extended hyphal branching in AMF. A comparable finding was also seen in the synthetic counterpart of sorgolactone [87]. This association enhanced nutrient uptake, particularly phosphorus, by increasing the surface area available for nutrient absorption [6][7][14][6,7,14]. SLs also regulate the development of symbiotic structures, such as arbuscules, which facilitate nutrient exchange between the fungus and the plant [7].
Plant defense responses: SLs contribute to plant defense responses against pathogens and pests [88]. They can prime plants for enhanced resistance by activating defense-related genes and signaling pathways [89]. For example, Nasir et al. [90] found that SLs positively regulated defense against Magnaporthe oryzae in rice, and Xu et al. [91] positively regulated defense against root-knot nematodes in tomatoes. SLs can also influence the synthesis of secondary metabolites involved in plant defense mechanisms [92].
Shoot branching: one of the well-known roles of SLs is their influence on shoot branching. SLs act as inhibitors of bud outgrowth, promoting apical dominance and limiting the growth of lateral buds [93]. By suppressing the branching of shoots, SLs ensure the allocation of resources to the main shoot and control the overall architecture of the plant [43]. Manipulating SL levels or signaling can lead to alterations in shoot-branching patterns and can be utilized to enhance branching or promote lateral bud growth when desired [94]. For example, the involvement of SLs in regulating bud growth in Pisum sativum and Arabidopsis thaliana was demonstrated by Brewer et al. [95].
Overall plant growth and development: SLs have broader effects on plant growth and development. They contribute to various processes, such as seed germination, stomatal closure and responses to environmental stresses [96][97][98][96,97,98]. For example, Ha et al. [99] showed that the genetic modulation of SL content/response could provide a new approach for the development of plants with better environmental stress tolerance. SLs can affect seed dormancy and germination by inhibiting or promoting the germination process, depending on the species and environmental conditions [100]. They also regulate stomatal aperture, reducing water loss through transpiration and aiding in water-use efficiency [97]. SLs influence plant responses to abiotic stresses, such as drought and salinity, by modulating physiological and molecular responses that enhance stress tolerance [99].