1. Introduction
Arecaceae (syn.
Palmae), colloquially known as palm trees, is one of the best known and most extensively cultivated plant families, comprising around 2600 species in 181 genera [
1]. Palms are important plants in terms of human exploitation in their native range. Some species, such as oil (
Elaeis species), coconut (
Cocos nucifera), rattan (
Calamus species), and date (
Phoenix dactylifera) palms, are extremely important in the international trade [
2,
3,
4]. Moreover, palms are highly prized as ornamentals due to their great decorative potential. Thus, although almost exclusively native to tropical or sub-tropical countries, these perennial monocotyledonous trees are currently distributed worldwide due to their use as ornamental plants [
4,
5]. Therefore, palm trees have become a distinctive component of the urban landscape and an important accessory in interior decoration and in floristry.
Over the last 30 years, a considerable number of studies have shown the association of a diverse range of fungi with palm tissues. The extensive research carried out in Southeast Asia and Australasia showed that palms are a rich source of previously unknown fungal taxa. Thus, many species and genera were formally identified and described as new to science based on palm collections, e.g., refs. [
6,
7,
8]. The comprehensive isolation of fungi from palm tissues has proven that these fungi are an important and taxonomically diverse assemblage that is often referred to as palm fungi or palmicolous fungi, e.g., ref. [
9]. While most studies on palm fungi have focused on systematic and descriptive taxonomy, a few studies have explored the biodiversity and ecology of these microfungi, e.g., refs. [
10,
11,
12,
13,
14,
15,
16]. Taxonomically, palm fungi are one of the most diverse groups of fungi. The currently established figure is that more than 1500 species of fungi have been described from palm hosts, with representatives from almost all major fungal classes [
17]. In earlier studies, most taxa were introduced, described, and arranged in different taxonomic ranks within the
Ascomycota based on morphological analyses. This approach was, however, subjective and many taxa were wrongly assigned or assigned to the
Ascomycota genera
incertae sedis.
2. Historical Account of Research on Palm Fungi and Reflections on Their Importance
The history of the study of palm fungi can be divided into three distinct periods. The first, between 1880 and 1920, includes the publications of classical mycologists, such as Hennings, Rehm, Penzig, Spegazzini, H. Sydow and P. Sydow, and Saccardo [
18,
19,
20,
21,
22,
23,
24,
25,
26,
27]. Although many fungal species were described from palm trees during this period, these publications are barely accessible or legible since most of them were written in Latin. The second, between 1920 and 1990, includes several mycologists who, for 70 years, occasionally reported the occurrence of fungi on palm tissues. However, almost no study on palm fungi in this period was very extensive. The third, which began in the 1990s and continues today, represents the first time that a group of mycologists has dedicated itself to specifically analysing the microfungi that occur on palm trees. This includes the research carried out by Hyde, his students, and colleagues, which represents not only a significant advance in the number of fungal species described on palm substrata, but also a considerable advance in the biodiversity of palm fungi.
2.1. History of Systematic and Descriptive Taxonomy Studies on Palm Fungi
There are numerous reports scattered throughout the literature on fungi collected from different parts of different species of palm trees and from different regions of the world. An overwhelming number of these studies have been dedicated to collecting and describing fungi that inhabit palm tissues, including new genera and new species. Although there are several reports from before the 1990s, the present overview focuses on the extensive studies carried out by Hyde and co-workers. Hyde’s research is the first body of theory to understand palm fungi as a fungal community with taxonomic characteristics, biological processes, and general diversity that is worth exploring in depth. Most of the descriptions prior to Hyde’s research, especially those from the early 20th century, consist of short Latin paragraphs that lack illustrations and/or give limited information about the identity of the fungi.
2.1.1. From Scattered to the First Systematic Reports on Palm Fungi
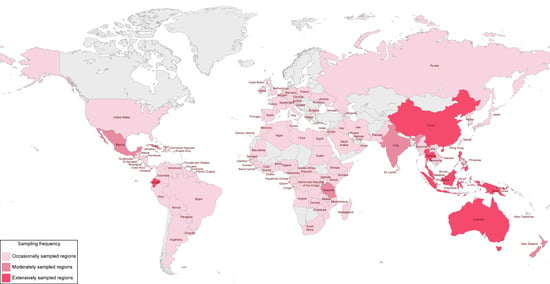
Before the extensive studies carried out by Hyde and co-workers, there were several occasional reports of fungi collected from different palms and tissues, such as leaf litter, green foliage, rachides, trunks, and stems. Although these collections come from different countries around the world, most of them come from tropical and subtropical regions where palm trees are almost exclusively native. A summary of some examples of these studies is given here considering the last decades of the 20th century. The sampling regions around the world where palm fungi have been recorded are mapped in Figure 1, according to the available literature.
Figure 1. World sampling regions of palm trees for associated fungi based on the available literature. Studies prior to 1950 were not considered when constructing the map, as most of the information is difficult to access. The regions/countries where palm trees were collected are named and delimited with coloured blocks and referred to in the chart legend. Extensively sampled regions are those where most studies have been carried out to specifically analyse palm fungi. Moderately and occasionally sampled regions are those where the reports of palm fungi were a consequence of occasional taxonomic or broader studies. Many occasionally sampled regions have not been mapped, as they are island countries that are barely visible due to the scale of the map. These include several Antilles islands of the Caribbean (Barbados, Dominica, Grenada, Guadeloupe, French West Indies, and Virgin Islands), Bermuda, African island countries (Comoro Islands, Mauritius, São Tomé and Príncipe, Réunion, and Seychelles), the Andaman and Nicobar Islands, and several islands in the Pacific Ocean (Cook Islands, French Polynesia, Niue, Samoa, and Tonga in Polynesia, Fiji, Vanuatu, and Solomon Islands in Melanesia, and Guam and Kiribati in Micronesia, Oceania). Figure source: created with a template available in the webserver for MapChart (
https://www.mapchart.net/, accessed on 15 September 2023).
Collections of palm species native to tropical regions of Central and South America and West and Southeast Africa (
Figure 1) have often yielded taxonomic novelties, including from new genera and species to new records on new hosts and new geographical distributions. For instance, Ellis [
28] reported new
Lacellina species on dead leaves of
Borassus aethiopum from Ghana and Sierra Leone (West Africa). Later, Ellis also reported new species of palm fungi from Central America, West Africa, and Malaysia on his series of papers and books concerning “Dematiaceous Hyphomycetes” [
29,
30,
31,
32].
Southeast Asia, Australasia, and India were also frequently surveyed (
Figure 1) and the first two would later become major regions for palm fungi investigation. For instance, while studying leaf-spotting hyphomycetes, Deighton [
33,
34] reported the new species
Cercospora raphiae and
Pseudocercospora carpentariae on leaves of
Raphia farinifera from Zimbabwe (Southeast Africa) and
Carpentaria acuminata from Australia. In the same year, several
Xylariaceae palmicolous fungi were recorded on the rainforests of North Sulawesi (Indonesia) by Rogers et al. [
35].
Several other regions were also only occasionally surveyed (
Figure 1), including, for instance, Venezuela (South America), where some palmicolous fungi, including new species, were described in the series of papers “Fungi venezuelani” [
36,
37,
38]; Argentina (South America), where Wright [
39] recorded the new genus and species
Agaricostilbum palmicola (currently synonymised under
Sterigmatomyces pulcherrimus) on weathered spathes of
Butia yatay and
Phoenix sp.; and Japan (East Asia), where Hino and Katumoto [
40,
41,
42] described some palmicolous fungi, including new species, in the series of papers entitled “Notes on fungi from western Japan”. Less frequently, the USA, European countries, and China, where only a few palm species are native, have also been the stage for collecting palm fungi (
Figure 1). For instance, several helicosporous fungi collected from palm tissues were reported from Hawaii (USA) by Goos [
43]; the new phialosporus hyphomycetes genus and species
Craspedodidymum elatum were reported on rotten petioles of
P. canariensis from Czech Republic (Central Europe) by Holubová-Jechová [
44]; and the new species
Zasmidium caryotae (as
Stenella caryotae) have been reported on the leaves of
Caryota mitis from Guangdong (China) by Liu and Liao [
45].
These first scattered reports of palmicolous fungi have established their importance as a highly diverse and understudied community that can be accessed on palms all over the world (
Figure 1). Some reports have expanded the geographical distribution of these fungi, which was important for the establishment of the geographical and ecological range of some important phytopathogens. For example, Samuels and Rossman [
46] during their studies on the
Amphisphaeriaceae-recorded
Leiosphaerella cocoes on fronds and fruits of
Cocos nucifera from several different regions, namely Dominican Republic, Guam, Indonesia, Mexico, USA, Tonga, Cook Islands, and Papua New Guinea, and reported two new species of
Oxydothis,
O. rhopalostylidis and
O. selenosporellae, on the leaf midrib of
Rhopalostylis sapida from New Zealand. It is worth mentioning that New Zealand was one of the first countries where several reports of palmicolous fungi were published more systematically, most of them from collections of the endemic palm tree
R. sapida (nikau palm), which is the only palm native to mainland New Zealand. Thus, several palm fungi, including many new species, were published in the series of papers “New Zealand Fungi” by Hughes [
47,
48,
49,
50,
51,
52,
53], as well as in many other occasional publications that were mostly later summarised by McKenzie et al. [
54] in their checklist of fungi recorded on nikau palm from New Zealand.
Two of the first most extensive studies on palm fungi include those of Pirozynski and Matsushima. Pirozynski [
55] reported forty-six species, including the new genus
Bondiella, six new species, and some new combinations of fungi from the oil palm
E. guineensis collected from Tanzania. Matsushima [
56,
57,
58,
59,
60,
61,
62,
63,
64,
65,
66,
67] and Matsushima and Matsushima [
68,
69] reported more than 300 fungi from palm litter, including 8 new genera, viz.
Apogaeumannomyces [
67],
Atrosetaphiale [
65],
Hyalobelemnospora,
Paradactylella [
64],
Setophiale,
Venustocephala [
65],
Veramyces [
64],
Verticimonosporium [
56], and more than 80 new species, mostly of rare and interesting hyphomycetous fungi. Most of these fungi were recorded in collections of palms from different regions of the world, from the Americas to Australia and Eastern Asia, including Peru, Guam, Taiwan, Cuba, Northern Queensland, and Ecuador (
Figure 1), were compiled in the classic book series “Matsushima mycological memoirs”.
Castañeda-Ruiz, Holubová-Jechová, Mena-Portales, and Mercado-Sierra were one of the first groups of mycologists to report several species of palm fungi growing on dead and decaying palm tissues, such as trunks, rachides, and petioles. Although these reports were abundant and consistent, their main purpose was not to report on the palmicolous fungi themselves, i.e., to understand the taxonomic or ecological structure of palm fungal communities. They were part of an investigation of hyphomycetes of Cuba, where many new species were introduced, e.g., refs. [
70,
71,
72,
73,
74,
75,
76,
77,
78,
79,
80,
81,
82,
83,
84,
85]. Most of these reports were summarised and discussed in series of papers, such as “Hifomicetes demaciáceos de Cuba” [
86,
87], “Studies on hyphomycetes from Cuba” [
88,
89,
90,
91,
92,
93,
94,
95], and “Nuevos o raros hifomicetes de Cuba” [
96,
97,
98,
99,
100,
101,
102], and books, such as “Hifomicetes demaciáceos de Sierra del Rosario, Cuba” [
103]. Although several of these reports were from undetermined palm trees and other particular known species, most of them were from the Cuban royal palm
Roystonea regia, where several new species were described, insomuch that Mercado-Sierra [
73] has described
R. regia as an “ideal substratum for the development of dematiaceous hyphomycetes”.
2.1.2. Hyde and Co-Workers and the Extensive Studies on Palm Fungi from Tropical Regions
The last 30 years has seen an extensive profusion of studies regarding palmicolous fungi by Hyde and co-workers. The extent and depth of their investigation yielded an impressive body of literature that made it possible to begin to understand and characterise the taxonomy of palm fungi, particularly the communities inhabiting tropical and subtropical palms growing on their native regions. These systematic studies culminated in the publication of three books entitled “Genera of ascomycetes from palms” [
104], “Palm microfungi” [
6], and “Microfungi of tropical and temperate palms” [
8], and a series of publications entitled “Fungi from palms”, comprising 49 papers where numerous new fungi to science were described [
105,
106,
107,
108,
109,
110,
111,
112,
113,
114,
115,
116,
117,
118,
119,
120,
121,
122,
123,
124,
125,
126,
127,
128,
129,
130,
131,
132,
133,
134,
135,
136,
137,
138,
139,
140,
141,
142,
143,
144,
145,
146,
147,
148,
149,
150,
151,
152,
153]. Genera that have been described as new to science and found on palm trees over the last three decades are summarised in
Table 1. Genera and respective families in subclasses of
Dothideomycetes and
Sordariomycetes with common representatives found on palm trees are summarised in
Table 2.
Hyde et al. have been studying microfungi from palms since 1988 when new species of the genera
Linocarpon,
Oxydothis, and
Astrosphaeriella (as
Trematosphaeria) were described from the fronds of the mangrove palm
Nypa fruticans from Brunei [
154,
155,
156], during their investigations concerning tropical marine mangrove fungi on three particular regions, i.e., the Indian Ocean (Seychelles), the Straits of Malacca (North Sumatra, Indonesia), and the South China Sea (Brunei) [
157,
158,
159]. These early studies were perhaps the driving force and what set up the research on palm fungi in the 1990s. After those first three taxonomical novelties, numerous further reports of new taxa were made on fronds of intertidal
N. fruticans, predominantly from Brunei, e.g., refs. [
105,
110,
117,
131,
160,
161,
162,
163,
164,
165] and Malaysia, e.g., refs. [
116,
143,
166,
167], which yielded more than thirty new species and seven new genera (
Table 1).
Nipa palm was found to be a “very distinct mangrove habitat” for fungal surveys, where a “largely distinct mycota” was found, including several “unique marine species belonging to genera consistently found on terrestrial palms” [
168]. Hyde [
110] observed that
Astrosphaeriella,
Linocarpon, and
Oxydothis, which are genera typically associated with terrestrial palm petioles in the tropics, were often associated with decaying intertidal Nipa palms. Not only was this mangrove palm tree important for systematic and descriptive taxonomy studies, but it was also a very important substratum for some of the first studies on the ecology of palm fungi, e.g., ref. [
7], as will be discussed later. Although most studies focused on the intertidal fungi occurring on
N. fruticans, studies have also been conducted in order to access the fungi inhabiting the aerial parts of this mangrove palm [
165].
Perhaps encouraged by those early records, during the last decade of the 20th century, Hyde et al. carried out an extensive survey of palmicolous fungi from a wide range of tropical palm species in different regions of the world (
Figure 1). The tropical regions of Southeast Asia and Australasia comprised the countries in which most of the collections were made, such as Brunei [
113,
115,
116,
117,
133,
134,
138,
154,
155,
160,
161,
162,
163,
164,
169,
170,
171], Indonesia [
109,
113,
115,
124,
126,
131,
133,
134,
172], Malaysia [
110,
116,
117,
118,
123,
131,
133,
134,
165,
167,
172], and Australia [
108,
109,
112,
114,
117,
133,
135,
138,
172]. Other regions and countries were moderately or occasionally sampled, including Thailand [
162], Papua New Guinea [
117,
123,
127,
131,
133,
173,
174,
175], Philippines [
117,
125,
167], Japan [
117,
133], Ecuador [
128,
138,
171], Brazil [
107], and the USA [
122,
124]. All the regions surveyed revealed the presence of an enormous diversity of fungi, among which, 22 genera and more than 80 species were described as new to science, several existing genera were re-examined, and some new combinations were proposed. In fact, one of the most remarkable outcomes of these early works by Hyde was the number of new genera introduced (
Table 1). Not only did they reveal the great untapped diversity of the fungal communities that inhabit palm trees in the tropics, but they also discovered a very particular unknown and underexplored taxonomic group of ascomycetes which make up what has been termed palm fungi (
Table 2). Therefore, this comprehensive research began to unveil and shape the community of palmicolous fungi, insomuch that it allowed to build up and consolidate the knowledge on the common mycota that typically inhabit the tissues of tropical and subtropical palms. According to Hyde [
111], fallen palm rachides and leaves in the tropics were found to be invariably colonised by fungi of the genera
Astrosphaeriella,
Linocarpon,
Oxydothis, and
Phomatospora. It later became clear that, in addition to
Oxydothis, several other genera of
Xylariales had common representatives on palm trees, including
Anthostomella,
Apioclypea,
Arecomyces,
Astrocystis,
Capsulospora,
Fasciatispora,
Nipicola, and
Pemphidium [
134,
138] (
Table 2).
Along with Hyde’s work on clarifying, redescribing, illustrating, and monographing existing genera [
105,
107,
111,
113,
117,
124,
126,
127,
130,
133,
171], these studies resolved certain taxonomic relationships within various important fungal families and orders. Moreover, they also expanded the knowledge of tropical mycology, especially with regard to the biodiversity of fungi that inhabit tropical hotspots, such as rainforests. Striking examples include the description of numerous new species in the genera
Linocarpon [
105,
154,
165,
172],
Oxydothis [
111,
112,
117,
156], and
Anthostomella [
133,
175], three of the most common genera found on palms (
Table 2); the clarification of certain unclear generic concepts for some of the genera, and their corresponding species, found on palms, including
Pemphidium [
107,
111,
135],
Guignardia [
126], and
Roussoella [
171,
176]; the description and discussion of some amphisphaeriaceous fungi occurring on palms, such as the genera
Fasciatispora [
124,
161],
Myelosperma [
113],
Seynesia [
127],
Arecophila [
131], and
Amphisphaeria [
136]; and the investigation of palmicolous fungi inhabiting the palms in tropical Australian, e.g., refs. [
112,
114], Bruneian, e.g., refs. [
170,
174], and Ecuadorian, e.g., refs. [
128,
171] rainforests. These first case studies in the early 1990s verified the existence of a well-represented set of morphological characters to describe these genera, as well as to distinguish them from other related genera, and to clarify the existence of new morphologically similar genera. As a result, it began to become clearer what the taxonomic placement of palm fungi was in higher taxonomic ranks, including different families of the
Xylariales and other less representative orders, e.g., ref. [
138] (
Table 2).
The large number of new genera and species reported on palms by Hyde in the early 1990s quickly prompted a growing interest in the mycobiota of these hosts and several of his students and collaborators, such as Fröhlich, Taylor, Aptroot, and Goh, worked to expand the knowledge of palmicolous fungi. Thus, in addition to the many new taxa recorded, different aspects of these fungi have been studied and have contributed to the fundamental knowledge of fungi and their biodiversity. This intensive research ultimately led to the description of at least three new families to accommodate genera that have been described and found to be common on palms, along with extant related genera, namely
Phaeochoraceae for
Cocoicola,
Phaeochora and
Serenomyces [
177],
Apiosporaceae for
Apiospora and
Appendicospora [
143], and
Myelospermaceae for
Myelosperma [
178].
Hyde, Fröhlich, Taylor, Aptroot, and Goh, studying ascomycetes developing on living, diseased, and dead palm material, surveyed different regions from East, South, and Southeast Asia, including, respectively, China (Hong Kong, Hubei, Hunan) [
6,
8,
129,
140,
142,
143,
144,
146,
147,
148,
149,
151,
179,
180,
181,
182,
183,
184,
185,
186], India [
152], and Brunei, Indonesia, Laos, Malaysia, Philippines, and Singapore [
6,
8,
122,
139,
140,
143,
147,
148,
149,
150,
153,
179,
181,
185,
187,
188,
189,
190,
191,
192,
193]. The Australasian regions were also intensively surveyed, including Australia [
6,
8,
121,
140,
141,
143,
144,
147,
148,
181,
185,
187,
189,
194,
195,
196,
197,
198,
199,
200,
201,
202,
203,
204] and Papua New Guinea [
150,
198]. Other regions and countries were also frequently or occasionally surveyed, including Ecuador (South America) [
6,
119,
143,
144,
150,
179,
181,
185,
205,
206,
207,
208], USA [
120], South Africa [
208,
209,
210], Seychelles [
8,
178,
181], and European countries, such as Switzerland and Great Britain [
8] (
Figure 1). As expected, the huge diversity of fungi collected from palm trees increased, with 22 new genera and more than 200 new species described. As a result, the importance of palm fungi began to become more evident as studies continuously revealed their broad taxonomic structure (
Table 2). Most of these studies and their outcomes have been compiled in the first book published by Hyde and co-workers in the Fungal Diversity Research Series concerning palmicolous fungi, which was entitled “Genera of ascomycetes from palms” and treated 100 selected genera of common fungi inhabiting the tissues of tropical palms [
104]. Moreover, another book on the same series, entitled “Palm microfungi”, has been published in the same year, which supplemented the previous information describing the ascomycetes found on palms during their comprehensive collections in Australia, Brunei, and Hong Kong [
6]. Remarkably, in addition to the ongoing revision of several genera of ascomycetes with common representatives on tropical palms, Fröhlich and Hyde [
6] described 65 taxa as new to science, including 3 new genera and an impressive number of 23 new species of
Oxydothis, considered to be the genus most commonly found on palms and invariably one of the earliest colonisers of dead palm leaves and fronds [
185,
211] (
Table 1 and
Table 2).
Following Hyde’s early studies of palm fungi in tropical rainforests, e.g., refs. [
112,
170,
171], one intensively studied region worth mentioning is the rainforests of North Queensland (Australia) (
Figure 1), where an immense diversity of fungi has been found in collections of different endemic palm species, such as
Archontophoenix alexandrae,
Laccospadix australasica,
Licuala ramsayi,
Linospadix microcaryus,
L. monostachyos, and
Oraniopsis appendiculata, e.g., refs. [
121,
140,
141,
143,
145,
147,
148,
181,
185,
187,
189,
194,
195,
196,
197,
198,
199,
200,
202]. While most of the studies conducted by Hyde and co-workers were dedicated to surveying saprobic fungi that develop on palm trees, some of these studies in the North Queensland rainforest were conducted to survey palm phytopathogens, particularly those parasitic on palm leaves and causing leaf spot diseases. Palm phytopathogens were also documented in studies conducted during an expedition of the British Mycological Society to Ecuador in August 1993 to gather fungi developing on palms in the rainforest of Cuyabeno [
119,
143,
144,
150,
181,
185,
206,
207,
208]. Moreover, Hyde and Cannon [
212] monographed members of the
Catabotrydaceae,
Phaeochoraceae, and
Phyllachoraceae families that occur in association with tar spots on palms. In addition to treating these families and their placement in the fungal classification, several genera were treated and analysed, some were re-introduced based on palm collections, and four genera and three species were described as new to science (
Table 1). Some of the genera discussed by Hyde and Cannon [
212] represent important taxa in the phytopathogenic mycobiota of palm trees, such as the genus
Serenomyces, a group of mostly biotrophic fungi apparently known only in association with members of
Arecaceae [
213,
214].
Although some palm phytopathogens have been identified, most studies on palmicolous fungi in the last decade of the 20th century focused on the myriad of saprobic fungi that inhabit the palms from tropical rainforests, where palm litter is a major component. Samples were collected from many different palm trees that inhabit the tropics, including from typical
Phytelaphas spp. of Central and South America to typical
Licuala spp. of Southern China (
Figure 1). Palm material, such as senescent and dead petioles, rachides, stems, fronds, and decaying and dead trunks, yielded plentiful novelties (
Table 1). One interesting palm tree worth mentioning is
A. alexandrae, an endemic palm from Queensland, Australia. Studies on the palmicolous fungi that inhabit the tissues of this palm have perhaps been the driving force for studying other questions about the taxonomy and ecology of palm fungi, as well as extending the research to fungi associated with palms from non-tropical habitats. In fact, its endemic nature and relative geographic isolation make
A. alexandrae an ideal substratum for studying host-specificity and fungal biogeography, which have been identified as important factors for the description of many novel palmicolous fungi when hosts are studied in their natural environment [
142,
194]. Likewise, the description of the new palmicolous genus
Cannonia [
204] collected from
Trachycarpus fortunei in Australia, outside its native temperate range in parts of China and some neighbouring countries (
Table 1), also raised questions about the importance of studying the biogeography of palm fungi. As a result, studies on palmicolous fungi that expanded their geographical boundaries and temperate palms also began to be surveyed [
215].
The constant description and illustration of new taxa found on palm trees has often led to discussions of their placement in the fungal classification. As a result, several families, and their respective genera, with common representatives on palms, have been extensively treated and the taxonomy of palmicolous fungi, at least in the tropics, has become clearer, e.g., refs. [
6,
8,
104,
143,
150,
171,
178,
181,
185,
212,
216,
217] (
Table 2). Noteworthy examples include the description and discussion of common genera that occur on palms with the introduction of several new species, such as
Nipicola [
137],
Nectria, and allied genera [
207],
Massarina [
139,
149],
Anthostomella [
143,
209,
218,
219,
220,
221],
Astrosphaeriella [
144,
148], and
Neolinocarpon [
140], whose geographical distribution has widened considerably throughout Australasia, Southeast Asia, South America (Ecuador), and China (including Hong Kong). Other common genera discussed, with the introduction of new species, include
Dictyosporium [
184] and
Lasiosphaeria and similar genera, such as
Chaetosphaeria,
Iodosphaeria, and the newly described genus
Arecacicola [
146,
181,
185] (
Table 1). Likewise, some doubtful species described for some genera that occur on palms have been clarified and revised, namely in
Mycosphaerella and
Sphaerella, which are common phytopathogens [
145],
Didymosphaeria and similar taxa, which include truly terrestrial ascomycetes with ascospores with appendages described from palms [
147,
222,
223], and genera of unitunicate ascomycetes with apiospores, which are frequently recorded on palms, such as
Anthostomella, Apioclypea,
Apiospora,
Appendicospora, and the new described genera
Brunneiapiospora and
Palmaria (as
Palmomyces) [
143] (
Table 1). Several xylariaceous genera recorded on palms have also been treated based on herbarium specimens and fresh material collected during the investigation on palmicolous fungi, which led to the description of several new species, for example, of the genus
Astrocystis [
150].
Following the incredible contribution to the knowledge of fungal biodiversity and their taxonomy, these studies have also made it possible, on several occasions, to discuss and even uncover some links between sexual and asexual morphs [
150,
167,
181,
203,
208,
224]. The importance of studying “anamorph-teleomorph” connections was strongly emphasised in the last compilation book published by Taylor and Hyde [
8] in the Fungal Diversity Research Series on palm fungi, entitled “Microfungi of tropical and temperate palms”. As well as continuing the previous studies on palm fungi carried out on tropical rainforest palms, Taylor and Hyde [
8] also studied fungi associated with palms in non-rainforest habitats in the tropics and palms in temperate habitats. Interestingly, this is the first book to deal not only with the identification of palmicolous fungi but also with the extent of their diversity and the factors that affect it, which reflects the complexity of the investigation carried out by Hyde and co-workers and the diversity of the approaches that were used. A total of thirty-four new species, including four new species of
Anthostomella, and one new genus,
Tribulatia, were described (
Table 1), which is a surprising number considering the extent of the sampling and the number of taxa already described from palms by Hyde and co-workers.
The studies on palmicolous fungi in tropical regions continued, with other regions beginning to be surveyed more systematically, including many reports from additional Hyde co-workers, namely McKenzie, Pinnoi, Pinruan, and Yanna. Although the description of new taxa remained the main objective of these studies, which yielded more than thirty species and seven genera described as new to science (
Table 1), they have also begun to provide ecological data on the communities of tropical palmicolous fungi that inhabit some tropical palm tree species, such as
Livistona chinensis and
Phoenix hanceana, e.g., refs. [
13,
14,
15]. Yanna et al. [
224,
225,
226,
227,
228,
229] described several new species from different palm tree species in Hong Kong, comprising the typical
Ascomycota assemblage commonly found on palms, particularly species of
Appendicospora [
225], to atypical hyphomycetes and coelomycetes genera, such as
Koorchaloma [
226],
Staurophoma [
224],
Endomelanconium [
227], and
Everhartia [
229].
Although the initial studies on palms focused mainly on the evaluation of the
Ascomycota coverage through the presence of its sexual morphs on the host, the evident potential of these hosts for biodiversity surveys has led to a diversification of approaches and, consequently, discoveries. As a result, a considerable number of reports began to describe several new species and genera of palmicolous hyphomycetes, particularly dematiaceous hyphomycetes, in addition to the usual sexual morphs of ascomycetes from well-studied regions, such as Australia (North Queensland) [
15,
189,
201,
202,
230], China (Hong Kong and Hainan) [
182,
183,
184,
231,
232,
233,
234,
235], Brunei, and Thailand [
166,
188,
189,
190,
228,
236,
237,
238] (
Figure 1). In addition to hyphomycetes, species of the discomycetes genus
Lachnum, new species of which are often found on palm trees [
239,
240], have also been reported in tropical China, e.g., ref. [
231].
Just before the input of molecular data began to broaden and strengthen the research of Hyde and co-workers on palm fungi, McKenzie, Pinnoi, and Pinruan et al., studying the fungal diversity on palms from the Sirindhom Peat Swamp Forest at Narathiwat, Southern Thailand, found and described several new palmicolous taxa [
236,
241,
242,
243,
244,
245,
246,
247]. Thus, in addition to new species of the typical
Ascomycota assemblage reported from palm tissues in the tropics, including new species of
Submersisphaeria [
247],
Jahnula [
242], and the new genus
Unisetosphaeria [
245] (
Table 1), some new palmicolous “anamorphs” were introduced, including new species of
Chalara [
241],
Dactylaria [
245],
Custingophora,
Vanakripa [
246],
Craspedodidymum [
243], and
Stachybotrys [
244]. Moreover, following the studies carried out by Yanna et al., ecological data on tropical palm fungal communities have also been documented in the peat swamp palms
Eleiodoxa conferta [
248] and
Licuala longicalycata [
249], as well as in
Calamus spp. [
16].
2.1.3. The Palmicolous Hyphomycetes from Central American Countries
A perusal of the available literature on palm fungi reveals that, to date, no intensive studies have been carried on palmicolous “anamorphs”, i.e., hyphomycetes and especially coelomycetes. However, it is worth mentioning a few scattered studies that reported new species and genera of palmicolous hyphomycetes. These studies have shown that the predominant group of “anamorphs” in palm litter are the dematiaceous hyphomycetes. In fact, one of the first consistent reports of palmicolous hyphomycetes is that of Ellis in the 1960s and 1970s, who reported several new species and some new combinations based on collections of palms from West Africa, Southeast Asia, and Central America in his series of papers and books about “Dematiaceous hyphomycetes” [
29,
30,
31,
32,
250,
251,
252,
253,
254,
255,
256,
257,
258].
Table 1. Genera described as new to science and found on Arecaceae hosts in the last three decades.
| Genus |
Type Species |
Host |
Country/Region |
Sequence Data 1 |
Reference |
| Acarocybellina |
A. arengae |
On a dead leaf of Arenga engleri |
Japan |
N/A |
[259] |
| Acarocybiopsis |
A. cubitaensis |
On a dead trunk of Roystonea regia |
Cuba |
N/A |
[260] |
| Acuminatispora |
A. palmarum |
On decaying petioles and rachides of an unidentified palm in mangrove |
Thailand |
A |
[261] |
| Agrabeeja |
A. kavakapriya |
On synnemata of Melanographium citri on a rachis of Korthalsia grandis |
Singapore |
N/A |
[262] |
| Allodiatrype |
A. arengae |
On a dead petiole of Arenga pinnata |
Thailand |
A |
[263] |
| Anabahusakala |
A. amapensis |
On decaying leaves of Syagrus sp. |
Brazil (Amapá) |
N/A |
[264] |
| Anisospadicoides |
A. macrocontinua
(as Spadicoides macrocontinua) |
On a rotten petiole of an unidentified palm |
Peru |
N/A |
[64,265] |
| Apioclypea |
A. livistonae |
On a rachis of Livistona sp. |
Papua New Guinea |
N/A |
[175] |
| Apogaeumannomyces |
A. perplexus |
On a decaying frond of an unidentified palm |
Peru |
N/A |
[67] |
| Appendicospora |
A. coryphae |
On dead rachides of Corypha elata |
Philippines |
N/A |
[125] |
| Appendispora |
A. frondicola |
On a dead rachis of Oncosperma horridum on forest floor |
Brunei |
N/A |
[115] |
| Arecacicola |
A. calami |
On a trunk of Calamus sp. |
Indonesia (Java) |
N/A |
[185] |
| Arecomyces |
A. frondicola |
On a rachis of Arenga undulatifolia |
Brunei |
N/A |
[138] |
| Arecophila |
A. gulubiicola |
On a dead trunk of Gulubia costata |
Papua New Guinea |
N/A |
[131] |
| Ashtaangam |
A. Sundaram |
On a rachis of an unidentified palm |
Malaysia |
N/A |
[266] |
| Astrosphaeriellopsis |
A. bakeriana |
On a petiole of Borassus sp. |
Thailand |
A |
[267] |
| Asymmetricospora |
A. calamicola |
On a dead stem of Calamus caryotoides |
Australia (Queensland) |
N/A |
[141] |
| Atrosetaphiale |
A. flagelliformis |
On a decayed petiole of an unidentified palm |
Peru |
N/A |
[65] |
| Aunstrupia |
A. nodipes |
On rotten and dead leaves and rotten petiole and branches of unidentified palms |
China (Guangdong) |
A |
[268] |
| Bacusphaeria |
B. nypae |
On a petiole base of Nypa fruticans |
Malaysia |
A |
[269] |
| Baipadisphaeria |
B. spathulospora |
On a trunk of Licuala longicalycata submerged in peat bog |
Thailand |
A |
[270] |
| Basauxia |
], and Texas (USA) [
571], in addition to other exclusively morphological studies that have reported
Hermatomyces species from palm trees collections in India [
572] and Sierra Leone [
573] (
Figure 1,
Table 2). More recently, Konta et al. [
285] described
Helminthosporium-like taxa from palms in Thailand and introduced two new genera in
Massarinaceae, viz.
Haplohelminthosporium and
Helminthosporiella (
Table 1). Konta et al. [
285] also provided a checklist for
Helminthosporium reported worldwide and most
Helminthosporium species described from palms were based on morphology alone and only one species,
H. livistonae on leaves of
Livistona australis from Australia [
302], was based on both morphology and sequence data. Chen et al. [
574] also introduced the new species
H. chinense on a decaying branch of an unidentified palm tree from China.
Many other occasional reports of palmicolous “anamorphs”, including new genera and species, are continuously published, revealing the plethora of genera that make up the hyphomycetous and coelomycetous assemblage that inhabit palm tissues, e.g., refs. [
268,
279,
310,
575,
576,
577,
578] (
Table 2). For example, Hongsanan et al. [
279] described the new genus and species
Discopycnothyrium palmae on the branches of an unidentified palm from Thailand (
Table 1). A number of new hypocrealean members have been recently introduced based on palm collections from French Guiana, including species of
Chaetopsina [
579],
Clonostachys [
580,
581],
Hydropisphaera [
582],
Ijuhya [
583,
584],
Lasionectria [
585], and
Volutella [
586]. Several novel anamorphic chaetosphaereaceous fungi, including new genera, were reported from palm collections from China [
268,
575,
577,
578] and Thailand [
310] (
Table 1).
Crane and Miller [
587] introduced some new species in
Torula-like genera from palms, such as
Bahusaganda and
Bahusandhika, but no molecular data was obtained to establish their natural placements. Although the importance of acquiring sequence data to identify fungal taxa is now widely recognised and advisable [
588,
589,
590], there are still several publications that have been and continue to be published without the support of molecular methods, introducing new morphological palmicolous taxa. For example, Wulandari et al. [
591], while synopsising the species of
Phyllosticta (as
Guignardia) described from palm trees, introduced two new species from Northern Thailand based on morphology. Lechat and Fournier [
592] introduced the new species
Lasionectria marigotensis on a decaying leaf of
Cocos nucifera from Guadeloupe (French West Indies) by comparing its morphology with a previous collection of
Lasionectria,
L. calamicola, from palms in Australia and Brunei Darussalam by Fröhlich and Hyde [
6]. Later, the new species
Dictyocheirospora indica (as
Dictyosporium indicum) was collected on a petiole of
Phoenix rupicola from India [
593] and the new species
Endophragmiella licualae was collected on dead branches of
Licuala fordiana from China [
594]. More recently, several palmicolous fungi, including new species, have been reported from the rainforests of the Andaman-Nicobar Islands (India) by Niranjan and Sarma [
453,
454,
455,
457] based on morphology alone, including important palmicolous genera whose taxonomic resolution highly relies on DNA sequence data, such as members of
Aigialaceae, viz.
Fissuroma and
Neoastrosphaeriella, and
Astrosphaeriellaceae, viz.
Astrosphaeriella. Likewise, as already mentioned, many new species and genera of palmicolous hyphomycetes have been described on the basis of their morphology in Central and South American countries, including Mexico, Argentina, and Brazil.
The consistent new discoveries of fungi from palm hosts suggest their considerable potential for the identification of novel fungal taxa. Thus, using morphology alone to introduce new palmicolous taxa is strongly discouraged here. Although many fungal species have unique and distinctive characters that make them easily identifiable, especially in genera of hyphomycetes, the existence of cryptic species and species complexes has highlighted the importance of molecular methods in fungal identification [
590,
595]. In fact, the morphological species concept is thought to underestimate the number of species, since morphological characters can be very plastic (phenotypic plasticity) and often defines groups of cryptic species [
590].
From Aquatic to Phytopathogenic Fungi: The Broad Taxonomic Spectrum of Palm Fungi
Freshwater and marine or mangrove ecosystems have also revealed a remarkable diversity of new palmicolous taxa with the incorporation of phylogenetics into the previous regular morphological studies [
9,
261,
269,
278,
596,
597,
598,
599,
600,
601,
602,
603,
604]. The mangrove palm
Nypa fruticans has proven to be a very distinctive habitat for fungal research since 1988, when Hyde began his studies to understand the assemblage of fungi that colonise palm tissues [
154,
162]. In fact, many new palmicolous fungi are continuously being described from
N. fruticans. For instance, Zhang et al. [
261] introduced the new genus
Acuminatispora isolated from decayed petioles and rachides of palms in mangrove habitats in Thailand, including
N. fruticans and
Phoenix paludosa. More recently, Jones et al. [
604] described the new species
Salsuginea phoenicis on a decaying petiole of
P. paludosa from the intertidal zone in Thailand. Thus, mangrove and peat swamp palms have been the substratum for the collection of various new aquatic fungi (freshwater and/or marine ascomycetes), whose phylogenetic resolution has often led to the description of new families and genera. In fact, four families have been recently introduced to accommodate new and extant taxa of aquatic fungi with affiliations to palm habitats.
Falciformispora and
Trematosphaeria, with representatives found on palms, were assigned to the new family
Trematosphaeriaceae [
596,
598]. Suetrong et al. [
599] introduced the new family
Tirisporellaceae in
Diaporthales to accommodate the genera
Tirisporella and
Thailandiomyces collected from palms in freshwater streams or peat swamps, and Abdel-Wahab et al. [
269] added the new genus
Bacusphaeria isolated from the petiole base of
N. fruticans from Malaysia. The new family
Dictyosporiaceae was established to accommodate saprobic fungi that occur on decaying wood and plant debris in terrestrial and freshwater habitats, including
Dictyosporium, often found on palm trees, and the new genus
Dictyopalmispora described from decaying leaves of
Licuala longecalycata in a peat swamp forest in Thailand [
278,
602,
603] (
Table 1). Later, Zhang et al. [
9] introduced another new pleosporalean family,
Striatiguttulaceae, to accommodate two monophyletic lineages described from decayed rachides of
N. fruticans and
P. paludosa, the new genera
Longicorpus and
Striatiguttula.
In the last decade, microfungi that occur as saprobes, pathogens, and endophytes on palm hosts are commonly introduced and reanalysed in mycological series, such as Fungal Diversity notes, e.g., refs. [
296,
605,
606,
607,
608], Fungal Planet description sheets, e.g., refs. [
609,
610,
611,
612,
613,
614,
615,
616], and Mycosphere notes, e.g., refs. [
617,
618,
619,
620], as well many other similar publications, e.g., refs. [
621,
622,
623,
624,
625,
626,
627,
628,
629]. A remarkable taxonomic novelty published recently based on a palm collection was the new order
Pararamichloridiales introduced to accommodate the new family
Pararamichloridiaceae based on the new genus and species
Pararamichloridium livistonae on leaves of
Livistona australis from New South Wales, Australia [
298]. Therefore, these occasional publications continue to expand both the broad taxonomic spectrum of palm fungi and the importance of studying them as phytopathogens and other ecological groups. In fact, in recent years, a series of new palmicolous phytopathogens have been introduced and have highlighted the lack of knowledge about the ecology of palmicolous fungi. For instance, the new species
Cercospora arecacearum has been found associated with necrotic leaflets of areca palms in Thailand by To-anun et al. [
630]. Kinge and Mih [
631] described the new basidiomycete
Ganoderma ryvardenii (as
G. ryvardense) associated with basal stem rot disease of oil palm in Cameroon, one of the main production constraints faced by agro-industries and smallholders’ farmers in oil palm production. Moreover, Mbenoun et al. [
632] described the new species
Thielaviopsis cerberus (as
Ceratocystis cerberus) on the stump of a felled
Elaeis guineensis tree from Cameroon, while studying species boundaries in the
C. paradoxa complex, a serious constraint to the cultivation of monocotyledonous crops, including the trunk rot affecting almost all palm species. Considerations on the importance of these diseases as one of the main causes of losses in oil and ornamental palms have been put forward by Aiello et al. [
633], while introducing the new phytopathogen species
Ilyonectria palmarum. More recently, the new genus and species
Palmeiromyces chamaeropicola was collected from diseased foliage of
Chamaerops humilis from Portugal [
295], revealing a new insight into
Teratosphaeriaceae leaf diseases, which are caused by important phytopathogens of various plant hosts.
Research into palm fungi continuously reveals the importance of the Arecaceae family as host plants to search for novel taxa. In addition, recent studies have been important in showing the imperative need to apply molecular data to resolve the taxonomic structure of palm fungi as an ecologically diverse and important assemblage.
2.2. History of Biodiversity and Ecological Studies on Palm Fungi
Palm trees have proven to be a diverse habitat, exhibiting intense fungal colonisation. Most studies on microfungi that colonise palms are taxonomic, insomuch as they have primarily focused on cataloguing fungi and describing new taxa collected on palm substrata from various regions of the world, especially in the tropics (Figure 1). However, there are a number of studies on the ecology of palm fungi, which have focused on different aspects of their biodiversity, mainly with regard to saprobes and endophytes. Although the approach of these studies is different, the description of new taxa has often resulted from initially ecological approaches that yielded several interesting fungi for further analysis. This suggests that both approaches are essential to explore the knowledge on fungi and their biodiversity. In fact, the implications of biodiversity and ecological data on the global numbers of fungi and their general knowledge have been pointed out, which will be discussed later in relation to palm fungi and their key role in biodiversity surveys.
A review of the literature on the ecology of palm fungi reveals that studies have been conducted over the past three decades. These have mainly resulted from the intensive research carried out by Hyde and his co-workers. They have made substantial contributions to knowledge of both the taxonomy and biodiversity of palm fungi, not only by identifying the fungal assemblage that occur on palms, but also by exploring the extent of their diversity and the factors that affect it. However, some ecological studies have also been carried out on Indian palm fungi, as well as palm fungi from Central American regions. A historical account of research into the ecology and biodiversity of palm fungi is presented herein.
Fröhlich and Hyde [
10] studied the biodiversity of palm fungi in the tropics, forecasting that the estimate of 1.5 million species would be a “very conservative estimate of the number of fungal species extant on the planet”. Taylor et al. [
12] investigated the biogeographical distribution of microfungi from temperate and tropical palms. They revealed that differences in fungal assemblages were more related to climatic influences than to the hosts sampled, as well as to the status of these hosts at the site sampled. Subsequently, Yanna et al. [
13,
14,
634] assessed the composition of palm fungal communities and their succession over time, pointing out that differences in fungal assemblages could be related to different collection sites, hosts, stages of decomposition, and tissues sampled. Later, Pinnoi et al. [
16] studied saprotrophic fungal communities associated with
Calamus spp. and reported differences in the assemblages of fungi inhabiting different microhabitats and tissues, with dry petioles supporting a greater diversity of species.
Several studies have been dedicated to endophytic palm fungi, which was one of the first ecological issues to be investigated in palm fungi [
10,
11,
468,
469,
472,
473,
635,
636,
637,
638]. These studies often report significant differences in the number of isolates and the taxonomic composition of fungi in respect to the plant growth stages, season, site, and tissues sampled, not only in tropical palms, but also in temperate palms, such as
Trachycarpus fortunei [
215]. Even so, research into palm endophytes, which began in the early 1990s, was primarily motivated by the lack of knowledge about endophytes in plants from tropical regions when compared with those in plants from temperate regions [
472,
473].
The first study on palm endophytes was by Rodrigues and Samuels [
469], who documented the occurrence of endophytes inhabiting the leaves of the Australian fan palm
Licuala ramsayi, pointing out preliminary differences in the endophyte assemblages recovered from different parts of
L. ramsayi leaves. In addition, the existence of a fungal assemblage composed mainly of xylariaceous fungi was documented. This was also later documented by Rodrigues and her colleagues when they published the first studies on the fungal endophytes that inhabit the foliage of the Amazonian palm
Euterpe oleracea from Combu Island (Belém, Pará). In fact, some of these studies were based on systematic and descriptive taxonomy, particularly for xylariaceous species commonly recorded as endophytes of palm tissues, including
Xylaria and
Idriella [
291,
470,
471]. However, a comprehensive ecological approach was conducted by Rodrigues [
468], who found that colonisation of
E. oleracea by endophytes was positively correlated with leaf age, plant growth stages, site, and season, as well as with the interactive effect of some of these factors. Later, a similar study was carried out by Fröhlich et al. [
11], who investigated the endophyte communities inhabiting different parts of the fronds of
L. ramsayi individuals from Australia and Brunei. They noted differences between the fungal assemblages of different palm tissues and tissues with different ages. Likewise, Taylor et al. [
215] observed differences in the endophytic communities recovered from different tissues of the frond of the temperate palm
Trachycarpus fortunei. They also documented important information regarding the effect of climate on endophytes assemblages by analysing individuals growing inside and outside their natural geographical range.
The importance of these studies and their implications for fungal biology and biodiversity rapidly increased interest in unveiling more details of the ecology of palmicolous endophytes. Molecular data began to be applied in such studies and made endophytes one of the main issues of palm fungi to be explored until recent years [
523,
639,
640,
641,
642,
643,
644,
645,
646,
647,
648]. Subsequently, the array of questions has diversified greatly. While some studies have invested in documenting the maximum possible diversity of palm endophytes communities, others have targeted particularly important palms and the impacts that endophytic communities can have on their pests and diseases. For instance, Guo et al. [
639,
640,
641] tried to decrease the percentage of sterile mycelium that often lacks identification in endophyte studies by using both morphology and rDNA sequences in a study of endophytes on
Livistona chinensis fronds. Later, Rungjindamai et al. [
643] and Pinruan et al. [
523] used both morphology and molecular techniques to characterise basidiomycete endophytes isolated from the leaves, rachides, and petioles of the oil palm
Elaeis guineensis, and to reflect on how these fungi can be used as a biocontrol management strategy against the palm pathogen
Ganoderma boninense. Similarly, Mahmouda et al. [
644] examined the endophytic fungal diversity associated with the roots of the date palm
Phoenix dactylifera growing in coastal dunes to generate a collection of strains that can be used as biocontrol agents against date palm root diseases. More recently, Azuddin et al. [
646] studied the fungal endophytes on the spines of
Calamus castaneus and evaluated their antagonistic activity against phytopathogens.
Although the importance of palm pathogens and their management are often mentioned, particularly in palms that are important in international trade, relatively few studies have focused on these fungi and their ecology, e.g., refs. [
199,
212]. Most publications on palmicolous pathogens have been motivated by systematic and descriptive taxonomic studies that have identified new taxa associated with palm diseases, e.g., refs. [
187,
195,
196,
197,
198,
208,
295,
556,
630,
631,
632,
633]. In fact, although the main fungal diseases that affect palm trees have virtually all been identified, e.g., refs. [
649,
650,
651] little is known about minor pathogens, including those that cause leaf spots, as shown by the investigation of Hyde and co-workers. Recently, Douanla-Meli and Scharnhorst [
551], while describing botryosphaeriaceous taxa associated with palm foliage from Mexico, reflected on the risk these taxa may pose to temperate countries, since palm foliage can be an import route for potentially phytopathogenic fungi. In addition, a number of studies have addressed the identification of fungal pathogens associated with
P. dactylifera in regions where this palm is an important agricultural crop, e.g., refs. [
652,
653,
654,
655,
656].
Ecological studies on palm fungi are often directed at palm species that are important world crops in international trade. For example, Asensio et al. [
657] investigated the mycobiota of the phylloplane of the date palm
P. dactylifera and their interactions. Kirkman et al. [
658] studied the diversity and ecological association of the oil palm
E. guineensis fungal microbiome across root, rhizosphere, and soil compartments, while Seephueak et al. [
659] studied the diversity of microfungi that occur in different tissues of the oil palm frond litter in a plantation in Southern Thailand. Very few studies on palm fungi have explored fungal diversity in roots and, as a result, palm root fungi are virtually unknown. However, few ecological studies on palm fungi have addressed the biodiversity of arbuscular mycorrhizal fungi (AMF) and reflected on the importance of these symbiotic associations to the health and growth of some palm species, such as
Attalea speciosa [
660],
Coccothrinax spp. [
661,
662,
663],
Cocos nucifera [
664],
Desmoncus orthacanthos [
665],
Metroxylon sagu [
666], and
P. dactylifera [
667,
668].
Many ecological studies have focused on the biodiversity of fungi from peat swamp [
248,
249,
669,
670] and mangrove palms [
7,
162,
671,
672,
673,
674,
675]. Such studies often reflect on the assemblage of freshwater and marine fungi that inhabit palm tissues, compared with the fungi typically associated with terrestrial palms. In addition, the presence of different fungal assemblages from different microhabitats and palm trees tissues is often addressed. For instance, Pinnoi et al. [
248], studying the fungal biodiversity of
Eleiodoxa conferta, reported that fungi are more abundant on the petioles of wet palm material. Similar ecological patterns were reported by Pinruan et al. [
249], who indicated that the petioles of dry material of
Licuala longicalycata supported the most diverse fungal communities. Furthermore, questions regarding host- and tissue-specificity are frequently addressed, particularly in studies on the brackish water palm
Nypa fruticans, whose colonisation by fungi has been well documented [
671,
674]. Hyde and Alias [
7] reported differences in the fungal composition of different palm structures of
N. fruticans, including leaves, leaf veins, rachides, petioles, and inflorescences, collected from intertidal and terrestrial habitats. Likewise, Hyde and Sarma [
672] reported differences in the fungal assemblages inhabiting
N. fruticans along a river, addressing some ecological observations regarding the horizontal and vertical distribution of fungi. One of the most recent publications in the ecology of palmicolous fungi reported the co-occurrence of certain species of fungi on
N. fruticans. This study revealed some aspects related to the structure of the fungal communities on tissues of the brackish water palm and helped to understand the dynamics of the ecosystem, suggesting, for example, the potential interaction established between the fungi of these communities [
675].
There are numerous studies on palm fungi and a plethora of data has been collected on different aspects of their biodiversity—from their taxonomic and systematic structure to ecological traits of their lifestyles inhabiting different palm tissues, microhabitats, and geographic regions. However, review studies on palm fungi are scarce. Most are confined to proceedings of mycological conferences or chapters in books that explore broader themes, e.g., refs. [
17,
676,
677,
678]. With the advent of DNA sequencing, information on palm fungi, particularly their taxonomy and systematics, has become considerably more complex. Thus, defining or understanding them as a complex and diverse group of fungi may not be an easy task.
Table 2. Synopsis of the taxonomic structure of palm fungi: genera and respective families in subclasses of Sordariomycetes and Dothideomycetes with common representatives found on Arecaceae hosts.
| Class |
Subclass |
Order |
Family |
Genera |
| Dothideomycetes |
Dothideomycetidae |
Dothideales |
Dothideaceae |
Uleodothis |
| |
|
Mycosphaerellales |
Extremaceae |
Castanedospora |
| |
|
|
Mycosphaerellaceae |
Cercospora, Distocercospora, Exosporium, Pallidocercospora, Passalora, Phaeophleospora, Pseudocercospora, Ramularia, Scolecostigmina, Uwemyces, Zasmidium |
| |
|
|
Teratosphaeriaceae |
Palmeiromyces, Stenella |
| |
|
|
Incertae sedis |
Pseudoepicoccum |
| |
Pleosporomycetidae |
Acrospermales |
Acrospermaceae |
Gonatophragmium |
| |
|
Pleosporales |
Acrocalymmaceae |
Acrocalymma |
| |
|
|
Aigialaceae |
Fissuroma, Neoastrosphaeriella |
| |
|
|
Arthopyreniaceae |
Mycomicrothelia |
| |
|
|
Astrosphaeriellaceae |
Astrosphaeriella, Astrosphaeriellopsis, Pteridiospora, Pithomyces, Javaria, Triseptatospora, Xenoastrosphaeriella |
| |
|
|
Coniothyriaceae |
Coniothyrium |
| |
|
|
Corynesporascaceae |
Corynesporasca |
| |
|
|
Delitschiaceae |
Delitschia |
| |
|
|
Dictyosporiaceae |
Dictyocheirospora, Dictyopalmispora, Dictyosporium, Pseudocoleophoma, Sporidesmiella |
| |
|
|
Didymosphaeriaceae |
Didymosphaeria, Montagnula, Paraconiothyrium, Paraphaeosphaeria, Pseudopithomyces |
| |
|
|
Hermatomycetaceae |
Hermatomyces |
| |
|
|
Lentimurisporaceae |
Bahusandhika |
| |
|
|
Leptosphaeriaceae |
Chaetoplea, Leptosphaeria, Quasiphoma |
| |
|
|
Lindgomycetaceae |
Lolia |
| Dothideomycetes (cont.) |
Dothideomycetidae (cont.) |
Pleosporales (cont.) |
Lophiostomataceae |
Lophiostoma, Vaginatispora |
| |
|
|
Massarinaceae |
Haplohelminthosporium, Helminthosporiella, |
| B. pulchra |
On a rachis of an unidentified palm |
Malaysia |
N/A |
[ | 266] |
| Helminthosporium | , | Massarina |
| |
|
|
Melanommataceae |
Asymmetricospora, Byssosphaeria, Camposporium, Herpotrichia |
Bhadradriella |
B. hyalina |
On fallen pods of Roystonea regia |
India (Andhra Pradesh) |
N/A |
[271] |
| Brachysporiopsis |
B. chinensis |
On a decaying rachis of Livistona chinensis |
China (Hong Kong) |
N/A |
[228] |
| Brobdingnagia |
B. nigeriensis |
| |
|
|
Morosphaeriaceae |
Helicascus |
| |
|
|
Neophaeosphaeriaceae |
Neophaeosphaeria |
On tissues of Calamus sp. |
Nigeria |
N/A |
[212] |
| |
|
|
Occultibambusaceae |
Brunneofusispora, Neooccultibambusa |
Brunneiapiospora |
B. javensis |
| | On a rachis of | Calamus sp. |
Indonesia (Java) |
N/A |
[143] |
| |
|
Periconiaceae |
Periconia |
Bulbocatenospora |
B. complanata |
On fallen leaves of Bactris setulosa |
Venezuela |
| |
| N/A |
[ | 272 | ] |
| |
Phaeosphaeriaceae |
Amarenographium | , | Parastagonospora, Phaeosphaeria, Septoriella, Wojnowiciella |
Cannonia |
C. australlis |
On rotten branches of Butia yatay |
Argentina |
N/A |
[204] |
| |
|
|
Pleosporaceae |
Bipolaris, Curvularia, Exserohilum |
Capsulospora |
C. frondicola |
On a rachis of Daemonorops |
| | sp. |
| Brunei |
N/A |
[134] |
| |
Pseudoastrosphaeriellaceae |
Carinispora | , | Pseudoastrosphaeriella |
Carinispora |
C. nypae |
On decaying intertidal fronds of Nypa fruticans |
Brunei |
N/A |
[162 |
| |
| ] |
| |
Pseudoberkleasmiaceae |
Pseudoberkleasmium |
Castanedospora |
C. pachyanthicola |
On the petiole of a dead leaf of |
| | Sabal palmetto |
| USA (Florida) |
A |
Roussoellaceae |
Appendispora, [273] |
| Neoroussoella | , | Roussoella |
Caudatispora |
C. palmicola |
On a dead rachis of Phytelaphas |
| | Ecuador |
N/A |
[ | 119 | ] |
| |
|
Salsugineaceae |
Salsuginea |
Cenangiumella |
C. rattanicola |
On a dead rattan sheath of Calamus conirostris |
Brunei |
N/A |
[6 |
| |
| ] |
| |
Striatiguttulaceae |
Longicorpus | , | Striatiguttula |
Chitinasiproducens |
C. palmae |
|
|
|
|
| |
|
|
Trematosphaeriaceae |
Falciformispora, Trematosphaeria |
Circinoconiopsis |
C. amazonica |
On decaying leaves of Oenocarpus sp. |
Brazil (Pará) |
N/A |
[274] |
| |
|
|
Teichosporaceae |
Parateichospora |
Cocoicola |
C. cylindrospora |
On petioles of Cocos nucifera |
Papua New Guinea |
N/A |
[123] |
| |
|
|
Tetraplosphaeriaceae |
Ernakulamia, Tetraploa |
Corynesporasca * |
C. caryotae |
On rotting leaves of Caryota urens |
Sri Lanka |
N/A |
[275 |
| |
| ] |
| |
Torulaceae |
Cylindrotorula | , | Torula |
Curvatispora |
C. singaporensis |
On a fallen decaying frond of |
| |
| Livistona spinosa |
Singapore |
N/A |
|
Incertae sedis |
Acuminatispora, Plectophomella, Repetophragma[153] |
| Cyanopulvis |
C. australiensis |
On a dead rattan of Calamus australis |
Australia (Queensland) |
| |
| N/A |
[ | 6 | ] |
| Hysteriales |
Hysteriaceae |
Gloniopsis |
Cylindrotorula |
C. indica |
On a decaying spathe of Cocos nucifera |
India (Maharashtra) |
A |
[276] |
| |
Incertae sedis |
Asterinales |
Asterinaceae |
Asterina, Cirsosia, Discopycnothyrium |
Diabolocovidia |
D. claustri |
On leaves of Serenoa repens |
USA (Florida) |
A |
[277] |
| |
|
|
Lembosiaceae |
Lembosia |
Dictyopalmispora |
D. palmae |
On decaying leaves of Licuala longicalycata |
Thailand |
A |
[278] |
| Discopycnothyrium |
D. palmae |
| |
|
|
Morenoinaceae |
Morenoina |
On the branches of an unidentified palm |
Thailand |
| |
|
Botryosphaeriales | A |
Botryosphaeriaceae |
Barriopsis, Botryosphaeria, Diplodia[279] |
| , | Lasiodiplodia | , | Neodeightonia |
Durispora |
D. elaeidicola |
On dead rachides of Elaeis guineensis |
Malaysia |
N/A |
| Dothideomycetes (cont.) |
Incertae sedis (cont.) | [ | 118 | ] |
| Botryosphaeriales | (cont.) |
Phyllostictaceae |
Phyllosticta |
Dwibahubeeja |
D. indica |
On leaves of C. tenuis |
India (Uttar Pradesh) |
N/A |
[280] |
| |
|
Jahnulales |
Aliquandostipitaceae |
Jahnula |
Endosporoideus |
E. pedicellatus
(as E. pedicellata) |
On a dead petiole of Phoenix hanceana |
China (Hong Kong) |
| | N/A |
[ |
|
| 235] |
| Manglicolaceae |
Manglicola |
Fasciatispora |
F. nypae |
On a rotten frond of intertidal Nypa fruticans |
| |
|
Kirschsteiniotheliales | Brunei |
A |
[ |
Kirschsteiniotheliaceae161] |
| Kirschsteiniothelia |
Fissuroma |
F. maculans |
On dead leaves of Arenga westerhoutii |
Thailand |
A |
[281] |
| Flammispora |
F. bioteca |
On dead leaves of Licuala longicalycata submerged in peat swamp |
Thailand |
A |
[282] |
| Fluviatispora |
F. tunicata |
On submerged rachides of Livistona sp. |
Papua New Guinea |
N/A |
[174] |
| Frondicola |
F. tunitricuspis |
On decaying fronds of Nypa fruticans |
Brunei |
N/A |
[162] |
| Frondisphaeria |
F. palmicola |
On a rachis of Eugeissona minor |
Brunei |
N/A |
[170] |
| Frondispora |
F. bicalcarata |
On dead petioles of Chamaerops humilis |
Italy |
N/A |
[111] |
| Gossypinidium |
G. sporodochiale |
On a dead rachis of Praestoea montana |
Puerto Rico |
A |
[283] |
| Guestia |
G. gonetropospora |
On a dead rachis of Mauritia flexuosa |
Ecuador |
N/A |
[150] |
| Haploanthostomella |
H. elaeidis |
On dead leaves and rachis of Elaeis guineensis |
Thailand |
A |
[284] |
| Haplohelminthosporium |
H. calami |
On living leaves and petioles of Calamus sp. |
Thailand |
A |
[285] |
| Helensiella (as Digitella) |
H. rigidophora
(as D. rigidophora) |
On a rachis of an unidentified palm |
Mexico (Veracruz) |
N/A |
[286,287] |
| Helminthosporiella |
H. stilbacea |
On a dead petiole of Cocos nucifera |
Thailand |
A |
[285,288] |
| Hemisynnema # |
H. malayasianum |
On a rachis of an unidentified palm |
Malaysia |
N/A |
[289] |
| Hyalobelemnospora |
H. amazonica |
On a rotten petiole of an unidentified palm |
Peru |
N/A |
[64] |
| Kalamarospora |
K. multiflagellata |
On rachides of dead leaves of Sabal palmetto |
USA (Florida) |
N/A |
[290] |
| Letendraeopsis |
L. palmarum |
On leaves of Euterpe oleracea |
Brazil (Pará) |
N/A |
[291] |
| Lockerbia |
L. palmicola |
On dead rachides of an unidentified palm |
Australia (Queensland) |
N/A |
[114] |
| Longicorpus |
L. striatisporus
(as L. striataspora) |
On a decayed rachis of Nypa fruticans |
Thailand |
A |
[9] |
| Mackenziella (as Mackenziea) |
M. livistonae |
On decaying rachides of Oraniopsis appendiculata |
Australia (Queensland) |
N/A |
[15] |
Maculatifrondes
(as Maculatifrondis) |
M. aequatoriensis |
On leaves of an unidentified palm in rainforest |
Ecuador |
N/A |
[208] |
| Maculatipalma |
M. frondicola |
On a leaf of Linospadix microcarya |
Australia (Queensland) |
N/A |
[197] |
| Malthomyces |
M. calamigena
(as M. calamigenus) |
On tissues of Calamus rudentum |
Sri Lanka |
N/A |
[212] |
| Manokwaria |
M. notabilis |
On dead rachides of an unidentified palm in freshwater swamp |
Indonesia |
N/A |
[109] |
| Monosporoschisma |
M. elegans |
On a dead material of an unidentifed palm |
Chian (Hainan) |
A |
[268] |
| Neoastrosphaeriella |
N. krabiensis |
On a petiole of Metroxylon sagu |
Thailand |
A |
[281] |
| Neobarrmaelia |
N. hyphaenes |
On leaves of Hyphaene sp. |
South Africa |
A |
[292] |
| Neolinocarpon |
N. globosicarpum |
On decaying intertidal fronds of Nypa fruticans |
Brunei |
N/A |
[162] |
| Neoxylaria |
N. arengae |
On a dead petiole of Arenga pinnata |
Thailand |
A |
[293] |
Nigromammilla
(as Nigramammilla) |
N. calami |
On a sheath of dead rattan of Daemonorops margaritae |
China (Hong Kong) |
N/A |
[179] |
| Nipicola |
N. carbospora |
On immersed fronds of Nypa fruticans |
Brunei |
N/A |
[163] |
| Nusia |
N. scheeleae |
On a rachis of Scheelea insignis |
Singapore |
N/A |
[294] |
| Nypaella |
N. frondicola |
On intertidal fronds of Nypa fruticans |
Brunei |
N/A |
[164] |
| Ornatispora # |
O. palmicola |
On a dead rachis of an unidentified palm |
Ecuador |
N/A |
[181] |
| Oxodeora |
O. petrakii |
On living fronds of Oreodoxa regia |
Dominican Republic |
N/A |
[212] |
| Palmaria (as Palmomyces) |
P. montanea
(as P. montaneus) |
On a leaf of Oraniopsis appendiculata |
Australia (Queensland) |
N/A |
[143] |
| Palmeiromyces |
P. chamaeropicola |
On leaf spots of Chamaerops humilis |
Portugal |
A |
[295] |
| Palmicola |
P. archontophoenicis |
On a fallen rachis of Archontophoenix alexandrae |
Australia (Queensland) |
N/A |
[108] |
| Paracapsulospora |
P. metroxyli |
On a dead Metroxylon sagu |
Thailand |
A |
[296] |
| Paradactylella |
P. peruviana |
On a rotten petiole of an unidentified palm |
Peru |
N/A |
[64] |
| Paraproliferophorum |
P. hyphaenes |
On living leaves of Hyphaene sp. |
South Africa |
A |
[297] |
| Pararamichloridium |
P. livistonae |
On leaves of Livistona australis |
Australia (New South Wales) |
A |
[298] |
| Parateichospora |
P. phoenicicola |
On leaves of Phoenix reclinata |
South Africa |
A |
[299] |
| Phaeochoropsis |
P. neowashingtoniae |
On leaves of Neowashingtonia filamentosa |
USA (California) |
N/A |
[212] |
| Phaeomonilia |
P. pleiomorpha |
On a decaying petiole of an unidentified palm submerged in stream |
Mexico (Veracruz) |
N/A |
[300] |
| Phruensis |
P. brunneispora |
On a dead trunk of Licuala longicalycata |
Thailand |
A |
[301] |
| Polybulbophiale |
P. palmicola |
On the decaying petiole of Licuala sp. |
Brunei |
N/A |
[190] |
| Porodiplodia |
P. livistonae |
On leaves of Livistona australis |
Australia (New South Wales) |
A |
[302] |
| Pseudopalawania |
P. siamensis |
On a dead rachis of Caryota sp. |
Thailand |
A |
[303] |
| Pulmosphaeria |
P. archontophoenicis |
On a dead petiole of Archontophoenix alexandrae |
Australia (Queensland) |
N/A |
[194] |
| Quasiphoma |
Q. hyphaenes |
On leaves of Hyphaene sp. |
South Africa |
A |
[292] |
| Rachidicola |
R. palmae |
On a rachis of Calamus sp. |
China (Hong Kong) |
N/A |
[129] |
| Rattania |
R. setulifera |
On leaves of Calamus thwaitesii |
India (Goa) |
N/A |
[304] |
| Rogergoosiella |
R. roystoneicola |
On a dead petiole of Roystonea regia |
Cuba |
N/A |
[305] |
| Sabalicola |
S. sabalensioides |
On petioles of Sabal serrulata |
USA (Florida) |
N/A |
[122] |
| Sawantomyces |
S. indicus (as S. indica) |
On a spathe of Cocos nucifera |
India (Maharashtra) |
N/A |
[306] |
| Setophiale |
S. unisetulata |
On a decayed petiole of an unidentified palm |
Peru |
N/A |
[65] |
| Sorokinella |
S. appendicospora |
On a dead petiole of Livistona chinensis |
China (Hong Kong) |
N/A |
[6] |
| Stratiphoromyces |
S. brunneisporus |
On decaying petioles of Licuala sp. |
Brunei |
N/A |
[189] |
| Striatiguttula |
S. nypae |
On a decayed rachis of Nypa fruticans |
Thailand |
A |
[9] |
| Thailandiomyces |
T. bisetulosus |
On senescent trunks of Licuala longicalycata |
Thailand |
A |
[307] |
| Tirisporella |
T. beccariana |
On decaying leaf bases of Nypa fruticans |
Malaysia |
N/A |
[167] |
| Tretendophragmia |
T. palmivora |
On a rachis of Korthalsia sp. |
Singapore |
N/A |
[308] |
| Tretocephala |
T. decidua |
On a leaf sheath and rachis of Oncosperma horridum |
Singapore |
N/A |
[309] |
| Tribulatia |
T. appendicospora |
On a dead petiole of Archontophoenix alexandrae |
Australia (Queensland) |
N/A |
[8] |
| Triseptatospora |
T. calami |
On dead petioles of Calamus sp. |
Thailand |
A |
[310] |
| Unisetosphaeria |
U. penguinoides |
On a petiole of Eleiodoxa conferta submerged in peat swamp |
Thailand |
N/A |
[245] |
| Uwemyces |
U. elaeidis |
On leaves of Elaeis oleifera |
Colombia |
A |
[288] |
| Venustocephala |
V. aequatorialis |
On a decayed petiole of an unidentified palm |
Ecuador |
N/A |
[65] |
Venustisporium
(as Venustusporium) |
V. chelyoforme
(as V. chelysforme) |
On fallen rotten leaves of Bactris setulosa |
Venezuela |
N/A |
[311] |
| Veramycella |
V. bispora |
On rachides of dead leaves of Sabal palmetto |
USA (Florida) |
N/A |
[312] |
| Veramyces |
V. manuensis |
On a rotten petiole of an unidentified palm |
Peru |
N/A |
[64] |
| Waihonghopes |
W. australiensis |
On a decaying rachis of Oraniopsis appendiculata |
Australia (Queensland) |
N/A |
[15] |
There are several scattered reports of palmicolous hyphomycetes in different regions of the world. For instance, species of helicosporous fungi from various genera, such as
Drepanospora,
Helicoma,
Helicomyces,
Helicosporium, and
Xenosporium, have been recorded in collections of palm tissues. These were reviewed by Goos [
318,
319,
320,
321,
322,
323,
324,
325] and Goos et al. [
326] during their studies on anamorphic genera of helicosporous fungi. In the 1990s, along with the examples previously cited by Hyde and co-workers, several palmicolous hyphomycetes were reported in studies carried out by Subramanian in India and in Southeast Asian countries, such as Malaysia and Singapore (
Figure 1). In these studies, Subramanian [
259,
262,
266,
289,
294,
308,
309,
327,
328] introduced many new species and genera of dematiaceous hyphomycetes from different palm trees species, including
Tretendophragmia [
308],
Tretocephala [
309],
Ashtaangam,
Basauxia [
266],
Acarocybellina [
259],
Agrabeeja [
262],
Nusia [
294], and
Hemisynnema [
289] (
Table 1). These records further emphasised the importance of palm trees for the description of taxonomic novelties and for the study of fungal biodiversity, and the same trend would be found in the palm trees of Central American countries.
Palmicolous hyphomycetes have been widely collected from palms in Central American countries, including Cuba and Mexico (
Figure 1), by Castañeda-Ruiz, Holubová-Jechová, Mena-Portales, Mercado-Sierra, and many other co-workers, following their previously mentioned investigation of hyphomycetes from Cuba in the 1980s. Although most of these studies were not conducted to explicitly evaluate palm fungi, the extent of these reports has made it possible to reveal the composition of the assemblage of palm hyphomycetes that inhabit palm tissues in the tropics. Furthermore, it has become evident, particularly in studies from Cuba, that some parts of palm trees, mainly their decaying rachides and large petioles, are exceptional substrata for the growth and development of microfungi, mainly hyphomycetes, and several new taxa have been identified [
260,
305,
329,
330,
331,
332,
333,
334,
335,
336,
337,
338,
339,
340,
341,
342,
343,
344].
Similar to what Hyde and co-workers discovered in their investigation in East and Southeast Asia, Australasia, and Ecuador, many of the fungi growing on palm trees in Cuba were described as new to science and found to form an autochthonous mycobiota, where host-specificity, sometimes at the host genus or species level, is often observed. The fungus–host plant relationship and the factors that affect it were expressly discussed and reviewed by Mercado-Sierra et al. [
329], particularly for genera of palm trees that grow abundantly in Cuba, namely
Roystonea,
Cocos, and
Coccothrinax. The Cuban royal palm
Roystonea regia appears to be a particular case where an enormous diversity of fungi was found, including 265 species from different taxonomic groups, a number much higher than that reported for other plant species endemic to Cuba [
345]. Moreover, its relevance and importance in studies of hyphomycetes were pointed out by Mercado-Sierra [
73]. Many of the fungi identified on palm trees from Cuba were new reports for Cuban mycobiota and several taxa were described as new to science, including more than thirty species and six genera, viz.
Consetiella [
75],
Holubovaea [
73],
Phragmospathulella [
96],
Cheiromyceopsis [
99],
Rogergoosiella [
305], and
Acarocybiopsis [
260] (
Table 1).
Several Cuban provinces and localities, such as Pinar Del Río, Camagüey, Matazanas, and Sancti Spíritus, have been extensively sampled during surveys of hyphomycetes in protected natural areas in Cuba, and on several occasions these hyphomycetes have been collected from palm trees, e.g., refs. [
338,
346,
347,
348,
349,
350,
351]. Although the extension of these studies made it possible to uncover the assemblage of palm hyphomycetes in those locations, their objective was mainly to study ecological stations, biosphere reserves, and protected areas in order to promote the conservation of important hotspots of fungal diversity. These studies continue to be carried out today and report an exceptional diversity of palm fungi. Recently, Mena-Portales et al. [
352], following Mercado-Sierra et al. [
329], reviewed and analysed the relationship between fungal diversity and palms trees in Cuba by compiling information on some interesting species of hyphomycetes found in different
Arecaceae hosts.
Similar studies have also been carried out in other Central American countries (
Figure 1). Mercado-Sierra et al. [
353,
354] reported some palmicolous hyphomycetes from Costa Rica. Recently, a checklist of asexual fungi from Costa Rica, which compiled information obtained during 1927 to 2018 based on scientific papers, was presented by Granados-Montero et al. [
355], including several reports of hyphomycetes and coelomycetes on palm trees. Very few palm fungi, including hyphomycetes, have been reported from Puerto Rico and most reports have been summarised in an annotated bibliography entitled “The Fungi of Puerto Rico and the American Virgin Islands” by Stevenson [
356]. However, some new species were later introduced based on occasional palm collections, e.g., refs. [
240,
357,
358]. Palmicolous hyphomycetes from Panama and Nicaragua have been listed in checklists of Panamanian and Nicaraguan fungi by Piepenbring [
359] and Delgado-Rodríguez [
360], respectively. Likewise, several palmicolous hyphomycetes were reported in Mexico during studies carried out in order to increase the knowledge about Mexican hyphomycetes [
286,
300,
361,
362,
363,
364,
365,
366,
367,
368,
369,
370,
371], where two new genera,
Phaeomonilia [
300] and
Digitella [
286], and some new species were introduced (
Table 1). Similar to the studies in Cuba, the investigation in Mexico, which continues today, was not pursued to study palm fungi, but to inventory and gain knowledge about conidial fungi from plant litter in tropical forests, particularly in the states of Campeche, Tabasco, and Veracruz, due to the accelerated deforestation of tropical habitats, e.g., refs. [
367,
371,
372,
373,
374,
375,
376,
377,
378,
379,
380]. As a result, since palm trees are an important component of the flora of these forests, several palmicolous hyphomycetes have been reported.
In addition to the studies in Central American countries, South American countries have also been occasionally surveyed (
Figure 1). Therefore, palmicolous hyphomycetes have been reported in collections from Argentina [
381], Colombia [
382], Peru [
265], and Venezuela [
272,
311,
383,
384,
385,
386,
387], where some new species and genera have been introduced (
Table 1).
2.1.4. Palm Fungi from Understudied Tropical Hotspots, Argentina, India, and Brazil
Some scattered studies have surveyed palmicolous fungi in Argentina. However, with the exception of Mercado-Sierra et al. [
381], these were not systematic studies of descriptive taxonomy, but studies carried out to better understand the diversity of ascomycetes in woody parts of palms in Argentina, especially in areas or parks that had been proposed as natural reserves for protection and where some native palm species are an important element of the local flora, including
Butia yatay,
Euterpe edulis, and
Syagrus romanzoffiana [
388,
389,
390]. Capdet and Romero [
389] summarised previous information on palm fungi and their occurrence in Argentina, reflecting on the lack of knowledge about palm fungi in the country.
Similarly, considering the available literature on palm fungi, except for a few stray collections, no comprehensive investigation on the fungal diversity that occurs on palms in India has been carried out (Figure 1). However, some remarkable reports by Subramanian and his students from Chennai, P. R. Rao, D. Rao, and V. Rao, and colleagues from Hyderabad and Bhat and colleagues from Goa, revealed a considerable diversity of palmicolous fungi, especially hyphomycetes, occurring on leaf litter. An overview of these studies is given here.
Subramanian [
391,
392,
393,
394,
395,
396,
397], in his series of seven papers “Fungi imperfecti from Madras”, reported several new palmicolous hyphomycetes from dead palm leaves in Chennai. Moreover, in addition to his previously mentioned reports on dematiaceous hyphomycetes [
259,
262,
266,
289,
294,
308,
309,
327,
328], Subramanian extensively studied hyphomycetes in tropical regions, especially India. These studies resulted in a major monograph of the Indian species [
398], many of which were collected from palm material and introduced as new to science [
399,
400,
401,
402,
403,
404,
405,
406]. A number of these new palmicolous hyphomycetes were introduced in his series of six papers entitled “Hyphomycetes”, where new genera and species were described on the leaves of several different palm species, such as
Cocos nucifera,
Phoenix canariensis,
Rhopalostylis sapida, and
Borassus flabellifer from Tamil Nadu [
407,
408,
409,
410,
411,
412].
Rao and Rao [
413,
414,
415,
416,
417,
418,
419,
420], Chaudhury and Rao [
421], and Rao and Chaudhury [
422], also reported several palmicolous fungi, including new taxa, from different palm species, such as
B. flabellifer,
Caryota urens,
C. nucifera, and
Livistona chinensis, mainly from Hyderabad. Moreover, new species of palmicolous fungi were reported by Rao [
423,
424,
425] from Maharashtra. Later, Varghese and Rao [
426,
427] recorded several palm fungi during their mycological survey of the forests of Kerala, near where Pande and Rao [
428] collected the new species
Rosellinia lakshadweepensis on the pericarp of
C. nucifera from the island of Kavaratti (Lakshadweep).
Bhat and co-workers isolated some new taxa of palmicolous litter-inhabiting hyphomycetes in palm collections from India in their studies on fungi from the forests of the Western Ghats hills in Goa, the rainforests of the Andaman-Nicobar Islands and, to a lesser extent, the humid mountains forests of Northeastern Himalayas, e.g., refs. [
429,
430,
431,
432]. In addition to these taxonomic studies, ecological studies were also carried out on litter colonisers and endophytes in plant species from the forests of the Western Ghats in Goa, which included palm trees species, such as
Calamus thwaitesii.
C. urens, and
Elaeis guineensis [
433,
434].
A vast area of the peninsular India still remains underexplored for fungal biodiversity [
435]. However, the fungi of the forests of the Western Ghats in Goa and the rainforests of the Andaman-Nicobar Islands, two of the biological hotspots of India, have been explored to some extent, e.g., refs. [
436,
437,
438,
439,
440,
441]. In turn, many palm fungi have been documented, including several new taxa (
Figure 1,
Table 1). Most of these studies were reviewed by Bhat [
437], who considered the forests of the Western Ghats as “an abode of novel and interesting microfungi” and presented a list of new hyphomycetes discovered in them, where several palmicolous fungi are cited. Further studies on the diversity of microfungi from these forests in Goa and in some parts of Karnataka, Kerala, Tamil Nadu, and Maharashtra were carried out by Pratibha et al. [
442,
443], which resulted in the documentation of a few more palmicolous hyphomycetes. Still, today studies on the fungi of the forests of the Western Ghats hills and the rainforests of the Andaman-Nicobar Islands continue to be carried out and to report an exceptional diversity of palm fungi. For instance, Dubey and Moonnambeth [
306,
444,
445,
446,
447,
448], Dubey [
449], and Dubey and Neelima [
450] documented several dematiaceous palmicolous hyphomycetes, including new taxa, during an investigation of fungi from the forests of the Western Ghats of Maharashtra (
Table 1). More recently, Niranjan and Sarma [
451] compiled a checklist of fungi reported from the Andaman-Nicobar Islands, where many palmicolous fungi have been documented. Several other palmicolous fungi, including new species, have been reported from the rainforests of these islands by Ram and Sinha [
452] and Niranjan and Sarma [
453,
454,
455,
456,
457].
Although information on Indian palm fungi is scarce and difficult to review and compile properly, fungi collected from living palm leaves, diseased palm foliage, and palm litter by various researchers have been included in periodically published lists and compilations of Indian fungi, for example “List of Indian fungi 1952–1956” [
404], “List of Indian fungi 1956–1960” [
458], “Fungi of India 1989–2001” [
459], “Ascomycetes of Peninsular India” [
460], and, more recently, “Bilgrami’s Fungi of India List and References (1988–2020)” [
461]. One of these compilations was dedicated exclusively to palm fungi under the title “Fungal records on palms from India” by Pande et al. [
462], who made a list of fungi recorded on palm trees up to 1999 and listed 355 species distributed in 188 genera described from 29 species of palm trees from different Indian states.
Brazil is a country rich in palm species, some of which have great socio-economic value [
463]. Even so, few taxonomic studies have been carried out on Brazilian palm fungi (
Figure 1) and most of them have focused on palm hyphomycetes. However, the few studies available have also revealed the trend towards the presence of a rich fungal diversity, from which many new palmicolous fungi have been introduced. A summary of some examples of these studies is given herein. In 1978, Hennen and Ono [
464] identified the first rust fungus on a palm tree, the new genus and species
Cerradoa palmaea on
Attalea ceraensis from Brasília. Several palm fungi were gathered from collections of
Astrocaryum from Amazonas and published by Farr [
465,
466,
467] in his series of papers “Amazonian foliicolous fungi”. Later, Rodrigues and her colleagues recorded some new palmicolous taxa when studying the endophytic fungi that inhabit the tissues of the Amazonian palm
Euterpe oleracea [
291,
468,
469,
470,
471,
472,
473] (
Table 1). Rodrigues [
468,
472] published the first study on the fungal endophytes inhabiting the foliage of
E. oleracea from Combu Island, growing in the Brazilian Amazon estuary and in Amazonian floodplains, and listed 57 species of palm ascomycetes, including several hyphomycetes. In addition, several fungi recorded on arecaceous hosts from Brazil have been compiled by Silva and Minter [
474] and Mendes et al. [
475,
476].
Many other sparse reports of palm fungi from Brazilian regions have been made. However, similar to the studies on palm fungi from Argentina, most of the studies on Brazilian palm fungi were not carried out as systematic studies of descriptive taxonomy. Instead, these studies aimed to better understand the diversity of conidial fungi, especially hyphomycetes, on woody palms of the Amazon rainforest and other biomes, where biodiversity research and conservation programs were being conducted. An overview of these studies is presented here.
The investigation on conidial fungi associated with the decomposition of palm leaves in the Amazon rainforest has been restricted to three areas, namely the National Forest of Caxiuanã [
274,
387,
477,
478,
479,
480,
481] and the Combu Island [
482,
483] in Pará, and the National Forest of Amapá in Amapá [
264,
387,
479,
481]. Several
Arecaceae hosts have been sampled, including members of
Astrocaryum,
Attalea,
Bactris,
Euterpe,
Geonoma,
Maximiliana,
Oenocarpus,
Socratea, and
Syagrus. Most of these studies were motivated both by the accelerated deforestation of the Amazon rainforest and by the lack of research into the diversity of palm fungi in the tropical regions of South America. In addition to the importance of these studies for expanding the knowledge about the composition and distribution of palm fungi in the Amazon rainforest, several new records for these Brazilian regions, as well as new genera and species, have been reported, e.g., refs. [
264,
274,
387,
479] (
Table 1).
Surveys of microfungi on palm trees have also been carried out in Bahia, Brasília, and Pernambuco, particularly in biomes in areas of Northeast Brazil where palm trees are a major component [
484,
485,
486,
487,
488,
489,
490,
491,
492,
493,
494,
495,
496,
497,
498,
499,
500,
501,
502,
503,
504,
505,
506,
507,
508,
509,
510,
511,
512,
513,
514,
515,
516]. These studies aimed to understand the fungal composition of important biomes of Northeast Brazil, such as the Atlantic rainforest, e.g., refs. [
496,
499,
500], the Caatinga, e.g., refs. [
497,
501,
502,
503,
504,
507,
508,
509,
512], and the Cerrado, e.g., refs. [
485,
486,
489], as well as to uncover the mycota that inhabit the tissues of important palms that can be found there, including
Acrocomia intumescens,
Attalea funifera,
Bactris acanthocarpa,
Cocos nucifera,
Elaeis guineensis,
Euterpe edulis,
E. oleracea,
Mauritia flexuosa,
Polyandrococos caudescens,
Syagrus botryophora, and
S. coronata. In turn, several new palmicolous taxa have been reported, including members of the typical palmicolous mycota found by Hyde and co-workers on their extensive investigation on East and Southeast Asian and Australasian countries. For instance, Vitória et al. [
491,
495] introduced the new species
Arecomyces attaleae and
Neolinocarpon attaleae on dead rachides of
A. funifera from Bahia.
2.1.5. Palm Fungi and Reflections on the Recent Input from Molecular Era
This comprehensive overview of the literature shows that palms support a vast array of fungi, especially ascomycetes. In the well-studied tropical regions of East and Southeast Asia, Australasia, and, to a lesser extent, Central America (
Figure 1), where these fungi have received considerable attention, a remarkable diversity of fungi has been revealed with the description of numerous new taxa, e.g., refs. [
6,
8,
104,
329,
352]. Much of this diversity can be attributed to the tropical and subtropical habitats surveyed, where the diversity of fungi is known to be higher [
10]. However, the few studies carried out on palms that thrive in temperate regions have also revealed a considerably rich fungal diversity, of which some taxa have been described as new to science. For instance, several palmicolous fungi have been recorded in New Zealand (
Figure 1), particularly from collections of
Rhopalostylis spp. [
54,
517,
518,
519,
520,
521]. McKenzie et al. [
54] noted that 147 named fungal species and 50 fungal records identified only to genus have been recorded on
Rhopalostylis, mainly on
R. sapida from New Zealand. Thus, the great diversity of palmicolous fungi recovered can be also attributed to the wide variety of palm hosts and habitats studied, including many different palm species and tissues in terrestrial, freshwater, and marine or mangrove ecosystems. While the first studies by Hyde and co-workers covered the diversity of fungi on mangrove palms, e.g., ref. [
168], most subsequent studies were dedicated to surveying saprobic fungi that develop on palm substrata from tropical rainforests, e.g., ref. [
150]. In addition, few studies were conducted to survey palm phytopathogens, e.g., ref. [
212].
Up to 2003, the intensive research carried out by Hyde and co-workers has reported more than 320 new species and more than 45 new genera of palmicolous fungi. However, in all these earlier studies, all the taxa reported were introduced, described, and arranged in different taxonomic ranks within the
Ascomycota based solely on their morphology. This is currently known as a subjective approach and many taxa have been assigned to
Ascomycota genera
incertae sedis. Given that palm trees are important hosts that harbour potential novel taxa, it is critical that these palmicolous fungi are recollected, epitypified where needed, and isolated so that molecular data can be obtained and used to establish their natural phylogenetic placements [
522]. For example, regarding the initial examples of some of the most common fungal genera found on palms, such as
Anthostomella,
Astrosphaeriella,
Linocarpon, and
Oxydothis, several species have been recorded on palms, e.g., refs. [
111,
134,
138], but only a small percentage of these have associated sequence data available. As a result, their position in a natural taxonomic framework is poorly supported and, consequently, their evolutionary relationships in higher taxonomic ranks are poorly understood.
Some of the first studies on palmicolous fungi that combined molecular data with morphological data were those by Pinruan et al. [
270,
282,
301,
307,
523] and Pinnoi et al. [
524,
525] in Thailand, who reported several new taxa of saprobic fungi on
Calamus species and on the peat swamp palms
Licuala longecalycata and
Eleiodoxa conferta. These studies were a continuation of previous solely morphological studies that had already yielded a remarkably rich fungal diversity from palms in the Sirindhom Peat Swamp Forest at Narathiwat, Southern Thailand [
236,
241,
242,
243,
244,
245,
246,
247]. Four new genera have been introduced (
Table 1) and their phylogenetic relationships among extant taxa have begun to be unveiled. An interesting example worth mentioning is the description of the new species
Astrocystis eleiodoxae on petioles of
E. conferta submerged in a peat swamp from Thailand by Pinnoi et al. [
525]. Much of the early studies by Hyde and co-workers focused on the treatment of several genera that occur on palms and their taxonomic relationships and placement based mainly on morphological characters related to asci, ascospores, and associated features. The taxonomic placement of the genus
Astrocystis, as well as several other xylariaceous-related genera commonly recorded on palms, has been analysed, although it has often remained obscure due to the assessment of its morphology alone, e.g., ref. [
150]. The description of
A. eleiodoxae by morphological and phylogenetic means, as well as other new xylariaceous taxa from palms, such as
Rosellinia capetribulensis on decaying rachides of
Calamus sp. from Northern Queensland, Australia by Bahl et al. [
526], underlines the importance of introducing molecular data into the study of palm fungi as a highly diverse fungal group. Although the assignment of these taxa to higher taxonomic ranks remained ambiguous and undetermined, the relationship with other xylariaceous genera started to become clearer than when these studies were conducted solely on the basis of morphology.
The introduction of DNA sequence data to study fungal biodiversity in the early 1990s has served as a stimulus for the description of new taxa from palms. Hence, several of the old collections have been accessed and their taxonomic placement clarified. Over the last 15 years, Hyde and co-workers have been revisiting their studies on palms, recollecting and epitypifying some of the taxa and clarifying their phylogenetic position among extant and new taxa [
522]. This, in turn, has refined the identity of the most common mycota that inhabits palm tissues by providing information on the natural grouping of palmicolous genera based on sequence analyses (
Table 2). The number of reports has been increasing rapidly, especially in collections from Northern Thailand (
Figure 1), revealing several new palmicolous taxa that have complexified the taxonomic structure of palm fungi (
Table 1 and
Table 2).
Several new species of the common palm mycota were introduced, along with new genera, including members of the
Sordariomycetes and
Dothideomycetes frequently found on palms. This ultimately led to the establishment of new families to accommodate and clarify their phylogenetic relationships. Indeed, new taxa of some of the most common fungal genera found on palms, including
Oxydothis [
527,
528,
529],
Linocarpon,
Neolinocarpon [
530],
Astrosphaeriella sensu lato [
267,
281,
531,
532,
533], and
Roussoella [
534], are continuously reported on palm tissues and the new families
Oxydothidaceae [
528],
Linocarpaceae [
530],
Astrosphaeriellaceae [
267], and
Roussoellaceae [
534], respectively, have been introduced to accommodate them (
Table 2). A historical account of molecular studies carried out on palm fungi is presented herein. In addition, some case studies of common palm taxa are highlighted to reflect on the impact of molecular data on the taxonomy and biodiversity of this group of fungi.
Astrosphaeriella-like Taxa: A Polyphyletic Nature Hiding Cryptic Genera
Astrosphaeriella is an interesting case study of how phylogenetics and its inputs had implications in the taxonomic structure of palm fungi. Although there have been many morphological-based studies of
Astrosphaeriella, including those major morphological studies by Hyde and Fröhlich [
144] and Hyde et al. [
148], based particularly on species of
Astrosphaeriella occurring on palms in tropical regions, no thorough molecular investigation of the genus has been carried out. The taxonomy of
Astrosphaeriella and its natural placement, especially at family level, stayed unresolved until recently. The frequent collection of
Astrosphaeriella-like species on palms and other monocotyledonous trees has made it possible to move towards their natural classification. Thus, in addition to understanding the polyphyletic nature of
Astrosphaeriella sensu lato, Liu et al. [
281] erected two new genera of
Astrosphaeriella-like species in
Aigialaceae, viz.
Fissuroma and
Neoastrosphaeriella (
Table 1). Later, Phookamsak et al. [
267] recognised that
Astrosphaeriella-like species can be distinguished into three families and established
Astrosphaeriellaceae for typical
Astrosphaeriella species (
sensu stricto),
Pseudoastrosphaeriellaceae to accommodate the new genus
Pseudoastrosphaeriella, and the new genus
Astrosphaeriellopsis for a distinct
Astrosphaeriella-like lineage basal to
Aigialaceae (
Table 1).
Astrosphaeriellopsis was later accommodated in
Astrosphaeriellaceae by Wanasinghe et al. [
532] following a multigene phylogeny and respective taxonomic circumscription of
Astrosphaeriella species and allied genera with the introduction of several novel palmicolous taxa in the genera
Astrosphaeriellopsis,
Fissuroma,
Neoastrosphaeriella, and
Pithomyces isolated from
Calamus,
Caryota, and
Licuala species in Northern Thailand and Southwest China. Further additions to
Fissuroma and
Neoastrosphaeriella were made by Konta et al. [
533] and Zhang et al. [
531] from palms in mangrove and terrestrial habitats in Thailand. Another genus,
Xenoastrosphaeriella (
Table 1), was introduced in
Astrosphaeriellaceae to accommodate saprobic fungi on bamboo and palms that previously represented a basal lineage in
Astrosphaeriellaceae [
535]. More recently, the new genus
Triseptatospora was introduced in
Astrophaeriellaceae to accommodate
T. calami found on dead petioles of
Calamus sp. in Thailand [
310] (
Table 1).
Several new
Dothideomycetes, along with
Astrosphaeriella sensu lato and
Roussoellaceae, are also being recorded from palm tree collections. Mapook et al. [
536] introduced the new family
Palawaniaceae to accommodate the
Palawania species, which are saprobes common on palms, occurring on dried fronds and spines. Later, Jayasiri et al. [
535] introduced two new species of palmicolous
Dothideomycetes in
Delitschia and
Vaginatispora on fallen fruit pericarp of
Nypa fruticans from Thailand. The new genus
Pseudopalawania was introduced in
Muyocopronaceae by Mapook et al. [
303] to accommodate
P. siamensis on a dead rachis of
Caryota sp. from Thailand (
Table 1). Recently, Yu et al. [
537] made new additions to
Occultibambusaceae based on collections of decaying petioles of
Trachycarpus fortunei from China. Several aquatic
Dothideomycetes, as well as
Sordariomycetes, have also recently been described from palm trees and will be discussed later.
Xylarialean and Related Sordariomycetes: The Enigmatic Anthostomella and Allied Genera
Regarding
Sordariomycetes, along with
Oxydothidaceae and
Linocarpaceae, many other new ascomycetes are being introduced and their phylogeny resolved based on collections from palms. However, several palmicolous genera are poorly represented with sequence data. In addition, several sequences are of poor quality, which can compromise the resolution of the phylogenetic placement of taxa, especially at higher taxonomic levels, such as family and class [
538]. This has often been observed in studies on palm fungi. For example, while introducing
Linocarpaceae to accommodate
Linocarpon and
Neolinocarpon, Konta et al. [
530] also introduced the new family
Leptosporellaceae in
Chaetosphaeriales to accommodate
Leptosporella, including two new species collected from palms, and provided a comparative morphological list of species in
Leptosporella,
Linocarpon, and
Neolinocarpon. However, as the authors stated, fresh collections of several of the known taxa with associated molecular data are needed to establish and strengthen their natural phylogenetic placements, since all the diagnosed clades are sparsely populated. Similarly, a checklist of
Diatrypaceae that occur on palms was presented by Konta et al. [
263], with the introduction of many new species and a new genus,
Allodiatrype (
Table 1). However, the generic taxonomic resolution of several lineages remained unclear, probably due to a lack of sequence data or previous misidentifications, so their phylogenetic placement will only be clear with fresh collections and adequate sequence data [
538].
The need to recollect and isolate some of the old collections of palm fungi was also reinforced by Daranagama et al. [
539], who re-examined the type specimens of some
Sordariomycetes genera to determine their family placement according to modern taxonomic concepts. As previously mentioned, the taxonomic placement of several xylariaceous genera, including those commonly recorded on palms, is often obscure due to the assessment of their morphology alone and the lack of sequence data to properly place them in a natural taxonomic framework. Moreover, novel taxa are often introduced when new collections and their respective molecular data are obtained. For instance, Konta et al. [
293] introduced the new genus
Neoxylaria (
Table 1) to accommodate the new species
N. arengae, as well as the morphological species
Xylaria juruensis (as
N. juruensis) from palm material in Brazil and
X. queenslandica (as
N. queenslandica) from
Archontophoenix alexandrae in Australia, using both their fresh collection of
N. arengae and morphology data, as there is no molecular data for these old collections. More recently, two new families were introduced for genera that frequently occur on palms, along with the introduction of new species.
Appendicosporaceae was introduced to accommodate
Appendicospora, with the analysis of a new fresh collection designed as a reference specimen for
A. hongkongensis on dead fronds of
Livistona chinensis from China [
540].
Fasciatisporaceae was introduced to accommodate
Fasciatispora, with the introduction of the new species
F. cocoes on decaying rachides of
Cocos nucifera from Thailand [
541].
Several species of
Fasciatispora have been introduced from palm trees since the first studies by Hyde et al. in the 1990s, and a synopsis of
Fasciatispora species, along with
Anthostomella, from decaying palm fronds collected in Indonesia and Thailand was presented by Hidayat et al. [
542]. However, only a few of them have molecular data available for inclusion in modern taxonomic concepts and most of the new species from the 1990s remain described based only on their morphology. Recently, the type species of the genus,
F. nypae, was re-collected on a frond of
Nypa fruticans from Thailand and a reference specimen was designated, which made it possible to begin to resolve the phylogeny of
Fasciatispora as a basal clade in
Xylariaceae [
543], which was later introduced as the family
Fasciatisporaceae [
541].
Likewise, several
Anthostomella species have been introduced from palm trees and, since its first synopsis by Hyde [
133], who accepted twenty-seven species (of which nine were new) occurring on palms, several other palmicolous
Anthostomella species have been described, e.g., refs. [
143,
209,
218,
219,
220,
221]. However, the phylogeny of this species-rich genus is still ambiguous. Although its polyphyletic nature has already been recognised, a significant phylogenetic and morphological re-assessment of
Anthostomella is still needed, including fresh collections and associated molecular data [
539,
544,
545]. None of the
Anthostomella species described from palms have been re-evaluated to gain knowledge about their phylogenetics, so it can be assumed that several new taxa have yet to be described under morphological
Anthostomella-like species. In fact, several
Anthostomella-like species have been analysed and placed in existing and new genera to properly accommodate them, e.g., refs. [
539,
541,
544,
545]. For instance, recently, Konta et al. [
284] introduced the new genus
Haploanthostomella from palms based on
H. elaeidis on dead leaves and rachides of
Elaeis guineensis from Thailand (
Table 1) and provided a key to genera with
Anthostomella-like characteristics. Konta et al. [
284] also provided a family replacement of
Endocalyx to
Cainiaceae.
Endocalyx is a genus of coelomycetes almost exclusively reported on palm hosts [
546,
547]. Although Delgado-Rodríguez et al. [
546] made a recent phylogenetic assessment of
Endocalyx, some species still lack molecular data, as is often the case with xylariaceous genera.
Palmicolous “Anamorphs”: A Plethora of Botryosphaeriaceae and Other Dothideomycetes
Following the trends of the late 1990s, along with the description of the
Ascomycota coverage through the presence of its sexual morphs on the host, reports on palmicolous “anamorphs” began to become more frequent, e.g., refs. [
241,
243,
244,
245,
246]. In recent years, several species of
Botryosphaeriaceae are being introduced as new to science based on palm collections from different regions of the world, particularly species of
Neodeightonia [
505,
547,
548,
549,
550,
551,
552,
553]. Moreover, some new pestalotioid-like species have recently been described on palms, including species of
Neopestalotiopsis,
Pestalotiopsis,
Pseudopestalotiopsis [
296,
310,
554,
555,
556,
557,
558,
559],
Seridium [
560], and
Morinia [
561].
Dematiaceous hyphomycetes, as demonstrated by studies on palmicolous hyphomycetes from Central American countries and India, are also frequently reported on palm tissues and their phylogenetics is continuously revealing new taxa. Li et al. [
562] introduced the new family
Zygosporiaceae to accommodate
Zygosporium, a widespread genus usually associated with monocotyledonous, including palms. Delgado-Rodríguez, in his series of papers on “South Florida microfungi”, in which many new species of palm hyphomycetes were described, e.g., refs. [
290,
312,
563,
564,
565,
566], introduced a new species of
Taeniolella on the petiole of a dead leaf of
Sabal palmetto, expanding the concept of the strong polyphyly of the genus among different classes [
567]. Later, Delgado-Rodríguez and co-workers introduced the new genus
Castanedospora to accommodate
Sporidesmium pachyanthicola based on an epitype specimen collected on the petiole of a dead leaf of
S. palmetto, redefining its placement at family level in the
Extremaceae [
273] (
Table 1). Moreover, the phylogenetic placement of
Ernakulamia cochinensis, one of the saprobic hyphomycetes taxa commonly found associated with palm hosts, as a member of
Tetraplosphaeriaceae in
Pleosporales, was investigated based on a representative specimen collected on
Astrocaryum standleyanum in Panama [
568]. Species of
Hermatomyces are being described on palms from different regions of the world, including Panama [
569], Thailand [
570
| |
| |
| |
| Incertae sedis |
| Taeniolella |
| |
| |
| Muyocopronales |
| Muyocopronaceae |
| Muyocopron |
| , |
| Pseudopalawania |
| |
| |
| Tubeufiales |
| Tubeufiaceae |
| Aquaphila |
| , |
| Berkleasmium |
| , |
| Helicoma |
| , |
| Helicomyces |
| , |
| Helicosporium |
| , |
| Thaxteriella |
| |
| |
| |
Wiesneriomycetaceae |
Wiesneriomyces |
| |
|
- |
Palawaniaceae |
Palawania |
| |
|
- |
Trichopeltinaceae |
Acrogenotheca |
| |
|
- |
- |
Letendraeopsis, Xenosporium, Brooksia, Dianesea, Leptomeliola, Scolionema |
| Sordariomycetes |
Diaporthomycetidae |
Annulatascales |
Annulatascaceae |
Annulatascus, Submersisphaeria |
| |
|
Diaporthales |
Diaporthaceae |
Diaporthe |
| |
|
|
Gnomoniaceae |
Maculatipalma |
| |
|
|
Melanconidaceae |
Melanconis, Melanconium |
| |
|
|
Schizoparmaceae |
Coniella |
| |
|
|
Incertae sedis |
Durispora, Phruensis |
| |
|
Distoseptisporales |
Distoseptisporaceae |
Distoseptispora |
| |
|
Magnaporthales |
Magnaporthaceae |
Gaeumannomyces |
| |
|
|
Ophioceraceae |
Ophioceras |
| |
|
|
Pseudohalonectriaceae |
Pseudohalonectria |
| |
|
Ophiostomatales |
Ophiostomataceae |
Hyalobelemnospora |
| |
|
Phomatosporales |
Phomatosporaceae |
Phomatospora |
| |
|
Tirisporellales |
Tirisporellaceae |
Bacusphaeria, Thailandiomyces, Tirisporella |
| Sordariomycetes (cont.) |
Diaporthomycetidae (cont.) |
Xenospadicoidales |
Xenospadicoidaceae |
Koorchaloma, Spadicoides |
| |
|
Incertae sedis |
Mesnieraceae |
Bondiella |
| |
|
|
Trichosphaeriaceae |
Unisetosphaeria |
| |
|
|
Thyridiaceae |
Thyridium |
| |
|
|
- |
Paraproliferophorum |
| |
Hypocreomycetidae |
Glomerellales |
Plectosphaerellaceae |
Acremoniisimulans, Brunneomyces |
| |
|
Hypocreales |
Bionectriaceae |
Acremonium, Clonostachys, Gossypinidium, Hydropisphaera, Ijuhya, Lasionectria, Nectriella, Paracylindrocarpon |
| |
|
|
Hypocreaceae |
Verticimonosporium |
| |
|
|
Nectriaceae |
Baipadisphaeria, Calonectria, Chaetopsina, Cosmospora, Dactylonectria, Fusarium, Ilyonectria, Nectria, Nectriopsis, Ophionectria, Pleiocarpon, Volutella |
| |
|
|
Neoacremoniaceae |
Neoacremonium |
| |
|
|
Niessliaceae |
Niesslia |
| |
|
|
Stachybotryaceae |
Alfaria, Stachybotrys, Virgatospora |
| |
|
Microascales |
Gondwanamycetaceae |
Custingophora |
| |
|
|
Halosphaeriaceae |
Aniptodera, Cirrenalia, Fluviatispora, Lignincola |
| |
|
|
Microascaceae |
Wardomycopsis |
| |
|
|
Triadelphiaceae |
Triadelphia |
| |
|
|
Ceratocystidaceae |
Ceratocystis, Thielaviopsis |
| |
|
Pararamichloridiales |
Pararamichloridiaceae |
Pararamichloridium |
| |
Savoryellomycetidae |
Conioscyphales |
Conioscyphaceae |
Conioscypha |
| |
|
Pleurotheciales |
Pleurotheciaceae |
Monotosporella |
| |
|
Savoryellales |
Savoryellaceae |
Ascotaiwania, Canalisporium, Savoryella |
| |
Sordariomycetidae |
Chaetosphaeriales |
Chaetosphaeriaceae |
Chaetosphaeria, Chloridium, Codinaea, Craspedodidymum, Cryptophiale, Dictyochaeta, Kionochaeta, Rattania, Sporoschisma, Thozetella |
| Sordariomycetes (cont.) |
Sordariomycetidae (cont.) |
Chaetosphaeriales (cont.) |
Helminthosphaeriaceae |
Endophragmiella |
| |
|
|
Leptosporellaceae |
Leptosporella |
| |
|
|
Linocarpaceae |
Linocarpon, Neolinocarpon |
| |
|
|
Incertae sedis |
Caudatispora |
| |
|
Coniochaetales |
Incertae sedis |
Cannonia |
| |
|
Meliolalles |
Meliolaceae |
Meliola |
| |
|
Phyllachorales |
Phaeochoraceae |
Cocoicola, Phaeochora, Phaeochoropsis, Serenomyces |
| |
|
|
Phyllachoraceae |
Brobdingnagia, Camarotella, Coccodiella, Coccostromopsis, Maculatifrondes, Malthomyces, Ophiodothella, Oxodeora, Phyllachora, Sphaerodothis, Tribulatia |
| |
|
Sordariales |
Chaetomiaceae |
Trichocladium |
| |
|
|
Lasiosphaeriaceae |
Cercophora, Lasiosphaeria |
| |
|
|
Incertae sedis |
Lockerbia |
| |
|
Incertae sedis |
- |
Arecacicola, Curvatispora, Nigromammilla, Paracapsulospora |
| |
Xylariomycetidae |
Amphisphaeriales |
Amphisphaeriaceae |
Amphisphaeria, Lepteutypa |
| |
|
|
Apiosporaceae |
Arthrinium, Dictyoarthrinium |
| |
|
|
Appendicosporaceae |
Appendicospora |
| |
|
|
Beltraniaceae |
Beltrania |
| |
|
|
Hyponectriaceae |
Arecomyces, Frondicola, Hyponectria, Rachidicola |
| |
|
|
Iodosphaeriaceae |
Iodosphaeria |
| |
|
|
Oxydothidaceae |
Oxydothis |
| |
|
|
Pseudomassariaceae |
Leiosphaerella, Pseudomassaria |
| |
|
|
Sporocadaceae |
Bartalinia, Morinia, Neopestalotiopsis, Pestalotiopsis, Pseudopestalotiopsis, Robillarda, Seiridium |
| |
|
Xylariales |
Barrmaeliaceae |
Barrmaelia |
| Sordariomycetes (cont.) |
Xylariomycetidae (cont.) |
Xylariales (cont.) |
Cainiaceae |
Arecophila, Seynesia, Endocalyx |
| |
|
|
Clypeosphaeriaceae |
Apioclypea, Brunneiapiospora, Palmaria |
| |
|
|
Diatrypaceae |
Allocryptovalsa, Allodiatrype, Anthostoma, Cryptovalsa, Diatrype, Diatrypella, Eutypa, Eutypella, Frondisphaeria, Peroneutypa |
| |
|
|
Fasciatisporaceae |
Fasciatispora |
| |
|
|
Graphostromataceae |
Biscogniauxia |
| |
|
|
Hansfordiaceae |
Hansfordia |
| |
|
|
Hypoxylaceae |
Annulohypoxylon, Hypoxylon |
| |
|
|
Microdochiaceae |
Idriella, Microdochium |
| |
|
|
Oxydothidaceae |
Oxydothis |
| |
|
|
Robillardaceae |
Robillarda |
| |
|
|
Xylariaceae |
Anthostomella, Ascotricha, Astrocystis, Diabolocovidia, Kretzschmaria, Nemania, Neoxylaria, Rosellinia, Stilbohypoxylon, Xylaria |
| |
|
|
Zygosporiaceae |
Zygosporium |
| |
|
|
Incertae sedis |
Capsulospora, Circinotrichum, Cyanopulvis, Gyrothrix, Guestia, Haploanthostomella, Lasiobertia, Neobarrmaelia, Nipicola, Palmicola, Pemphidium, Pulmosphaeria, Sabalicola |
| |
|
Incertae sedis |
Myelospermataceae |
Myelosperma |
| |
|
|
- |
Frondispora, Manokwaria |
| |
Incertae sedis |
Catabotryales |
Catabotryaceae |
Catabotrys |
| |
|
- |
Acrodictyaceae |
Acrodictys |
| |
|
- |
- |
Apogaeumannomyces, Flammispora, Mangrovispora |