Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Maria Antonia De Francesco and Version 2 by Jason Zhu.
Infections due to the Aspergillus species constitute an important challenge for human health. Invasive aspergillosis represents a life-threatening disease, mostly in patients with immune defects. Drugs used for fungal infections comprise amphotericin B, triazoles, and echinocandins. However, in the last decade, an increased emergence of azole-resistant Aspergillus strains has been reported, principally belonging to Aspergillus fumigatus species. Therefore, both the early diagnosis of aspergillosis and its epidemiological surveillance are very important to establish the correct antifungal therapy and to ensure a successful patient outcome.
- drug
- resistance
- Aspergillus
- azole
1. Introduction
Aspergillus spp. are filamentous fungi found ubiquitously in the environment, in places such as soil, decaying vegetative material, and dust [1]. Furthermore, the fungi might colonize oligotrophic water systems: more than 400 different species have been found to inhabit different water sources [2], underlining that as well as air, water might also be a potential source of the transmission of filamentous fungi [3].
In different geographical areas, climatic factors such as humidity, rainy season, and temperature influence the prevalence of Aspergillus spp. [4].
The inhalation of Aspergillus conidia gives rise to different respiratory infections, affecting immunocompromised patients more severely.
Despite great advances in the diagnosis and treatment of aspergillosis, mortality remains high, particularly in subjects with important immune defects and invasive diseases.
Infections are generally due to Aspergillus fumigatus, even if other species are increasingly detected as etiological agents [5].
Recently, Aspergillus fumigatus was added to the list of the 19 fungal pathogens to be prioritized by the World Health Organization (WHO) and inserted into the critical group together with Cryptococcus neoformans, Candida auris, and Candida albicans [6].
Antifungal resistance is an emerging and important challenge in different parts of the world, with up to 20% of Aspergillus isolates displaying de novo resistance to commonly used antifungal drugs [7].
2. Amphotericin B Resistance
Amphotericin B acts by interacting with sterols, in particular with ergosterol, the principal component of the fungal cell membrane. The integration of amphotericin B into the fungal membrane leads to the formation of channels (Figure 1). This formation impairs barrier membrane function, increasing the permeability responsible for the leakage of potassium, protons, cations, and cytoplasmic materials that induce cell death [8][74]. Furthermore, amphotericin B can produce reactive oxygen species (ROS) that give rise to cellular damage [9][75]. Among Aspergillus spp., Aspergillus terreus is the strain harboring intrinsic resistance to amphotericin B [10][76].
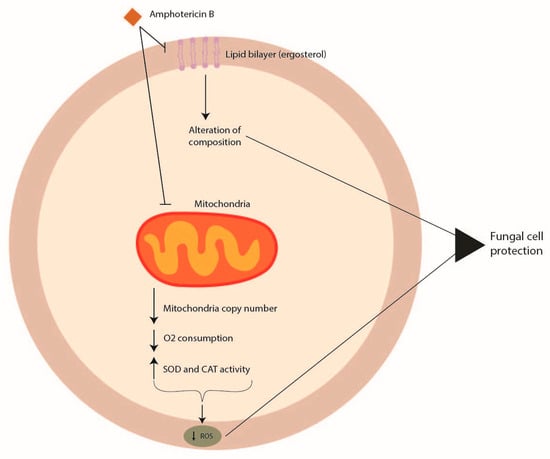
Figure 1. Mechanisms of Aspergillus spp. antifungal drug resistance: amphotericin B resistance.
The mode of action of amphotericin B in Aspergillus terreus has not been well elucidated, and no genomic features have been identified to date that might be linked to amphotericin B resistance. However, amphotericin B resistance in section Terrei seems to be associated with the modulation of molecular chaperones, targeting ROS by mitochondria and influencing cellular redox homeostasis, with an increase in the level of catalase and superoxide dismutase with respect to other Aspergillus species [10][11][12][76,77,78] (Figure 1).
Following the increasing rate of azole resistance [13][79], the use of amphotericin B has enhanced, and this might be the reason why, recently, an increase in MIC values for amphotericin B in different Aspergillus species has been reported [14][15][16][17][80,81,82,83], even if resistance to this drug remains extremely rare [18][84].
In a recent study [19][85], among 26,909 Aspergillus isolates analyzed, resistance to amphotericin B was detected in 36.8% of Aspergillus terreus, 14.9% of Aspergillus flavus, 5.2% of Aspergillus niger, and 2.01% of Aspergillus fumigatus isolates. Furthermore, some Aspergillus lentulus and Aspergillus ustus isolates have been reported to show amphotericin B resistance [20][21][86,87]. Additionally, an increasing trend in amphotericin B resistance was observed in Aspergillus fumigatus between 2016 and 2020, together with a decreasing trend in amphotericin B resistance in Aspergillus terreus and Aspergillus flavus [19][85].
3. Azole Resistance
Azole antifungal drugs act by interfering with the synthesis of ergosterol, mediated by the fungal sterol 14 alpha-demethylases (Cyp51A and Cyp51B) (Figure 2).
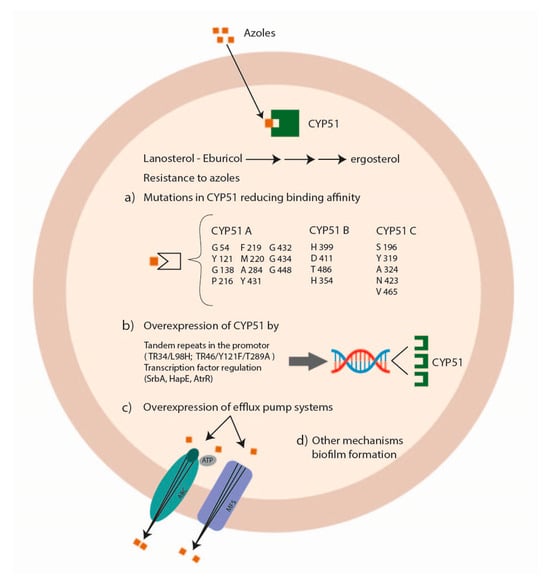
Figure 2. Mechanisms of Aspergillus spp. antifungal drug resistance: azole resistance.
The principal mechanisms of azole resistance involve (a) mutations in the sterol-demethylase gene cyp51A reducing the affinity between the azole drug and its target; (b) the overexpression of the sterol-demethylase gene cyp51A leading to an increase of the azole concentration able to inhibit fungal growth; (c) the overexpression of efflux pump systems decreasing the intracellular drug concentration [22][23][88,89]. Alternate mechanisms associated with azole resistance are the modification of HapE [24][90], the involvement of the mitochondrial complex 1 and the cytochrome b5-CybE redox systems [25][26][91,92], the transcription factors SrbA and AtrR [27][28][93,94] and biofilm formation (Figure 3) [29][95].
The acquisition of azole resistance occurs in two ways: in vivo, by selection of resistant isolates during long therapy with azoles, and in vitro, by the selection of resistant isolates as a consequence of an extensive use of azole fungicide in agriculture [30][31][32][96,97,98].
3.1. Mutations in the Sterol-Demethylase Gene Cyp51
Cyp51 genes are the major targets studied for azole resistance in fungal pathogens—resistance that might also be acquired by horizontal gene transfer (HGT) [33][99]. Different species of Aspergillus exhibit several numbers of cyp51 paralogs in their genome. This genetic redundancy gives the fungus an advantage to survive when exposed to fungicides, increasing its azole resistance [34][100]. In Aspergillus fumigatus, Aspergillus terreus, and Aspergillus niger, there are only two paralogs (cyp51A and cyp51B), while in Aspergillus flavus, three paralogs exist (cyp51A, cyp51B and cyp51C) [35][101]. However, only specific mutations in Cyp51A and Cyp51C proteins have been shown to have an impact on azole resistance, while the Cyp51B protein might play other roles that need to be studied in detail [36][102].
In Aspergillus fumigatus, amino-acid mutations in the Cyp51A protein related to azole resistance were G54, Y121, G138, P216, F219, M220, A284, Y431, G432, G434, and G448 [37][38][39][40][41][42][43][103,104,105,106,107,108,109]. No mutations were associated with azole resistance in the CypB protein [22][44][88,110].
Also, in Aspergillus lentulus, it was demonstrated by targeted cyp51A gene knockout that intrinsic azole resistance was related to this gene [45][111].
In Aspergillus flavus with reduced voriconazole susceptibility, different mutations were reported in Cyp51A, Cyp51B, and Cyp51C proteins, even if their role in azole resistance needs to be clarified by further studies. Amino-acid changes in Cyp51A protein were identified at positions R450S, K197N/D282E/M288L, and Y132N/T469S [46][112]; in Cyp51B protein, detected mutations were H399P, D411N, T454P, T486P 105, and Q354K [47][113]; in Cyp51C protein, the Y319H mutation was identified from an azole-resistant clinical isolate [48][114] and the mutations S196F, A324P, N423D, and V465M [49][50][115,116].
In section Nigri, in Cyp51A protein, many mutations were identified, but their role in inducing azole resistance is still uncertain [51][117].
In particular, in Aspergillus tubingensis, a recent study found the amino-acid change H467Q in combination with the mutations K64E or V377I exclusively in non-wildtype isolates [52][118], while in Aspergillus niger, no mutation is associated with azole resistance [52][118].
However, it has been found that single-gene deletions of cyp51A and cyp51B genes in Aspergillus tubingensis and Aspergillus niger decrease the voriconazole MIC values below the ECV established by CLSI [53][119].
In Aspergillus braziliensis with reduced azole susceptibility, there were no mutations present in Cyp51A protein [52][118].
Regarding the Terrei section, few studies are available in the literature about their azole resistance; only a mutation of methionine in position 217 in Aspergillus terreus has been reported [54][55][120,121].
In the Usti section, the intrinsically azole-resistant species Aspergillus calidoustus exhibited a mutation M220V in Cyp51A protein, a position already associated with azole resistance in Aspergillus fumigatus [22][88].
3.2. Overexpression of the Sterol-Demethylase Cyp51
The overexpression of Cyp51 is considered another mechanism of azole resistance in Aspergillus spp., in particular in Aspergillus fumigatus.
Regarding Aspergillus flavus, the overexpression of Cyp51A and Cyp51B was not related to azole resistance because the levels of gene expression were the same both in wild and non-wildtype strains [56][57][122,123]. Also, in the Nigri section, this mechanism seems not to be related to azole resistance, even if Cyp51A is upregulated after azole exposure [52][58][118,124].
Changes in the promoter region of cyp51 have been described as a mechanism to contrast azole toxicity, mostly in Aspergillus fumigatus, such as the insertion of tandem repeats (TR) of 34bp, 46bp, and 53 bp leading to upregulation of cyp51 [59][125]. These insertions were often associated with amino-acid substitutions in Cyp51A and have been observed in strains that exhibit reduced susceptibility to azoles [60][126].
In particular, the TR34/L98H mutation was found principally in resistant environmental isolates, linked to total resistance to itraconazole and reduced susceptibility to voriconazole and posaconazole [23][61][62][89,127,128].
Furthermore, the TR46/Y121F/T289A mutation was found in isolates with high levels of resistance to voriconazole and other azoles [63][129]. For the tandem repeat of 53 bp, no amino-acid substitution has been found to date [62][64][128,130].
Other transcription factors were found able to regulate cyp51 expression in Aspergillus fumigatus, such as SrbA, HapE, and AtrR.
SrbA is a transcriptional regulator that is involved in different processes, as well as sterol biosynthesis [65][66][67][131,132,133], and its deletion leads to greater azole susceptibility [27][93].
The mutation P88L in heme activator protein E (HapE), a subunit of the CCAAT-binding transcription factor complex (CBC), determines an upregulation of the cyp51A gene and confers azole resistance [68][134]. A decrease in CBC activity, a negative regulator of the ergosterol pathway, increases the expression of the enzymes involved in its biosyntheses, such as HMG-CoA-synthase, HMG-CoA-reductase, and sterol C14-demethylase contributing to azole resistance [24][90].
Furthermore, a Zn2Cys6 cluster-containing transcription factor, called ABC transporter regulator or AtrR, was found to be important for azole tolerance both in Aspergillus fumigatus and Aspergillus flavus [69][70][135,136]. Additionally, cytochrome b5CybE has been found to be able to regulate the expression level of cyp51A [26][92].
Furthermore, biofilm production was also hypothesized to play a role in the azole resistance of Aspergillus fumigatus. The cell density reached in a mature biofilm, together with the production of polysaccharide extracellular matrix, protects the mold by the action of the immune system and the antifungal drugs [30][96].
3.3. Overexpression of Efflux Pump Systems
Efflux pumps are transmembrane proteins that expel drugs from the cell, reducing their intracellular concentration. Therefore, the overexpression of these proteins might be related to azole resistance. The principal types of efflux pumps are the ATP-binding cassette (ABC) transporter and the major facilitator superfamily (MFS) transporter, which differ in structure and activity [71][137]. ABC transporters use ATP as an energy source, while MFS transporters use a proton gradient to pump out the drugs [72][138].
The more studied ABC transporters involved in azole resistance are cdr1B, mdr1, mdr2, mdr3, mdr4, abcD, abcE, atrI, atrB, atrC, and atrF. In Aspergillus fumigatus and Aspergillus flavus, only the ABC transporter Cdr1B has been found to be related to azole resistance [73][74][139,140].
MFS transporters were less studied, and among them, only mdrA has been associated with an increase of itraconazole and voriconazole susceptibility in Aspergillus fumigatus [75][141].
4. Echinocandin Resistance
Echinocandins (caspofungin, anidulafungin, and micafungin) act by inhibiting the glucan synthase, an enzyme codified by the FKS1 and FSK2 genes, important for the synthesis of the beta1, 3 glucan (Figure 3). However, because echinocandins have only a fungistatic activity against Aspergillus spp., they are used only in combination with a polyene or an azole to obtain an important synergistic effect. To date, the echinocandin resistance is rarely found in Aspergillus spp. [76][142].
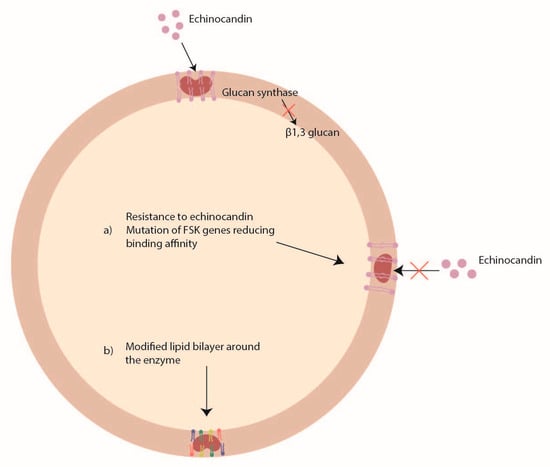
Figure 3.
Mechanisms of
Aspergillus
spp. antifungal drug resistance: echinocandin resistance.
Some studies have reported mutations in FSK genes [77][143] and changes in the lipid profile around the enzyme [78][144] as possible mechanisms of echinocandin resistance (Figure 3). In Aspergillus fumigatus, two mutations (S678P and E671Q) in the FSK1 gene were associated with echinocandin resistance [79][80][145,146], while in Aspergillus flavus it was shown that P-type ATPase and ubiquinone biosynthesis methyltransferase COQ5 might be involved in caspofungin resistance [81][147].