1. Introduction
Xylan is the major constituent of hemicellulose in plant cell walls
[1]. It has a complex and heterogeneous structure with a linear chain composed of xylose units linked together via β-1,4 glycosidic bonds
[2]. The xylose backbone is variably substituted with a range of sugars and/or organic acids, restricting the accessibility of the enzyme to the target site. Endo-xylanases (EC 3.2.1.8) randomly hydrolyze the β-1,4 glycosidic linkages in the xylan backbone, converting the polymeric xylan into xylooligosaccharides with different degrees of polymerization and the xylose monomer
[3]. Chemical and structural information on glycoside hydrolases (GHs) associated with xylanases is available in the carbohydrate-active enzyme database (CAZy). This database classifies relevant enzymes based on sequence similarity
[4]. The major GH families associated with xylanase are 5, 7, 8, 9, 10, 11, 12, 16, 26, 30, 43, 44, 51, and 62
[5]. Enzymes in GH families 5, 7, 8, 10, 11, and 43 have a single distinct catalytic domain, whereas those in GH families 16, 51, and 62 have two catalytic domains with bifunctional properties
[6]. Enzymes in GH families 9, 12, 26, 30, and 44 have secondary xylanolytic action. Moreover, according to the CAZy database, most endo-xylanases belong to GH families 10 and 11. Although the GH11 family is considered the only “true xylanase” family that has high specificity and selectivity toward xylan substrates, it lacks cellulase or xylosidase activity
[7][8][9][7,8,9]. GH11 xylanases have high application value owing to their catalytic activity and broad temperature/pH spectra
[10].
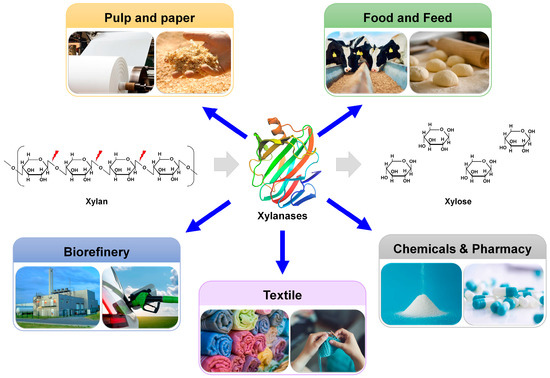
Xylanases are used as biocatalysts in multiple industrial fields, including the paper/pulp, food/feed, textile, chemical/pharmaceutical, and biorefinery industries
[11][12][11,12] (
Figure 1). During the bleaching of Kraft pulp in the paper industry, the lignin in wood chips is removed through sequential treatment with chlorine, chlorine dioxide, and NaOH
[13]. Chlorine and chlorine dioxide generate persistent organic chemicals that are toxic to organisms and pose risks to human health. The pretreatment of paper pulp with xylanases increases the chemical lignin extraction efficiency and reduces the amount of chlorine and chlorine dioxide required for extraction. Therefore, xylanases are frequently used for the biobleaching of pulp, thereby improving final paper quality
[11]. In the food industry, xylanases can be added to baking dough to improve the quality of baking products by enhancing gluten elasticity, texture, and moisture content
[14]. In the feed industry, xylanases are applied as feed ingredients to improve digestibility and produce bioactive prebiotic xylooligosaccharides
[15]. In biorefinery, lignocellulosic biomass is converted to various industrially valuable materials, including sugars, fuels, and chemicals
[16]. Xylanases are further involved in the saccharification process of lignocellulose biorefinery, contributing to the production of fermentable sugars
[17].
Figure 1.
Industrial application of xylanases.
GH11 xylanases are highly desirable for these industrial applications, particularly because they lack cellulase activity. These bioprocesses often require extreme conditions in terms of pH, temperature, and inhibitors
[14][17][14,17]. For example, pulp bleaching involves heat (70 °C) and alkalinity (pH > 9), requiring thermoalkaline enzymes
[14]. In biotechnological processes, such as baking, fruit juice clarification, or xylo-oligosaccharides production, xylanases with high activity at acidic pH are preferred
[11][18][11,18]. When using yeast that expresses xylanase for cellulosic ethanol production, enzymes that are tolerant to toxic solvents, such as ethanol and acetate, are preferred
[19][20][19,20].
Protein engineering is a multidisciplinary scientific field that focuses on protein design, modification, and optimization to improve their characteristics or achieve specific functions
[21]. Protein engineering, guided by directed evolution (DE) and (semi)rational engineering, has emerged as a potent technology with the potential to advance the biotechnology era
[22]. Powered by recent outstanding advancements in artificial intelligence (AI) technologies, AlphaFold, a cutting-edge AI system developed by DeepMind, has revolutionized structural biology by rapidly and accurately predicting the 3D structure of a protein directly from its amino acid sequence
[23]. It can provide a wealth of structural information and enhance
theour understanding of the structural basis of protein function. Thus, it enables researchers to make more informed decisions in the design and modification of proteins. Moreover, AI-assisted powerful tools for designing and creating high-quality and new proteins de novo have been recently developed, exemplified by ProteinMPNN
[24].
Xylanases with high activity and stability under extreme conditions have been screened from various extremophiles or developed based on DE or rational design (RD)-guided protein engineering
[25][26][27][28][29][25,26,27,28,29]. However, there is limited information on semi-RD-based xylanase engineering, which often involves computational prediction followed by site-saturation mutagenesis. Particularly, xylanases belonging to the GH11 family are mostly sourced from non-extremophiles
[30] and require engineering to become suitable for harsh bioprocessing conditions.
The temperature and pH-related biochemical properties of GH11 xylanases are important factors to consider to achieve a feasible and eco-friendly bioprocess. In particular, by shifting the optimal pH, altering the pH profile, and ensuring pH stability, these enzymes can effectively carry out their intended functions, leading to improved efficiency in various relevant industrial processes. To engineer the pH-associated features of GH11 enzymes, various strategies, such as site-directed mutagenesis
[31][32][31,32], RD, modeling/site-directed mutagenesis, structural comparison, redesign of electrostatic potential
[33], structure analysis
[34][35][34,35], and bioinformatics- and biostatistics-based approaches
[36], have been used. In a recent review, comprehensive up-to-date strategies for creating a diverse xylanase gene library; implementing high-throughput systems to screen enhanced xylanase mutants derived by DE; and utilizing in silico methods for the prediction based on, for example, Framework for Rapid Enzyme Stabilization by Computational libraries (FRESCO) and in-depth analysis of target mutations were thoroughly examined
[37] to obtain desirable industrial characteristics, such as alkaliphilic enhancement, thermal stability, and catalytic performance. Nevertheless, protein engineering studies that involve altering the pH profile and improving pH stability are relatively scarce compared with the efforts put into engineering temperature-related properties, such as improving xylanase thermostability. Furthermore, to date, there is no systematic review on this topic.
2. Functions and Structures of GH11 Xylanases
2.1. Catalytic Mechanisms
Hydrolysis by GHs occurs through a retaining or inverting mechanism
[38]. Enzymatic hydrolysis by GH family members requires two acidic residues (glutamate (Glu) or aspartic acid (Asp)) to act as a proton donor and nucleophile/base, respectively
[38]. The GH11 family utilizes a retention mechanism catalyzed by two conserved Glu residues. Xyl-11 from
Bacillus sp. KT12 is the most widely used model for the GH11 catalytic mechanism investigation. The history of the catalytic mechanism of Xyl-11 identification is well documented
[7].
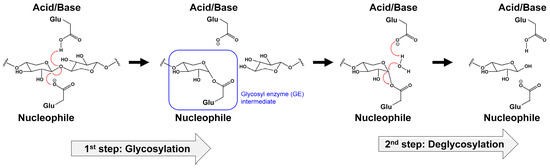
The two catalytic Glu residues of Xyl-11 catalyze a double-displacement mechanism involving a covalent glycosyl enzyme intermediate
[39][40][41][39,40,41]. The retaining mechanism of GH11 can be divided into two main steps: glycosylation and deglycosylation (
Figure 2). In the first step (glycosylation), the acid–base Glu residue acts as an acid and protonates the substrate. The nucleophile Glu residue facilitates the departure of the leaving group and the formation of the α-glycosyl enzyme intermediate during the oxocarbenium-ion-like transition state. In the second step (deglycosylation), the acid–base Glu residue acts as a base and abstracts a proton from a water molecule. Subsequently, the molecule attacks the anomeric carbon of the α-glycosyl enzyme intermediate. Furthermore, a second substitution occurs, where the anomeric carbon produces a product with the β configuration identical to that of the substrate (
Figure 2).
Figure 2.
Retaining catalytic mechanism of GH11 xylanase with double-displacement of two glutamate residues.
This traditionally identified retaining mechanism of GH11 xylanases is consistent with recent computational analysis findings. This analysis used ab initio quantum mechanics/molecular mechanics calculations with a neutron crystal structure that supported the glycosylation reaction mechanism steps. It revealed the protonation states of the residues involved in the reaction mechanism
[42].
2.2. Structures
To date, more than 100 GH11 xylanase structures from various organisms have been deposited in the Protein Data Bank (PDB) (
https://www.rcsb.org/, accessed on 9 August 2023). Representative crystal structures of GH11 xylanases from various organisms are listed in
Table 1.
Table 1.
Xylanase GH11 xylanase structures.
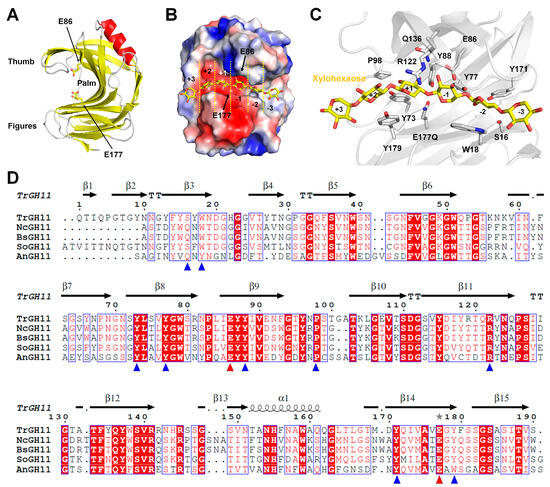
GH11 xylanases have a β-jelly roll structure consisting of two antiparallel β-sheets connected by interconnecting loops
[7] (
Figure 3A). A unique α-helix is packed underneath one side of the β-sheet. Numerous hydrogen bonds between the β-strands stabilize the framework. The two β-sheets resemble palms and fingers, whereas loops β12-β13 resemble a thumb-like domain
[7]. The β-sheets in the finger domain are almost planar, whereas the β-sheets in the thumb domain are partially twisted to form vertical angles. Long (~25 Å), narrow (~4 Å), and deep (~9 Å) clefts are located between the finger and thumb domains and form the substrate-binding site. The substrate-binding site of GH11 xylanases contains six subsites (
Figure 3B), and the catalytic dyad consisting of two Glu residues is located near the center of the substrate-binding pocket. These two Glu residues are positioned facing each other, near the subsite between +1 and –1 (
Figure 3B). The narrowest zone of the cleft is located between the conserved Pro residue of the thumb and the conserved aromatic residue (e.g., Trp or Tyr) of the β-strand on the finger domain. Within the substrate-binding cleft, mainly aromatic residues with a few polar residues are located (
Figure 3C). The overall architecture, topology, and catalytic cleft residues of GH11 xylanases are widely conserved
[7].
Figure 3. GH11 xylanases structure and sequence alignments of GH11 family members: (A) crystal structure of GH11 from Trichoderma reesei (PDB code: 1ENX); (B) electrostatic surface of TrGH11 with xylohexose; (C) close-up view of the active site of TrGH11 complexed with xylohexose (PDB code: 4HK8); (D) structure-based sequence alignment of GH11 from Trichoderma reesei (TrGH11, UniProt: P36217), Niallia circulans (NcGH11, P09850), Bacillus subtilis (BsGH11, P18429), Streptomyces olivaceoviridis (SoGH11, A0A7G1MBT0), and Aspergillus niger (AnGH11, UniProt: P55329).
GH11 from
Trichoderma reesei is the most structurally studied model among the GH11 family deposited in the PDB (
Table 1). The crystal structures of native TrGH11 and its complexes with 4,5-epoxypentyl beta-D-xyloside (PDB code: 1RED), 3,4-epoxybutyl beta-D-xyloside (1REE), 2,3-epoxypropyl beta-D-xyloside (1REF), 2,6-pyridinedicarboxylic acid (3LGR), xylohexaose (4HK8), and xylotriose (4HK9) have been reported. These structures tentatively provide insights into the substrate recognition or inhibitor of GH11. In particular, the crystal structure of TrGH11 complexed with xylohexaose provides detailed information about substrate recognition (
Figure 3C). TrGH11–xylohexaose was able to maintain the xylohexaose complex by replacing the catalytic residue E177 with glutamine. In TrGH11-E177Q, the xylohexaose substrate interacted with residues S16, W18, Y77, Y88, E86, P98, R122, Q136, Y171, Y179, and E177Q (
Figure 3C). This complex structure indicates where and how substrates are recognized at the six subsites of GH11. Therefore, it potentially provides insight into protein engineering for amino acids that may be involved in altering GH11 substrate specificities. TrGH11 substrate recognition residues are highly conserved with other GH11 from
Niallia circulans,
Bacillus subtilis,
Streptomyces olivaceoviridis, and
Aspergillus niger (
Figure 3D). Conversely, TrGH11 S16 and W18 are not absolutely conserved. This result suggests differences in substrate binding and recognition at subsites −2 and −3 for each GH11 (
Figure 3D).
3. Engineering of GH11 Xylanases for Optimal pH Shifting
Protein engineering involves modifying a protein sequence by substituting, inserting, or deleting nucleotides in the encoding gene to obtain a modified protein that is more suitable for a specific use or purpose. In targeted or site-directed mutagenesis, a specific site within a gene sequence is altered
[43]. To shift the optimal pH of GH11 xylanases through engineering, various protein engineering strategies have been performed. These include site-directed mutagenesis
[31][32][31,32], RD, modeling/site-directed mutagenesis, structural comparison, redesign of electrostatic potential
[33], structure analysis
[34][35][34,35], and bioinformatics- and biostatistics-based approaches (
Table 2)
[36]. In what follows,
thwe re
searchers review and discuss important protein engineering achievements in shifting the optimal pH of GH11 xylanases for industrial applications. In particular,
the researchers cwe carefully examine the correlation between variant amino acids and functions in engineered GH11 using crystal and modeled structures.
Table 2.
Engineering of GH11 xylanases for optimal pH shifting.
|
Enzyme
(Wild-Type)
|
Source
|
pH Optimum/Stability
|
Condition
|
Mutation
Approach
|
Reference
|
|
XynA
|
Bacillus subtilis
|
pH 6 → 6.5
|
80 °C
|
DE
(epPCR and
DNA shuffling)
|
|
|
|
| pH 7 → 6 |
|
65 °C, alkali-pretreated rice straw
|
in silico analysis
|
[10]
|
|
TlXynA
|
Thermomyces
lanuginosus
|
acid tolerance
alkali tolerance
|
65 °C,
|
RD
|
[46]
|
|
BaxA
|
Bacillus amyloliquefaciens
|
pH 6 (broad pH spectrum) → 7 (narrow pH spectrum)
|
50 °C
|
DE
(EPTD-PCR)
|
[47]
|
|
BCX
|
Bacillus circulans
|
pH 6.5 → 5–6
|
50 °C
|
RD
|
[18]
|
|
XynB
|
Thermotoga maritima
|
pH 5.1 → 5.5, overall pH shift upwards by 0.5
|
90 °C
|
RD
|
[48]
|
|
CbX-CD
|
Caldicellulosiruptor bescii
|
pH 6.5 → 5.0
|
70 °C
|
RD
Biomathematics and biostatistics
|
[36]
|
|
XynB
|
Aspergillus niger
|
pH 5.0 → 5.5
|
50 °C
|
Biomathematics
RD
|
[31]
|