Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Jason Zhu and Version 1 by Diego Estrada-Luna.
Inflammation and oxidative stress are essential components in a myriad of pathogenic entities that lead to metabolic and chronic diseases. Moreover, inflammation in its different phases is necessary for the initiation and maintenance of a healthy pregnancy. Therefore, an equilibrium between a necessary/pathologic level of inflammation and oxidative stress during pregnancy is needed to avoid disease development. High-density lipoproteins (HDL) are important for a healthy pregnancy and a good neonatal outcome. Their role in fetal development during challenging situations is vital for maintaining the equilibrium.
- inflammation
- oxidative stress
- pregnancy
- high-density lipoproteins
1. Introduction
Lipid metabolism during pregnancy becomes relevant since lipid concentrations change according to maternal requirements and fetal growth; subsequently, dysregulation of lipid metabolism is associated with endothelial dysfunction or immunological changes [150][1], while major alterations are found in the concentration of triglycerides and cholesterol and in the number of LDL and HDL particles [157][2], hyperlipidemia being a common condition even in normal pregnancy that allows glucose and calories to be utilized by the fetus. Nevertheless, it has been reported that maternal lipid levels during pregnancy are significantly correlated with the lipid profile of children during the first years of life [158][3]. Additionally, it is well known that lipid dysregulation is an important risk factor associated with the development of preeclampsia and cardiovascular disease in pregnancy.
2. Lipoproteins
Lipoproteins are macromolecular complexes composed of hydrophobic lipids such as triglycerides and cholesterol esters on the inside, whereas their surface is formed by amphipathic lipids like phospholipids and free cholesterol. Moreover, there are proteins, known as the apolipoproteins (Apo), providing stability to the surface and conferring part of their own properties [5][4] (Figure 1). In recent decades, the role that certain lipoproteins play in different chronic-degenerative diseases has sparked interest, especially high-density lipoproteins (HDL).
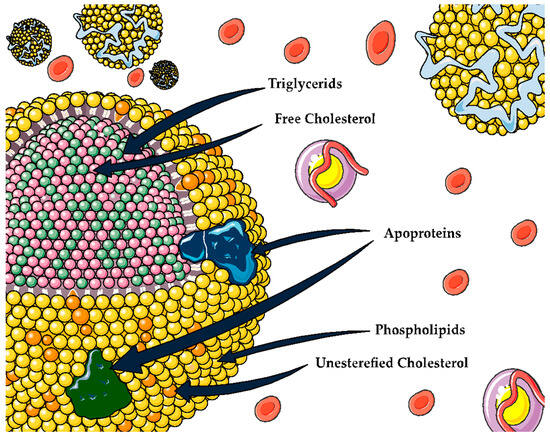
Figure 1. General structure for lipoproteins.
3. HDL
High-density lipoproteins (HDL) are complex and heterogeneous structures constituting a lipid transport mechanism. Different components (lipids and proteins) of HDL are continuously being exchanged. As a result, they modify the composition, charge, and size of these particles. Currently, it has been described that HDL particles can similarly transport other compounds (about 250), such as sphingosine-1-phosphate, paraoxonase-1 (PON1), acute phase proteins (SAA) [159][5], platelet-activating factor acetylhydrolase (PFA-AH) enzymes and proteins, such as cholesterol ester transporter protein (CETP) and phospholipid transporter protein (PLTP), among many other components [160,161][6][7].
In this context, HDL has been attributed to exert some cardioprotective properties, including reverse cholesterol transport and antioxidant, anti-inflammatory, and antiatherogenic activities (Table 1) [162,163,164,165][8][9][10][11]. Many of these functions are important for a healthy pregnancy and good neonatal outcomes [166,167][12][13].
Table 1. HDL main functions.
| HDL Function | Proteins and Lipids Associated with HDL Function | Ref. | |
|---|---|---|---|
| Reverse cholesterol transport | HDL promotes cholesterol efflux from various cell types. | ABCAI, ABCG1, SR-BI, cubilin, ApoE receptor | [168,169][14][15] |
| Removing excess cholesterol from lipid-laden macrophages is a crucial process in HDL-mediated vascular protection. | |||
| Oxidant | HDL has antioxidant properties whereby it can remove and inactivate lipid peroxides from LDL and cells. | PON 1, Apo AI, PAF-AH, LCAT, Apo M, S1P, phospholipids | [159][5] |
| Inflammation | Controlling the activation of monocytes, preventing macrophage migration, and inhibiting the oxidation of LDL by blocking the 12-lipoxygenase that produces lipid hydroperoxides and leads to the oxidation of the LDL | VCAM, ICAM, TNF-α, SAA, ceramides | [169][15] |
| Vascular function | Modulation of endothelial nitric oxide synthase (eNOS) expression, leading to increased nitric oxide (NO) production and vasodilation | ABCA1, SR-BI, S1PR, S1P, Apo M | [170,171,172][16][17][18] |
These positive effects are explained by the structure and chemical composition of these particles. However, it has been shown that these lipoproteins can lose or reduce their cardioprotective capacity, giving rise to prooxidant, proinflammatory, and proatherogenic lipoproteins, contributing to the process of atherosclerosis; this phenomena has been termed “dysfunctional HDL” [173,174][19][20].
Some study groups proposed the hypothesis that HDL delivers lipids to cells. For instance, Pérez-Mendez et al. demonstrated that HDL delivers cholesterol and sphingomyelin to endothelial cells in vitro [175][21]. Therefore, the possibility of the regulation of these lipoproteins on cell function after internalization and the delivery of their content is extremely high. Hence, the lipid delivery of HDL to cells becomes of particular importance when cell membranes should be intensively synthesized or re-structured, i.e., during fetal growth. HDL-C plasma levels and composition may change drastically during inflammatory processes.
It has been described that HDL can inhibit the oxidation of other molecules, such as LDL through free radical damage, which results in the generation of oxidized lipids with pro-inflammatory activity [176][22]. Nonetheless, in certain conditions such as obesity, diabetes, and other cardiovascular diseases, it has been observed that HDL loses its protective properties, becoming dysfunctional HDL [168[14][15][16],169,170], and leads to an increase in inflammatory processes and OS in several conditions, including pregnancy (Figure 2) [166,167][12][13].
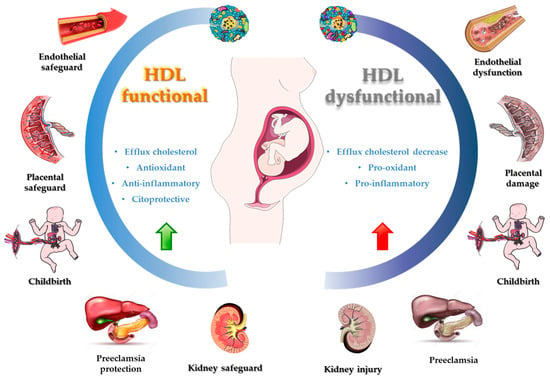
Figure 2. Functional and dysfunctional HDL during pregnancy.
Given that this is of great relevance in chronic degenerative diseases, the use of bioactive compounds from fruits, vegetables, foods of animal origin, and plants has become an alternative for improving the functionality and chemical composition of HDL. Thus, they are able to regulate the negative effects caused by OS and inflammatory processes.
4. HDL Role and Upregulation Contribution in Inflammatory Processes and OS
Pregnant women normally experience physiological changes, involving carbohydrate and lipid metabolism, insulin resistance, inflammation, coagulation, and OS, all of them causing endothelial damage [170][16]. Despite this unfavorable environment, pregnant women have better vascular function. Likewise, during embryogenesis and fetal development, the levels of apolipoproteins, lipoproteins, and lipids increase significantly. HDL-C levels change during pregnancy: in the first trimester, changes are insignificant, but, in the second trimester, these changes increase and then slightly decrease in the third trimester. In chronic inflammatory processes, the functional activities of HDL are reduced, the formation of new particles decreases, and catabolism increases. Also, structural changes occur at the protein level, such as the replacement of PON1 or Apo A1 molecules by proinflammatory proteins, including ceruloplasmin and SAA [177,178][23][24] that converts HDL to HDL-proinflammatory and results in increased chemoattractant activity, oxidation of LDL, and the release of additional proinflammatory molecules [179,180][25][26]. In this chronic inflammation, HDL-proinflammatory may accelerate immune responses toward pathogens, due to HDL remodeling. It is well known that immunological changes occurring in pregnancy for improved fetal tolerance lead to an increased susceptibility to infections. In the acute phase response, HDL levels decrease constantly, with an increase in SAA and ceruloplasmin concentration and a respective decrease in PON1 and Apo A1. Consequently, this could be one of the major risk factors during pregnancy (in trimesters of increased inflammatory processes) for the development of diseases or negative conditions.
Furthermore, an important role of HDL in pregnancy has been reported in the reducing of OS levels, both at placental level and in umbilical cord blood, which are mainly associated with PON1 activity [181][27]. An important factor in pregnancy is the higher activity of lipoprotein-associated phospholipase A2 (LpPla2) (mainly LDL and HDL), which is an enzyme synthesized predominantly by macrophages and associated with inflammatory processes and higher triglyceride levels, as well as in conditions of elevated OS such as GDM concentration of LpPla2, which is highly elevated compared to healthy women. However, this enzyme can be associated with HDL because it improves the antioxidant and anti-inflammatory functions of HDL, thereby reducing OS levels in plasma [182][28].
Another important complication of pregnancy caused by the increase in OS is preeclampsia, which affects both pregnant women and newborns and presents oxidative alterations in both LDL and HDL, caused by lipoperoxidation and inactivation of PON1, potentially leading to improper placentation [183][29].
It has been shown that a maternal diet rich in saturated and trans fatty acids causes harmful changes in the bacteria that colonize the intestine of the offspring, and these in turn produce metabolites that can subsequently affect different organs. Organic acids produced by intestinal bacteria may be involved in inflammatory mechanisms and play a key role in changes in the metabolism and develop neonatally or in adulthood. A high-fat diet during perinatal life predisposes greater expression of the NF-κB gene, which is a transcription factor of multiple biological processes, including immune and inflammatory responses and cell growth and survival [184,185][30][31].
Several studies have suggested that HDL dysfunction is a common pathological factor that connects the metabolic syndrome to NAFLD and cardiovascular disease development. The composition and structure of HDL particles seem to be characterized by the depletion of polyunsaturated fatty acid phospholipids and enrichment of saturated fatty acid ceramides [151,186][32][33]. In this context, preclinical studies have provided mechanistic insights as to how PUFA (especially essential fatty acids, EFA) deficiency promotes hepatic steatosis. EFA can negatively modulate the hepatic de novo lipogenesis machinery toward the negative modulation of the liver X receptor (LXR) of SREBP-1 and/or of the carbohydrate response element binding protein (ChREBP). Also, PUFA can activate the peroxisome proliferator, activated receptor-alpha (PPARα), and may promote fatty acid oxidation [186][33]. This is strong evidence for the role of PUFAs in modulating hepatic lipid metabolism [151][32].
5. Bioactive Compounds and HDL Functionality
Bioactive compounds have been widely studied as mediators of inflammation and OS in several conditions and diseases. Nevertheless, their mechanism of action remains unclear. Some studies describe that they are an important part of the secretion of inflammatory molecules (cytokines, adipokines, etc.), of the mediation of metabolic pathways, or of the regulation of gene expression at the muscle or adipose tissue level [187][34]. Currently, the most commonly studied bioactive compounds are folates [188][35], polyphenolic compounds [189][36], polyunsaturated fatty acids [190][37], prebiotics [191][38], and probiotics [192][39], along with their derivatives.
Studies have described that part of the functionality of HDL is linked to its chemical composition. This, in turn, depends on the appearance of some diseases causing HDL dysfunction [193][40], especially chronic degenerative diseases, which may also change HDL size as well as the number of circulating particles. In DM and coronary heart disease patients, it has been reported that, in addition to Apo A concentration modifications, the presence of OX increases glycoxidation and peroxidation of protein and lipid fractions of HDL, respectively [194,195,196,197][41][42][43][44].
An alternative way to reverse the mentioned effects is via bioactive compounds from functional foods. It is well documented that foods, for example, fruits and vegetables, fish, legumes, cereals, red wine, and elements of the Mediterranean diet, increase the concentration of HDL, TRC, and antioxidant activity at the same time, increasing the activity and/or expression of paraoxonase-1 (PON1), an atheroprotective enzyme that is bound to HDL [198,199,200][45][46][47]. Likewise, it has been reported that foods rich in polyphenols, hydrolysable tannins, and polyunsaturated fatty acids (PUFAs) modify the protein [201][48] c-HDL, Tg-HDL, and Phos-HDL content of HDL [197,198][44][45].
Alternative dietary modifications such as Mediterranean diet [202][49] seek to enhance HDL functionality via regulation of RCT. On the other hand, olive oil consumption [203][50] seems to have a similar effect but is attributable to its effect on the size and stability of HDL. For instance, a study based on red yeast rice extract and additional compounds shown to reduce cardiovascular risk in humans, a significant decrease in the lipid profile associated with cardiovascular risk, mainly c-LDL, and an increase in Apo A1 were found in 102 participants. However, there was no significant difference in the levels of c-HDL [204][51]. In contrast, a pilot study of 167 patients with metabolic syndrome features, bioactive compounds such as docosahexaenoic acid, β-glucans, and anthocyanins were proved as components of fortified functional foods, and a significant decrease in triglycerides and an increase in LDL-C were observed [205][52]. Moreover, in a study that evaluated the structure and function of HDL in adults with overweight, obesity, and cardiovascular risk, it was observed that there is a relationship between the decrease in inflammation markers such as IL-6 with sphingosine 1 phosphate (SP1) of HDL under a diet based on a Mediterranean diet [206][53].
Studies by a research group have demonstrated this: in an animal model as well as in women with acute coronary ischemic syndrome (ACS), by using a microencapsulated product enriched in antioxidants and PUFAs, mainly punicic acid, it was observed that this treatment improved the lipid profile, PON1 activity, and endothelial function mediated by HDL. In ACS women, the dysfunctionality of HDL was reverted by regulating the protein and lipid composition of the smallest subclasses (HDL3) [201,207][48][54]. Two possible explanations for these results suggest that bioactive compounds present in this microencapsulated product could remodel alterations in HDL under conditions of dyslipidemia, OS, and inflammation. Also, another possible explanation is that HDL acts as a vector for bioactive compounds, enhancing its bioavailability and potentially increasing its health benefits.
References
- Li, J.; Xu, Y.; Sun, Z.; Cai, Y.; Wang, B.; Zhang, M.; Ban, Y.; Hou, X.; Hao, Y.; Ouyang, Q.; et al. Differential Lipids in Pregnant Women with Subclinical Hypothyroidism and Their Correlation to the Pregnancy Outcomes. Sci. Rep. 2021, 11, 19689.
- Charlton, F.; Tooher, J.; Rye, K.A.; Hennessy, A. Cardiovascular Risk, Lipids and Pregnancy: Preeclampsia and the Risk of Later Life Cardiovascular Disease. Hear. Lung Circ. 2014, 23, 203–212.
- Adank, M.C.; Johansen, A.K.; Benschop, L.; Van Streun, S.P.; Smak Gregoor, A.M.; Øyri, L.K.L.; Mulder, M.T.; Steegers, E.A.P.; Holven, K.B.; Roeters van Lennep, J.E. Maternal Lipid Levels in Early Pregnancy as a Predictor of Childhood Lipid Levels: A Prospective Cohort Study. BMC Pregnancy Childbirth 2022, 22, 588.
- Estrada-Luna, D.; Ortiz-Rodriguez, M.A.; Medina-Briseño, L.; Carreón-Torres, E.; Izquierdo-Vega, J.A.; Sharma, A.; Cancino-Díaz, J.C.; Pérez-Méndez, O.; Belefant-Miller, H.; Betanzos-Cabrera, G. Current Therapies Focused on High-Density Lipoproteins Associated with Cardiovascular Disease. Molecules 2018, 23, 2730.
- Bausserman, L.L.; Herbert, P.N.; McAdam, K.P. Heterogeneity of Human Serum Amyloid A Proteins. J. Exp. Med. 1980, 52, 641–656.
- Davidson, W.S.; Shah, A.S.; Sexmith, H.; Gordon, S.M. The HDL Proteome Watch: Compilation of Studies Leads to New Insights on HDL Function. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2022, 1867, 159072.
- Shah, A.S.; Tan, L.; Long, J.L.; Davidson, W.S. Proteomic Diversity of High Density Lipoproteins: Our Emerging Understanding of Its Importance in Lipid Transport and Beyond. J. Lipid Res. 2013, 54, 2575–2585.
- Ouimet, M.; Barrett, T.J.; Fisher, E.A. HDL and Reverse Cholesterol Transport: Basic Mechanisms and Their Roles in Vascular Health and Disease. Circ. Res. 2019, 124, 1505–1518.
- Cuchel, M.; Rader, D.J. Macrophage Reverse Cholesterol Transport: Key to the Regression of Atherosclerosis? Circulation 2006, 113, 2548–2555.
- Chen, C.; Khismatullin, D.B. Oxidized Low-Density Lipoprotein Contributes to Atherogenesis via Co-Activation of Macrophages and Mast Cells. PLoS ONE 2015, 10, e0123088.
- Rysz, J.; Gluba-Brzózka, A.; Rysz-Górzyńska, M.; Franczyk, B. The Role and Function of HDL in Patients with Chronic Kidney Disease and the Risk of Cardiovascular Disease. Int. J. Mol. Sci. 2020, 21, 601.
- Buendía-Abad, M.; García-Palencia, P.; de Pablos, L.M.; Alunda, J.M.; Osuna, A.; Martín-Hernández, R.; Higes, M. First Description of Lotmaria Passim and Crithidia Mellificae Haptomonad Stages in the Honeybee Hindgut. Int. J. Parasitol. 2022, 52, 65–75.
- Stadler, J.T.; Scharnagl, H.; Wadsack, C.; Marsche, G. Preeclampsia Affects Lipid Metabolism and HDL Function in Mothers and Their Offspring. Antioxidants 2023, 12, 795.
- Eren, E. High Density Lipoprotein and It’s Dysfunction. Open Biochem. J. 2012, 6, 78–93.
- Gordon, T.; Castelli, W.P.; Hjortland, M.C.; Kannel, W.B.; Dawber, T.R. High Density Lipoprotein as a Protective Factor against Coronary Heart Disease. The Framingham Study. Am. J. Med. 1977, 62, 707–714.
- Patanapirunhakit, P.; Karlsson, H.; Mulder, M.; Ljunggren, S.; Graham, D.; Freeman, D. Sphingolipids in HDL—Potential Markers for Adaptation to Pregnancy? Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2021, 1866, 158955.
- Luna-Luna, M.; Niesor, E.; Pérez-Méndez, Ó. HDL as Bidirectional Lipid Vectors: Time for New Paradigms. Biomedicines 2022, 10, 1180.
- Chen, R.; Chen, Q.; Zheng, J.; Zeng, Z.; Chen, M.; Li, L.; Zhang, S. Serum Amyloid Protein A in Inflammatory Bowel Disease: From Bench to Bedside. Cell Death Discov. 2023, 9, 154.
- Streese, L.; Habisch, H.; Deiseroth, A.; Carrard, J.; Infanger, D.; Schmidt-Trucksäss, A.; Madl, T.; Hanssen, H. Lipoprotein Subclasses Independently Contribute to Subclinical Variance of Microvascular and Macrovascular Health. Molecules 2022, 27, 4760.
- Woudberg, N.J.; Goedecke, J.H.; Blackhurst, D.; Frias, M.; James, R.; Opie, L.H.; Lecour, S. Association between Ethnicity and Obesity with High-Density Lipoprotein (HDL) Function and Subclass Distribution. Lipids Health Dis. 2016, 15, 92.
- Muñoz-Vega, M.; Massó, F.; Páez, A.; Vargas-Alarcón, G.; Coral-Vázquez, R.; Mas-Oliva, J.; Carreón-Torres, E.; Pérez-Méndez, Ó. Hdl-Mediated Lipid Influx to Endothelial Cells Contributes to Regulating Intercellular Adhesion Molecule (Icam)-1 Expression and Enos Phosphorylation. Int. J. Mol. Sci. 2018, 19, 3394.
- Brites, F.; Martin, M.; Guillas, I.; Kontush, A. Antioxidative Activity of High-Density Lipoprotein (HDL): Mechanistic Insights into Potential Clinical Benefit. BBA Clin. 2017, 8, 66–77.
- Jayaraman, S.; Haupt, C.; Gursky, O. Thermal Transitions in Serum Amyloid A in Solution and on the Lipid: Implications for Structure and Stability of Acute-Phase HDL. J. Lipid Res. 2015, 56, 1531–1542.
- Göçmen, A.Y.; Şahin, E.; Semiz, E.; Gümüslü, S. Is Elevated Serum Ceruloplasmin Level Associated with Increased Risk of Coronary Artery Disease? Can. J. Cardiol. 2008, 24, 209–212.
- Khovidhunkit, W.; Kim, M.S.; Memon, R.A.; Shigenaga, J.K.; Moser, A.H.; Feingold, K.R.; Grunfeld, C. Effects of Infection and Inflammation on Lipid and Lipoprotein Metabolism: Mechanisms and Consequences to the Host. J. Lipid Res. 2004, 45, 1169–1196.
- Rosenson, R.S.; Brewer, H.B., Jr.; Ansell, B.J.; Barter, P.; Chapman, M.J.; Heinecke, J.W.; Kontush, A.; Tall, A.R.; Webb, N.R. Dysfunctional HDL and Atherosclerotic Cardiovascular Disease. Nat. Rev. Cardiol. 2016, 13, 48–60.
- Zafaranieh, S.; Stadler, J.T.; Pammer, A.; Marsche, G.; van Poppel, M.N.M.; Desoye, G. The Association of Physical Activity and Sedentary Behavior with Maternal and Cord Blood Anti-Oxidative Capacity and HDL Functionality: Findings of DALI Study. Antioxidants 2023, 12, 827.
- Schliefsteiner, C.; Hirschmugl, B.; Kopp, S.; Curcic, S.; Bernhart, E.M.; Marsche, G.; Lang, U.; Desoye, G.; Wadsack, C. Maternal Gestational Diabetes Mellitus Increases Placental and Foetal Lipoprotein-Associated Phospholipase A2 Which Might Exert Protective Functions against Oxidative Stress. Sci. Rep. 2017, 7, 12628.
- León-Reyes, G.; Espino, Y.; Sosa, S.; Medina-Navarro, R.; Guzmán-Grenfell, A.M.; Medina-Urrutia, A.X.; Fuentes-García, S.; Hicks, G.J.J.; Torres-Ramos, Y.D. Oxidative Modifications of Foetal LDL-c and HDL-c Lipoproteins in Preeclampsia. Lipids Health Dis. 2018, 17, 110.
- Jayalekshmi, V.S.; Jaikumar, V.S.; Mehra, P.; Thulaseedharan, T.; Vinod, V.M.; Ramachandran, S. Differential Expression of Lipid Metabolic Genes in Hypercholesterolemic Rabbit Placenta Predisposes the Offspring to Develop Atherosclerosis in Early Adulthood. Life Sci. 2023, 327, 121823.
- Rodríguez-Hernández, H.; Simental-Mendía, L.E. The Triglycerides and Glucose Index Is Highly Associated with Non-Alcoholic Fatty Liver Disease in Overweight and Obese Women. Ir. J. Med. Sci. 2023, 2023, 11845.
- Zhou, Y.; Peng, H.; Xu, H.; Li, J.; Golovko, M.; Cheng, H.; Lynch, E.C.; Liu, L.; McCauley, N.; Kennedy, L.; et al. Maternal Diet Intervention before Pregnancy Primes Offspring Lipid Metabolism in Liver. Lab. Investig. 2020, 100, 553–569.
- Mocciaro, G.; Allison, M.; Jenkins, B.; Azzu, V.; Huang-Doran, I.; Herrera-Marcos, L.V.; Hall, Z.; Murgia, A.; Susan, D.; Frontini, M.; et al. Non-Alcoholic Fatty Liver Disease Is Characterised by a Reduced Polyunsaturated Fatty Acid Transport via Free Fatty Acids and High-Density Lipoproteins (HDL). Mol. Metab. 2023, 73, 101728.
- Pelczyńska, M.; Moszak, M.; Wesołek, A.; Bogdański, P. The Preventive Mechanisms of Bioactive Food Compounds against Obesity-Induced Inflammation. Antioxidants 2023, 12, 1232.
- Pathikkal, A.; Puthusseri, B.; Divya, P.; Rudrappa, S.; Chauhan, V.S. Folate Derivatives, 5-Methyltetrahydrofolate and 10-Formyltetrahydrofolate, Protect BEAS-2B Cells from High Glucose–Induced Oxidative Stress and Inflammation. Vitr. Cell. Dev. Biol.—Anim. 2022, 58, 419–428.
- Zhou, K.; Raffoul, J.J. Potential Anticancer Properties of Grape Antioxidants. J. Oncol. 2012, 2012, 803294.
- Delmastro-Greenwood, M.; Freeman, B.A.; Wendell, S.G. Redox-Dependent Anti-Inflammatory Signaling Actions of Unsaturated Fatty Acids. Annu. Rev. Physiol. 2014, 76, 79–105.
- Gong, S.; Ji, X.; Su, J.; Wang, Y.; Yan, X.; Wang, G.; Xiao, B.; Dong, H.; Xiang, X.; Liu, S. Yeast Fermentate Prebiotic Ameliorates Allergic Asthma, Associating with Inhibiting Inflammation and Reducing Oxidative Stress Level through Suppressing Autophagy. Mediat. Inflamm. 2021, 2021, 4080935.
- Dai, Y.; Quan, J.; Xiong, L.; Luo, Y.; Yi, B. Probiotics Improve Renal Function, Glucose, Lipids, Inflammation and Oxidative Stress in Diabetic Kidney Disease: A Systematic Review and Meta-Analysis. Ren. Fail. 2022, 44, 862–880.
- Kelesidis, T.; Jackson, N.; Mccomsey, G.A.; Wang, X.; Elashoff, D.; Dube, M.P.; Brown, T.T.; Yang, O.O.; Stein, J.H.; Currier, J.S. Oxidized Lipoproteins Are Associated with Markers of Inflammation and Immune Activation in HIV-1 Infection. AIDS 2016, 30, 2625–2633.
- Vergès, B. Dyslipidemia in Type 1 Diabetes: A Masked Danger. Trends Endocrinol. Metab. 2020, 31, 422–434.
- Ganjali, S.; Dallinga-Thie, G.M.; Simental-Mendía, L.E.; Banach, M.; Pirro, M.; Sahebkar, A. HDL Functionality in Type 1 Diabetes. Atherosclerosis 2017, 267, 99–109.
- Stadler, J.T.; Marsche, G. Dietary Strategies to Improve Cardiovascular Health: Focus on Increasing High-Density Lipoprotein Functionality. Front. Nutr. 2021, 8, 1–16.
- Cardner, M.; Yalcinkaya, M.; Goetze, S.; Luca, E.; Balaz, M.; Hunjadi, M.; Hartung, J.; Shemet, A.; Kränkel, N.; Radosavljevic, S.; et al. Structure-Function Relationships of HDL in Diabetes and Coronary Heart Disease. JCI Insight 2020, 5, 1–18.
- Luna-Castillo, K.P.; Lin, S.; Muñoz-Valle, J.F.; Vizmanos, B.; López-Quintero, A.; Márquez-Sandoval, F. Functional Food and Bioactive Compounds on the Modulation of the Functionality of Hdl-c: A Narrative Review. Nutrients 2021, 13, 1165.
- Vaisar, T.; Kanter, J.E.; Wimberger, J.; Irwin, A.D.; Gauthier, J.; Wolfson, E.; Bahnam, V.; Wu, I.H.; Shah, H.; Keenan, H.A.; et al. High Concentration of Medium-Sized HDL Particles and Enrichment in HDL Paraoxonase 1 Associate with Protection from Vascular Complications in People with Long-Standing Type 1 Diabetes. Diabetes Care 2020, 43, 178–186.
- Gu, X.; Huang, Y.; Levison, B.S.; Gerstenecker, G.; DiDonato, A.J.; Hazen, L.B.; Lee, J.; Gogonea, V.; DiDonato, J.A.; Hazen, S.L. Identification of Critical Paraoxonase 1 Residues Involved in High Density Lipoprotein Interaction. J. Biol. Chem. 2016, 291, 1890–1904.
- Dorantes-Morales, A.; Estrada-Luna, D.; Bautista-Pérez, R.; Betanzos-Cabrera, G.; Luna-Luna, M.; Flores-Castillo, C.; Vargas-Alarcón, G.; Fragoso, J.M.; Pérez-Méndez, Ó.; Carreón-Torres, E.; et al. Microencapsulated Pomegranate Modifies the Composition and Function of High-Density. Molecules 2020, 25, 3297.
- Hernáez, Á.; Castañer, O.; Elosua, R.; Pintó, X.; Estruch, R.; Salas-Salvadó, J.; Corella, D.; Arós, F.; Serra-Majem, L.; Fiol, M.; et al. Mediterranean Diet Improves High-Density Lipoprotein Function in High-Cardiovascular-Risk Individuals. Circulation 2017, 135, 633–643.
- Hernáez, Á.; Fernández-Castillejo, S.; Farràs, M.; Catalán, Ú.; Subirana, I.; Montes, R.; Solà, R.; Muñoz-Aguayo, D.; Gelabert-Gorgues, A.; Díaz-Gil, Ó.; et al. Olive Oil Polyphenols Enhance High-Density Lipoprotein Function in Humans: A Randomized Controlled Trial. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2115–2119.
- Sola, R.; Valls, R.M.; Puzo, J.; Calabuig, J.R.; Brea, A.; Pedret, A.; Morinã, D.; Villar, J.; Millán, J.; Anguera, A. Effects of Poly-Bioactive Compounds on Lipid Profile and Body Weight in a Moderately Hypercholesterolemic Population with Low Cardiovascular Disease Risk: A Multicenter Randomized Trial. PLoS ONE 2014, 9, e101978.
- Bub, A.; Malpuech-Brugère, C.; Orfila, C.; Amat, J.; Arianna, A.; Blot, A.; Di Nunzio, M.; Holmes, M.; Kertész, Z.; Marshall, L.; et al. A Dietary Intervention of Bioactive Enriched Foods Aimed at Adults at Risk of Metabolic Syndrome: Protocol and Results from PATHWAY-27 Pilot Study. Nutrients 2019, 11, 1814.
- Muralidharan, J.; Papandreou, C.; Soria-Florido, M.T.; Sala-Vila, A.; Blanchart, G.; Estruch, R.; Martínez-González, M.A.; Corella, D.; Ros, E.; Ruiz-Canela, M.; et al. Cross-Sectional Associations between HDL Structure or Function, Cell Membrane Fatty Acid Composition, and Inflammation in Elderly Adults. J. Nutr. 2022, 152, 789–795.
- Estrada-Luna, D.; Carreón-Torres, E.; Bautista-Perez, R.; Betanzos-Cabrera, G.; Dorantes-Morales, A.; Luna-Luna, M.; Vargas-Barrón, J.; Mejía, A.M.; Fragoso, J.M.; Carvajal-Aguilera, K.; et al. Microencapsulated Pomegranate Reverts High-Density Lipoprotein (HDL)-Induced Endothelial Dysfunction and Reduces Postprandial Triglyceridemia in Women with Acute Coronary Syndrome. Nutrients 2019, 11, 1710.
More