The intracellular concentrations of oxygen and reactive oxygen species (ROS) in living cells represent critical information for investigating physiological and pathological conditions. Real-time measurement often relies on genetically encoded proteins that are responsive to fluctuations in either oxygen or ROS concentrations. The direct binding or chemical reactions that occur in their presence either directly alter the fluorescence properties of the binding protein or alter the fluorescence properties of fusion partners, mostly consisting of variants of the green fluorescent protein. Oxygen sensing takes advantage of several mechanisms, including (i) the oxygen-dependent hydroxylation of a domain of the hypoxia-inducible factor-1, which, in turn, promotes its cellular degradation along with fluorescent fusion partners; (ii) the naturally oxygen-dependent maturation of the fluorophore of green fluorescent protein variants; and (iii) direct oxygen binding by proteins, including heme proteins, expressed in fusion with fluorescent partners, resulting in changes in fluorescence due to conformational alterations or fluorescence resonance energy transfer.
- oxygen sensor
- reactive oxygen species
- fluorescent proteins
1. Relevance of O2 and ROS Sensing
2. Biosensors for O2
2.1. O
2
Sensing Mediated by Prolyl Hydroxylases
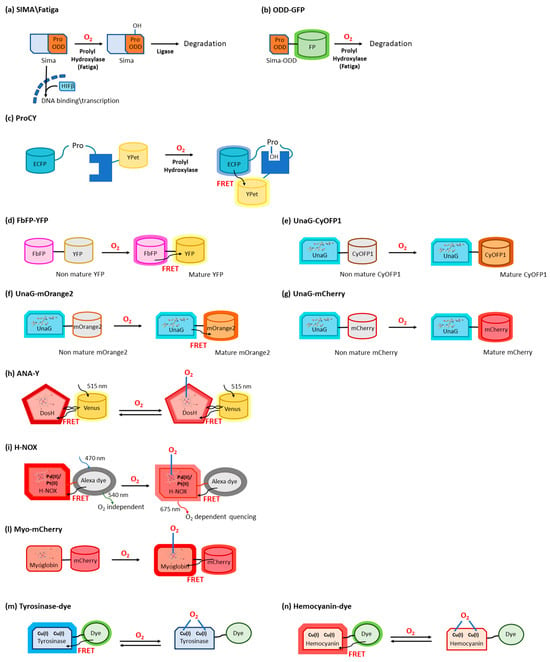
The transcription factor hypoxia-inducible factor-1 (HIF-1) is the main actor in modulating O2-mediated gene expression levels in all Parahoxozoa. HIF-1 is a αβ heterodimer with the levels of the β subunit being O2-independent, whereas those of the α subunit are O2-dependent [10][3] through its O2-dependent proteolysis [11][4]. In fact, the α subunit is constitutively synthesized but rapidly degraded under normoxic conditions [12][5] as a consequence of enzymatic hydroxylation of conserved proline residues (Pro402 and Pro564) located in its oxygen-dependent degradation domain (ODD) [13,14][6][7]. The enzymes responsible for proteolysis are 2-oxoglutarate-dependent dioxygenases containing a prolyl hydroxylase domain (PHD) [15][8]. Hydroxylated HIF-1α binds to the von Hippel–Lindau tumor suppressor protein (VHL) and is then ubiquitylated by the VHL-E3 ligase complex (Figure 1a).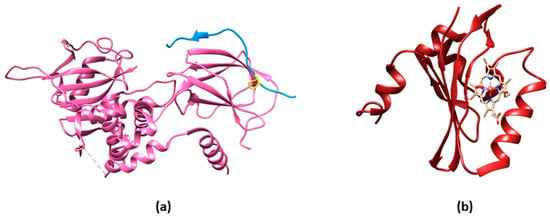
2.2. O
2
Sensors Based on O
2
-Dependent Maturation of GFP Variants
2.3. O
2
Sensors Based on O
2
-Binding Heme Proteins
Oxygen sensing has been also approached by exploiting heme-binding proteins, which use the iron-containing heme group as the O2 sensing element to be transduced in other cellular signals. These constructs are commonly composed of an N-terminal heme-containing globin domain, whose conformational variation upon O2 binding activates a functional domain that triggers a catalytic domain. This latter can have a diguanylate cyclase (DGC) [37][26] or phosphodiesterase (PDE) [38][27] activity toward cyclic diGMP (c-diGMP) and histidine kinase (HK) activity [39][28]. An example is a genetically encoded O2 biosensor based on the direct O2 sensor DosP from E. coli. This protein contains a heme-binding globin domain called DosH (Figure 1b) and a PDE catalytic domain, which converts cyclic-di-GMP to linear-di-GMP [40][29]. However, based on the crystallographic structures [18[10][30],41], the conformational variation of DosP upon O2 binding or dissociation is not sufficient to develop a suitable FRET signal; to overcome this limitation, DosH has been associated with a fluorescent protein to exploit the heme spectroscopic changes in Soret and Q peaks upon O2 binding shifting from 425 and 560 nm to 414 and 580 nm, respectively [42][31]. The yellow FP Venus—a GFP variant with a suitable spectral overlap with DosH—was conjugated with DosH using an optimized antiparallel coiled-coil linker. When Venus was excited at 500 nm, its fluorescence emission at 527 nm was absorbed by DosH in the O2-free form, resulting in low Venus quantum yield; when DosH was O2-bound, Venus’s fluorescence intensity increased. The change in heme absorption was therefore amplified by a change in fluorescence intensity. This sensor, called ANA-Y (anaerobic/aerobic sensor yellow), was used to sense O2 in the micromolar range (Figure 2h) [43][32]. The heme nitric oxide/oxygen-binding protein (H-NOX) of the thermophilic bacterium Caldanaerobacter subterraneus has been exploited to develop an O2 sensor as a robust protein scaffold also able to bind unnatural heme cofactors. The natural heme was replaced with a Pd(II) or Pt(II) porphyrin, a phosphorescent cofactor (650–800 nm range) that, in the triplet excited state, can interact with molecular oxygen; in particular, Pd(II) porphyrins are more sensitive to low O2 levels and Pd(II) has a larger range of O2 sensitivity. A ratiometric sensor based on H-NOX incorporating Pd(II) or Pt(II) porphyrins was developed by conjugating to the protein an Alexa fluorescent dye that is the O2-independent FRET donor to the porphyrin. The selective excitation of Alexa dye guarantees that the porphyrin emission will only be from FRET, thus minimizing the background signal (Figure 2i) [44][33]. As a different approach, the spectroscopic changes of heme upon O2 binding have been exploited and amplified in a fusion protein (Myo-mCherry), where myoglobin has been fused with mCherry fluorescent protein; spectral changes resulted in a change of FRET, captured as a change in fluorescence lifetime within cells by FLIM (Figure 2l).2.4. O
2
Sensors Based on O
2
-Binding Copper Proteins
The use of type 3 copper proteins to sense O2 has been also approached; these proteins have a binding site with two Cu(I) ions that specifically bind to O2, and the complex, once formed, gives absorption peaks at 340 and 570 nm [47][34]. O2 binding typically also causes Trp fluorescence changes, but the use of fluorescence coming from an exogenous fluorophore with a more efficient and higher-wavelength emission is made necessary to increase sensitivity (Trp has a moderate quantum yield, around 0.14) and to allow the detection in tissues showing significant autofluorescence. An example is given by the tyrosinase from the bacterium Streptomyces antibioticus and hemocyanin from the arthropod Carcinus aestuarii, which have been conjugated at the N-terminus with different fluorescent dyes (Alexa 350, Atto390, Cy3, Cy5, Atto655), showing an absorption overlapping Trp emission, thus allowing the detection as a variation in FRET upon O2 binding (Figure 2m) [48][35]. The detection of a single O2 molecule has been obtained by exploiting the O2 carrier hemocyanin from the tarantula Eurypelma californicum. This protein shows spectral differences when it binds oxygen, and these absorption changes can be used to sense O2 presence. Moreover, to amplify the signal, TAMRA fluorescent dye was conjugated with hemocyanin, and its fluorescence quantum yield was shown to decrease by a factor of two when hemocyanin was oxygenated [49][36]. This was caused by a FRET where TAMRA was the donor and the O2 binding site was the acceptor (Figure 2n) [50][37].3. Biosensors for ROS
3.1. ROS Sensors Based on roFPs
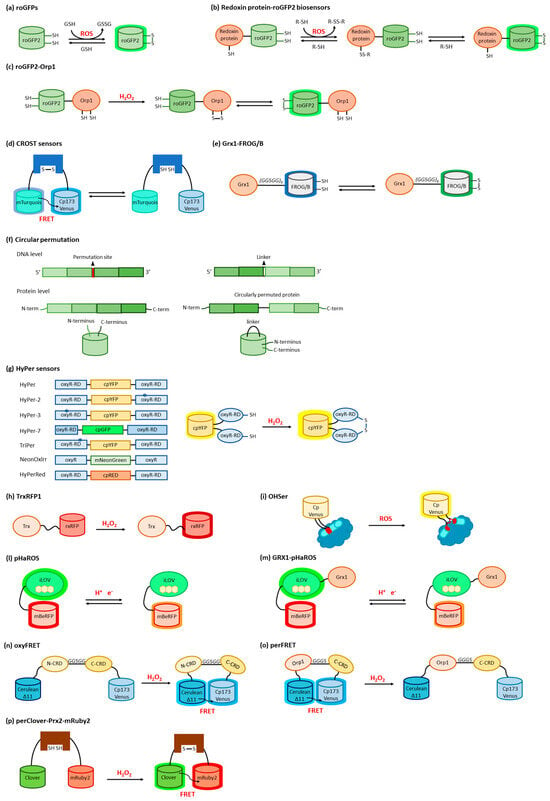
Reduction-oxidation sensitive GFPs (roGFPs) are non-natural variants of A. victoria GFP obtained by substituting surface-exposed residues with cysteine residues appropriately distanced to form disulfide bonds at suitable redox potentials (Figure 3a,b and Figure 4a). The first examples of roFPs consisted of the introduction of an artificial pair of cysteine residues on YFP, giving rise to one of the first classes of redox-sensitive protein-based fluorescent sensors. The first rxYFP was conceived in 2001 by Winther’s research group, when the YFP sequence was modified to mutate Asn149 and Ser202 to two cysteine residues able to form a disulfide bridge under oxidizing conditions (Figure 3a) [51][38].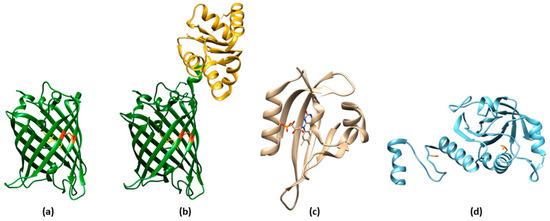
3.2. ROS Sensors Based on roGFP Fusion Proteins
3.3. ROS Sensors Based on Circularly Permuted FPs
Circular permutation of proteins, a naturally occurring event first described in lectins [100][58], consists of the modification of their primary structure by fusing the natural N- and C-termini with peptide linkers and creating new extremities, generally on exposed flexible loops (Figure 4f). Based on this approach, it is possible to insert ROS-sensitive domains inside GFP or other FP sequences, producing a chimeric structure. The native folding is easily preserved but, at the same time, it is possible to modulate FP intrinsic fluorescence through conformational rearrangements promoted by ligand binding to the receptor domain [101,102][59][60]. HyPer sensors were developed, starting in 2006, by Belousov and co-workers, by exploiting circularly permuted FPs (Figure 4g) [103,104,105,106][61][62][63][64]. A circularly permuted YFP (cpYFP) was modified to develop a chimeric construct with the regulatory domain (RD, residues 80–310) of the OxyR transcription factor from E. coli, specifically sensitive to H2O2 [103][61]. Under oxidizing conditions, OxyR switched to the DNA binding-competent form with Cys199 and Cys208 engaging in a disulfide bond. The cpYFP sequence was inserted between residues 205 and 206 of OxyR-RD by exploiting two short linkers, and the chimeric structure assembled as weak dimers [105][63]. This reaction required the formation of sulfenic acid derivatives between H2O2 and Cys199; Cys199 was then repelled by the hydrophobic pocket and approached Cys208 to bridge [107][65]. In the following years, an advanced version of HyPer was developed by the same research group to improve the biosensor dynamic range. The A406V mutation, corresponding to an amino acid substitution on the OxyR-RD moiety (A233V on the OxyR wild type sequence), was fortuitously discovered as improving two-fold the HyPer dynamic range; the new probe was called HyPer-2 [105][63]. rxRFP features have been exploited also by Ai’s research group to develop the first genetically encoded biosensor for Trx redox system monitoring [114][66]. Specifically, the optimal coupling of the redox cysteine pairs of human Trx1 and rxRFP was obtained by inserting a Gly-Ser-rich linker between the two proteins; several mutation rounds led to the set-up of TrxRFP1 (Figure 4h), which showed a fluorescence fingerprint identical to rxRFP1 alone, presenting excitation and emission maxima at 576 nm and 600 nm, respectively, with a 5.7-fold fluorescence increase in dynamic range. The result was the development of a fluorescent probe highly responsive to Trx peroxidase-mediated H2O2 oxidation, even at a nanomolar concentration; its midpoint redox potential of −281 mV makes this sensor suitable for cytoplasmic measurements in live cells where, also, TrxRFP1 is not directly oxidized by H2O2 and is not influenced by the presence of glutathione, a characteristic that consents the specific monitoring of the cellular Trx redox system [114,115][66][67]. The transcriptional regulator OhrR from Xanthomonas campestris can sense and respond to organic hydroperoxides via the oxidation of a cysteine residue, which induces a large conformational rearrangement predisposing the formation of an intermolecular disulfide bridge. Chen’s group exploited the behavior of OhrR to develop an organic hydroperoxide sensor by inserting a circularly permuted version of the fluorescent Venus protein (cpVenus) between OhrR residues 119 and 120 (Figure 4i) [117][68]. The probe, named OHSer (organic hydroperoxide sensor), showed a fluorescence emission at 526 nm upon excitation at 519 nm, a spectral property that adapted well to the reduction of biological damage during in vivo measurements.3.4. ROS Sensors Based on LOV Domains
The light–oxygen–voltage-sensing domains (LOV domains) [118][69] are part of the PAS domain superfamily [119][70] (named after three proteins in which it occurs, i.e., Per, Arnt, and Sim) and are present in proteins from higher plants, microalgae, fungi, and bacteria. LOV-containing proteins are involved in the detection and adaptation to environmental changes. Particularly, LOV domains are involved in controlling phototropism, chloroplast movements, and stomatal opening in higher plants [120][71]. In fungi, they are involved in the circadian temporal organization of the cells. LOV domains consist of 110 to 140 amino acids organized in 5 antiparallel β-sheets and several α-helices and contain a blue-light sensitive flavin chromophore, usually flavin mononucleotide (FMN) (Figure 3c) [121][72]. The LOV domain of LOV-containing proteins serves as a photoswitch, activating kinase, phosphodiesterase, and DNA-binding domains upon light absorption [122][73]. Other than being used as fluorescent reporters, engineered LOV domains have been used to devise sensors, ranging from metals [124][74] to O2 [30][21]. LOV-based chimeras have also been used to develop sensors for the intracellular redox state. Among LOV-based sensors, a dual-function pH and redox-sensitive FP named pHaROS has been devised as a sensor for both H2O2 and pH in living cells. It consists of the iLOV domain fused with mBeRFP, a variant of the monomeric far-red FP mKATE (Figure 4l) [125,126][75][76]. The iLOV portion of pHaROS can reversibly gain an electron and displays fluorescence intensity changes depending on the redox state. GRX1-pHaROS is a fusion protein of pHaROS and Grx1, which confers higher redox specificity [125][75], in a manner similar to Grx1-roGFP2 (Figure 4m) [84,85][44][45].3.5. ROS Sensors Based on YAP1
YAP1 is a transcriptional regulator that activates the transcription of genes involved in oxidative stress response and redox homeostasis in S. cerevisiae. YAP1, which is partially disordered, can undergo conformational changes affecting disulfide bond formation at two cysteine-rich domains (CRDs) that mask the nuclear export signal, thus preventing its nuclear export. In S. cerevisiae Yap1 and Orp1 constitute a redox relay system, where Yap1 is oxidized by the peroxidase Orp1 in the presence of H2O2. Two genetically encoded sensors for H2O2 based on Yap1 were reported, based on the FRET couple Cerulean Δ11 and Cp173 Venus separated by the two CRDs of Yap1 (OxyFRET, Figure 4n) [127][77] or by Orp1 linked with the C-terminal CRD of Yap1 (PerFRET, Figure 4o) [127][77].3.6. ROS Sensors Based on Peroxiredoxin
Peroxiredoxins (Prxs) (Figure 3d) belonging to the “AhpC-Prx1” subfamily, such as mammalian Prx1 and Prx2, carry the peroxidatic and resolving cysteines on two different subunits. Upon oxidation in response to H2O2 increases, these Prx undergo a conformational rearrangement followed by dimerization. This mechanism has been used to devise a fusion-protein FRET couple with FPs perClover and mRuby2 separated by human Prx2 (Figure 4p) [130][78]. Upon excitation at 488 nm, the 625/525 nm emission ratio increased in function of H2O2 sensing, without the interference of pH fluctuations. The probe was highly selective toward H2O2, with the only exception of TBHP, which naturally reacted with Prxs. To characterize the response of the cytosolic probe to H2O2, HeLa cells were transfected with the probe-encoding gene, put in contact with an external bolus addition of H2O2, and imaged with a widefield microscope [130][78].References
- Raymond, J.; Segrè, D. The Effect of Oxygen on Biochemical Networks and the Evolution of Complex Life. Science 2006, 311, 1764–1767.
- Della Rocca, Y.; Fonticoli, L.; Rajan, T.S.; Trubiani, O.; Caputi, S.; Diomede, F.; Pizzicannella, J.; Marconi, G.D. Hypoxia: Molecular pathophysiological mechanisms in human diseases. J. Physiol. Biochem. 2022, 78, 739–752.
- Wang, G.L.; Jiang, B.H.; Rue, E.A.; Semenza, G.L. Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS heterodimer regulated by cellular O2 tension. Proc. Natl. Acad. Sci. USA 1995, 92, 5510–5514.
- Görlach, A.; Camenisch, G.; Kvietikova, I.; Vogt, L.; Wenger, R.H.; Gassmann, M. Efficient translation of mouse hypoxia-inducible factor-1α under normoxic and hypoxic conditions. Biochim. Biophys. Acta Gene Struct. Expr. 2000, 1493, 125–134.
- Jewell, U.R.; Kvietikova, I.; Scheid, A.; Bauer, C.; Wenger, R.H.; Gassmann, M. Induction of HIF–1α in response to hypoxia is instantaneous. FASEB J. 2001, 15, 1312–1314.
- Bruick, R.K.; McKnight, S.L. Oxygen Sensing Gets a Second Wind. Science 2002, 295, 807–808.
- Pugh, C.W.; O’Rourke, J.F.; Nagao, M.; Gleadle, J.M.; Ratcliffe, P.J. Activation of hypoxia-inducible factor-1; definition of regulatory domains within the alpha subunit. J. Biol. Chem. 1997, 272, 11205–11214.
- Masson, N.; Ratcliffe, P.J. HIF prolyl and asparaginyl hydroxylases in the biological response to intracellular O2 levels. J. Cell Sci. 2003, 116, 3041–3049.
- Hon, W.-C.; Wilson, M.I.; Harlos, K.; Claridge, T.D.W.; Schofield, C.J.; Pugh, C.W.; Maxwell, P.H.; Ratcliffe, P.J.; Stuart, D.I.; Jones, E.Y. Structural basis for the recognition of hydroxyproline in HIF-1α by pVHL. Nature 2002, 417, 975–978.
- Park, H.; Suquet, C.; Satterlee, J.D.; Kang, C. Insights into Signal Transduction Involving PAS Domain Oxygen-Sensing Heme Proteins from the X-ray Crystal Structure of Escherichia Coli Dos Heme Domain (Ec DosH). Biochemistry 2004, 43, 2738–2746.
- Arquier, N.; Vigne, P.; Duplan, E.; Hsu, T.; Therond, P.P.; Frelin, C.; D’Angelo, G. Analysis of the hypoxia-sensing pathway in Drosophila melanogaster. Biochem. J. 2005, 393, 471–480.
- Lavista-Llanos, S.; Centanin, L.; Irisarri, M.; Russo, D.M.; Gleadle, J.M.; Bocca, S.N.; Muzzopappa, M.; Ratcliffe, P.J.; Wappner, P. Control of the Hypoxic Response in Drosophila melanogaster by the Basic Helix-Loop-Helix PAS Protein Similar. Mol. Cell. Biol. 2002, 22, 6842–6853.
- Bacon, N.C.M.; Wappner, P.; O’Rourke, J.F.; Bartlett, S.M.; Shilo, B.; Pugh, C.W.; Ratcliffe, P.J. Regulation of the Drosophila bHLH-PAS Protein Sima by Hypoxia: Functional Evidence for Homology with Mammalian HIF-1α. Biochem. Biophys. Res. Comm. 1998, 249, 811–816.
- Youssef, S.; Ren, W.; Ai, H.-w. A Genetically Encoded FRET Sensor for Hypoxia and Prolyl Hydroxylases. ACS Chem. Biol. 2016, 11, 2492–2498.
- Remington, S.J. Fluorescent proteins: Maturation, photochemistry and photophysics. Curr. Opin. Struct. Biol. 2006, 16, 714–721.
- Chapman, S.; Faulkner, C.; Kaiserli, E.; Garcia-Mata, C.; Savenkov, E.I.; Roberts, A.G.; Oparka, K.J.; Christie, J.M. The photoreversible fluorescent protein iLOV outperforms GFP as a reporter of plant virus infection. Proc. Natl. Acad. Sci. USA 2008, 105, 20038–20043.
- Drepper, T.; Eggert, T.; Circolone, F.; Heck, A.; Krauß, U.; Guterl, J.-K.; Wendorff, M.; Losi, A.; Gärtner, W.; Jaeger, K.-E. Reporter proteins for in vivo fluorescence without oxygen. Nat. Biotech. 2007, 25, 443–445.
- Mukherjee, A.; Walker, J.; Weyant, K.B.; Schroeder, C.M. Characterization of Flavin-Based Fluorescent Proteins: An Emerging Class of Fluorescent Reporters. PLoS ONE 2013, 8, e64753.
- Delvigne, F.; Boxus, M.; Ingels, S.; Thonart, P. Bioreactor mixing efficiency modulates the activity of a prpoS::GFP reporter gene in E. coli. Microb. Cell Factories 2009, 8, 15.
- Garcia, J.R.; Cha, H.J.; Rao, G.; Marten, M.R.; Bentley, W.E. Microbial nar-GFP cell sensors reveal oxygen limitations in highly agitated and aerated laboratory-scale fermentors. Microb. Cell Factories 2009, 8, 6.
- Potzkei, J.; Kunze, M.; Drepper, T.; Gensch, T.; Jaeger, K.-E.; Büchs, J. Real-time determination of intracellular oxygen in bacteria using a genetically encoded FRET-based biosensor. BMC Biol. 2012, 10, 28.
- Bauer, N.; Maisuls, I.; Pereira da Graça, A.; Reinhardt, D.; Erapaneedi, R.; Kirschnick, N.; Schäfers, M.; Grashoff, C.; Landfester, K.; Vestweber, D.; et al. Genetically encoded dual fluorophore reporters for graded oxygen-sensing in light microscopy. Biosens. Bioelectron. 2023, 221, 114917.
- Panicucci, G.; Iacopino, S.; De Meo, E.; Perata, P.; Weits, D.A. An Improved HRPE-Based Transcriptional Output Reporter to Detect Hypoxia and Anoxia in Plant Tissue. Biosensors 2020, 10, 197.
- Kumagai, A.; Ando, R.; Miyatake, H.; Greimel, P.; Kobayashi, T.; Hirabayashi, Y.; Shimogori, T.; Miyawaki, A. A Bilirubin-Inducible Fluorescent Protein from Eel Muscle. Cell 2013, 153, 1602–1611.
- Bettati, S.; Pasqualetto, E.; Lolli, G.; Campanini, B.; Battistutta, R. Structure and single crystal spectroscopy of Green Fluorescent Proteins. Biochim. Biophys. Acta Proteins Proteom. 2011, 1814, 824–833.
- Sawai, H.; Yoshioka, S.; Uchida, T.; Hyodo, M.; Hayakawa, Y.; Ishimori, K.; Aono, S. Molecular oxygen regulates the enzymatic activity of a heme-containing diguanylate cyclase (HemDGC) for the synthesis of cyclic di-GMP. Biochim. Biophys. Acta Proteins Proteom. 2010, 1804, 166–172.
- Shimizu, T. The Heme-Based Oxygen-Sensor Phosphodiesterase Ec DOS (DosP): Structure-Function Relationships. Biosensors 2013, 3, 211–237.
- Stranava, M.; Martínek, V.; Man, P.; Fojtikova, V.; Kavan, D.; Vaněk, O.; Shimizu, T.; Martinkova, M. Structural characterization of the heme-based oxygen sensor, AfGcHK, its interactions with the cognate response regulator, and their combined mechanism of action in a bacterial two-component signaling system. Proteins Struct. Funct. Bioinform. 2016, 84, 1375–1389.
- Tuckerman, J.R.; Gonzalez, G.; Sousa, E.H.S.; Wan, X.; Saito, J.A.; Alam, M.; Gilles-Gonzalez, M.-A. An Oxygen-Sensing Diguanylate Cyclase and Phosphodiesterase Couple for c-di-GMP Control. Biochemistry 2009, 48, 9764–9774.
- Park, H.; Suquet, C.; Savenkova, M.I.; Satterlee, J.D.; Kang, C. Cloning, purification, crystallization and preliminary X-ray analysis of DOS heme domain, a new heme oxygen sensor in Escherichia coli. Acta Crystallogr. D 2002, 58, 1504–1506.
- Ishitsuka, Y.; Araki, Y.; Tanaka, A.; Igarashi, J.; Ito, O.; Shimizu, T. Arg97 at the Heme-Distal Side of the Isolated Heme-Bound PAS Domain of a Heme-Based Oxygen Sensor from Escherichia coli (Ec DOS) Plays Critical Roles in Autoxidation and Binding to Gases, Particularly O2. Biochemistry 2008, 47, 8874–8884.
- Nomata, J.; Hisabori, T. Development of heme protein based oxygen sensing indicators. Sci. Rep. 2018, 8, 11849.
- Lemon, C.M.; Hanley, D.; Batka, A.E.; Marletta, M.A. Ratiometric Oxygen Sensing with H-NOX Protein Conjugates. Inorg. Chem. 2022, 61, 10521–10532.
- Solomon, E.I.; Sundaram, U.M.; Machonkin, T.E. Multicopper Oxidases and Oxygenases. Chem. Rev. 1996, 96, 2563–2606.
- Zauner, G.; Lonardi, E.; Bubacco, L.; Aartsma, T.J.; Canters, G.W.; Tepper, A.W.J.W. Tryptophan-to-Dye Fluorescence Energy Transfer Applied to Oxygen Sensing by Using Type-3 Copper Proteins. Chem. Eur. J. 2007, 13, 7085–7090.
- Solomon, E.I.; Chen, P.; Metz, M.; Lee, S.-K.; Palmer, A.E. Oxygen Binding, Activation, and Reduction to Water by Copper Proteins. Angew. Chem. Int. Ed. 2001, 40, 4570–4590.
- Erker, W.; Sdorra, S.; Basché, T. Detection of Single Oxygen Molecules with Fluorescence-Labeled Hemocyanins. J. Am. Chem. Soc. 2005, 127, 14532–14533.
- Østergaard, H.; Henriksen, A.; Hansen, F.G.; Winther, J.R. Shedding light on disulfide bond formation: Engineering a redox switch in green fluorescent protein. Embo J. 2001, 20, 5853–5862.
- Håkansson, K.O.; Winther, J.R. Structure of glutaredoxin Grx1p C30S mutant from yeast. Acta Crystallogr. D Biol. Crystallogr. 2007, 63 Pt 3, 288–294.
- Christie, J.M.; Hitomi, K.; Arvai, A.S.; Hartfield, K.A.; Mettlen, M.; Pratt, A.J.; Tainer, J.A.; Getzoff, E.D. Structural Tuning of the Fluorescent Protein iLOV for Improved Photostability. J. Biol. Chem. 2012, 287, 22295–22304.
- Schröder, E.; Littlechil, J.A.; Lebedev, A.A.; Errington, N.; Vagin, A.A.; Isupov, M.N. Crystal structure of decameric 2-Cys peroxiredoxin from human erythrocytes at 1.7 Å resolution. Structure 2000, 8, 605–615.
- Østergaard, H.; Tachibana, C.; Winther, J.R. Monitoring disulfide bond formation in the eukaryotic cytosol. J. Cell Biol. 2004, 166, 337–345.
- Björnberg, O.; Østergaard, H.; Winther, J.R. Mechanistic insight provided by glutaredoxin within a fusion to redox-sensitive yellow fluorescent protein. Biochemistry 2006, 45, 2362–2371.
- Morgan, B.; Sobotta, M.C.; Dick, T.P. Measuring E(GSH) and H2O2 with roGFP2-based redox probes. Free Radic. Biol. Med. 2011, 51, 1943–1951.
- Gutscher, M.; Pauleau, A.-L.; Marty, L.; Brach, T.; Wabnitz, G.H.; Samstag, Y.; Meyer, A.J.; Dick, T.P. Real-time imaging of the intracellular glutathione redox potential. Nat. Methods 2008, 5, 553–559.
- Trautsch, I.; Heta, E.; Soong, P.L.; Levent, E.; Nikolaev, V.O.; Bogeski, I.; Katschinski, D.M.; Mayr, M.; Zimmermann, W.H. Optogenetic Monitoring of the Glutathione Redox State in Engineered Human Myocardium. Front. Physiol. 2019, 10, 272.
- Mohring, F.; Jortzik, E.; Becker, K. Comparison of methods probing the intracellular redox milieu in Plasmodium falciparum. Mol. Biochem. Parasitol. 2016, 206, 75–83.
- Hatori, Y.; Kubo, T.; Sato, Y.; Inouye, S.; Akagi, R.; Seyama, T. Visualization of the Redox Status of Cytosolic Glutathione Using the Organelle- and Cytoskeleton-Targeted Redox Sensors. Antioxidants 2020, 9, 129.
- Pal, R.; Basu Thakur, P.; Li, S.; Minard, C.; Rodney, G.G. Real-time imaging of NADPH oxidase activity in living cells using a novel fluorescent protein reporter. PLoS ONE 2013, 8, e63989.
- Henríquez-Olguín, C.; Renani, L.B.; Arab-Ceschia, L.; Raun, S.H.; Bhatia, A.; Li, Z.; Knudsen, J.R.; Holmdahl, R.; Jensen, T.E. Adaptations to high-intensity interval training in skeletal muscle require NADPH oxidase 2. Redox Biol. 2019, 24, 101188.
- Bhaskar, A.; Chawla, M.; Mehta, M.; Parikh, P.; Chandra, P.; Bhave, D.; Kumar, D.; Carroll, K.S.; Singh, A. Reengineering redox sensitive GFP to measure mycothiol redox potential of Mycobacterium tuberculosis during infection. PLoS Pathog. 2014, 10, e1003902.
- Van Loi, V.; Antelmann, H. Method for measurement of bacillithiol redox potential changes using the Brx-roGFP2 redox biosensor in Staphylococcus aureus. MethodsX 2020, 7, 100900.
- Van Loi, V.; Harms, M.; Müller, M.; Huyen, N.T.T.; Hamilton, C.J.; Hochgräfe, F.; Pané-Farré, J.; Antelmann, H. Real-Time Imaging of the Bacillithiol Redox Potential in the Human Pathogen Staphylococcus aureus Using a Genetically Encoded Bacilliredoxin-Fused Redox Biosensor. Antioxid. Redox Signal 2017, 26, 835–848.
- Ebersoll, S.; Bogacz, M.; Günter, L.M.; Dick, T.P.; Krauth-Siegel, R.L. A tryparedoxin-coupled biosensor reveals a mitochondrial trypanothione metabolism in trypanosomes. Elife 2020, 9, e53227.
- Sugiura, K.; Nagai, T.; Nakano, M.; Ichinose, H.; Nakabayashi, T.; Ohta, N.; Hisabori, T. Redox sensor proteins for highly sensitive direct imaging of intracellular redox state. Biochem. Biophys. Res. Commun. 2015, 457, 242–248.
- Sugiura, K.; Yokochi, Y.; Fu, N.; Fukaya, Y.; Yoshida, K.; Mihara, S.; Hisabori, T. The thioredoxin (Trx) redox state sensor protein can visualize Trx activities in the light/dark response in chloroplasts. J. Biol. Chem. 2019, 294, 12091–12098.
- Sugiura, K.; Mihara, S.; Fu, N.; Hisabori, T. Real-time monitoring of the in vivo redox state transition using the ratiometric redox state sensor protein FROG/B. Proc. Natl. Acad. Sci. USA 2020, 117, 16019–16026.
- Heinemann, U.; Hahn, M. Circular permutation of polypeptide chains: Implications for protein folding and stability. Prog. Biophys. Mol. Biol. 1995, 64, 121–143.
- Baird, G.S.; Zacharias, D.A.; Tsien, R.Y. Circular permutation and receptor insertion within green fluorescent proteins. Proc. Natl. Acad. Sci. USA 1999, 96, 11241–11246.
- Topell, S.; Hennecke, J.; Glockshuber, R. Circularly permuted variants of the green fluorescent protein. FEBS Lett. 1999, 457, 283–289.
- Belousov, V.V.; Fradkov, A.F.; Lukyanov, K.A.; Staroverov, D.B.; Shakhbazov, K.S.; Terskikh, A.V.; Lukyanov, S. Genetically encoded fluorescent indicator for intracellular hydrogen peroxide. Nat. Methods 2006, 3, 281–286.
- Bilan, D.S.; Pase, L.; Joosen, L.; Gorokhovatsky, A.Y.; Ermakova, Y.G.; Gadella, T.W.J.; Grabher, C.; Schultz, C.; Lukyanov, S.; Belousov, V.V. HyPer-3: A Genetically Encoded H2O2 Probe with Improved Performance for Ratiometric and Fluorescence Lifetime Imaging. ACS Chem. Biol. 2013, 8, 535–542.
- Markvicheva, K.N.; Bilan, D.S.; Mishina, N.M.; Gorokhovatsky, A.Y.; Vinokurov, L.M.; Lukyanov, S.; Belousov, V.V. A genetically encoded sensor for H2O2 with expanded dynamic range. Bioorg Med. Chem. 2011, 19, 1079–1084.
- Pak, V.V.; Ezeriņa, D.; Lyublinskaya, O.G.; Pedre, B.; Tyurin-Kuzmin, P.A.; Mishina, N.M.; Thauvin, M.; Young, D.; Wahni, K.; Martínez Gache, S.A.; et al. Ultrasensitive Genetically Encoded Indicator for Hydrogen Peroxide Identifies Roles for the Oxidant in Cell Migration and Mitochondrial Function. Cell Metab. 2020, 31, 642–653.e6.
- Lee, C.; Lee, S.M.; Mukhopadhyay, P.; Kim, S.J.; Lee, S.C.; Ahn, W.-S.; Yu, M.-H.; Storz, G.; Ryu, S.E. Redox regulation of OxyR requires specific disulfide bond formation involving a rapid kinetic reaction path. Nat. Struct. Mol. Biol. 2004, 11, 1179–1185.
- Fan, Y.; Makar, M.; Wang, M.X.; Ai, H.-W. Monitoring thioredoxin redox with a genetically encoded red fluorescent biosensor. Nat. Chem. Biol. 2017, 13, 1045–1052.
- Fan, Y.; Ai, H.W. Development of redox-sensitive red fluorescent proteins for imaging redox dynamics in cellular compartments. Anal. Bioanal. Chem. 2016, 408, 2901–2911.
- Simen Zhao, B.; Liang, Y.; Song, Y.; Zheng, C.; Hao, Z.; Chen, P.R. A Highly Selective Fluorescent Probe for Visualization of Organic Hydroperoxides in Living Cells. J. Am. Chem. Soc. 2010, 132, 17065–17067.
- Herrou, J.; Crosson, S. Function, structure and mechanism of bacterial photosensory LOV proteins. Nat. Rev. Microbiol. 2011, 9, 713–723.
- Gu, Y.Z.; Hogenesch, J.B.; Bradfield, C.A. The PAS superfamily: Sensors of environmental and developmental signals. Annu. Rev. Pharmacol. Toxicol. 2000, 40, 519–561.
- Sakai, T.; Kagawa, T.; Kasahara, M.; Swartz, T.E.; Christie, J.M.; Briggs, W.R.; Wada, M.; Okada, K. Arabidopsis nph1 and npl1: Blue light receptors that mediate both phototropism and chloroplast relocation. Proc. Natl. Acad. Sci. USA 2001, 98, 6969–6974.
- Christie, J.M.; Salomon, M.; Nozue, K.; Wada, M.; Briggs, W.R. LOV (light, oxygen, or voltage) domains of the blue-light photoreceptor phototropin (nph1): Binding sites for the chromophore flavin mononucleotide. Proc. Natl. Acad. Sci. USA 1999, 96, 8779–8783.
- Crosson, S.; Rajagopal, S.; Moffat, K. The LOV domain family: Photoresponsive signaling modules coupled to diverse output domains. Biochemistry 2003, 42, 2–10.
- Ravikumar, Y.; Nadarajan, S.P.; Lee, C.S.; Rhee, J.K.; Yun, H.D. A New-Generation Fluorescent-Based Metal Sensor—iLOV Protein. J. Microbiol. Biotechnol. 2015, 25, 503–510.
- Zhao, H.; Zhang, Y.; Pan, M.; Song, Y.; Bai, L.; Miao, Y.; Huang, Y.; Zhu, X.; Song, C.-P. Dynamic imaging of cellular pH and redox homeostasis with a genetically encoded dual-functional biosensor, pHaROS, in yeast. J. Biol. Chem. 2019, 294, 15768–15780.
- Shcherbo, D.; Merzlyak, E.M.; Chepurnykh, T.V.; Fradkov, A.F.; Ermakova, G.V.; Solovieva, E.A.; Lukyanov, K.A.; Bogdanova, E.A.; Zaraisky, A.G.; Lukyanov, S.; et al. Bright far-red fluorescent protein for whole-body imaging. Nat. Methods 2007, 4, 741–746.
- Enyedi, B.; Zana, M.; Donkó, Á.; Geiszt, M. Spatial and temporal analysis of NADPH oxidase-generated hydrogen peroxide signals by novel fluorescent reporter proteins. Antioxid. Redox Signal 2013, 19, 523–534.
- Langford, T.F.; Huang, B.K.; Lim, J.B.; Moon, S.J.; Sikes, H.D. Monitoring the action of redox-directed cancer therapeutics using a human peroxiredoxin-2-based probe. Nat. Commun. 2018, 9, 3145.