Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Jin-Xian Gao and Version 2 by Lindsay Dong.
Rapid eye movement (REM) sleep is the main sleep correlate of dreaming. Ponto-geniculo-occipital (PGO) waves are a signature of REM sleep. They represent the physiological mechanism of REM sleep that specifically limits the processing of external information. PGO waves look just like a message sent from the pons to the lateral geniculate nucleus of the visual thalamus, the occipital cortex, and other areas of the brain. The dedicated visual pathway of PGO waves can be interpreted by the brain as visual information, leading to the visual hallucinosis of dreams. PGO waves are considered to be both a reflection of REM sleep brain activity and causal to dreams due to their stimulation of the cortex.
- rapid eye movement sleep
- ponto-geniculo-occipital waves
- non-rapid eye movement sleep
1. Introduction
Dreams are images and experiences that people have while they sleep. In ancient cultures, dreams were believed to contain messages from the gods or omens of the future. In the late 19th century, dreams became a subject of study for psychologists and psychoanalysts. Sigmund Freud’s The Interpretation of Dreams [1] initially developed the most prominent psychoanalytic theory of dreams. This model has been quite influential in sleep research and continues to have strong adherents to this day. For Freud, the dream is a highly meaningful mental product that is the result of specific mental processes under the conditions of sleep. Dreams that have “hidden meanings” and “repressed desires” are well established in popular folk psychology. Although Freud theorized that the purpose of dreaming is wish fulfillment, there is little experimental evidence to support this concept.
Modern sleep science has evolved significantly since the late 1950s, largely due to the seminal discoveries of rapid eye movement (REM) sleep [2][3][4][5][2,3,4,5]. Aserinsky and Kleitman [2] found that 74% of awakenings from REM sleep resulted in the recall of a dream, as compared to only 9% of awakenings from non-REM (NREM) sleep. The linking of REM sleep to dreaming ushered in a new era in the study of dreams [6]. During REM sleep, in addition to rapid eye movements (REMs), cortical electroencephalogram (EEG) desynchronization (or activation), loss of muscle tone, and autonomic fluctuations [2][3][4][5][2,3,4,5], many other physiological and behavioral features have also been found in humans and other mammals. These include high-amplitude spiky potentials of ponto-geniculo-occipital (PGO) waves, high-amplitude hippocampal EEG theta waves, penile erections, sporadic limb twitching, increases in brain/body temperature, and an elevated arousal threshold [7][8][9][10][11][12][13][14][15][16][17][7,8,9,10,11,12,13,14,15,16,17]. In these characteristic components of REM sleep, PGO waves are undoubtedly powerful internal sensory signals that convey a large amount of information to the visual cortex and seem to “compose the song sheet of dreams” [18]. Accordingly, a “dream state generator”, located mainly in the pontine reticular formation and producing PGO waves, has been postulated to be the cause of both REMs and the periodic intrusion of new content into hallucinatory dreams [19][20][21][22][19,20,21,22].
2. The Neural Mechanisms Underlying Dreaming
2.1. PGO Waves
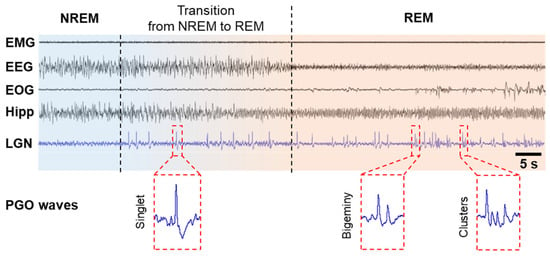
PGO wave activity was first discovered in cats in the 1950s [5][23][24][5,23,24]. Because these local field potentials originate in the pons (P) and propagate to the lateral geniculate nucleus (G) of the visual thalamus and the occipital cortex (O), they are called PGO waves [18][25][26][18,25,26]. PGO waves occur just before the onset of REM sleep and continue throughout its duration. They are characterized as biphasic, sharp field potentials lasting 60–120 ms, with an amplitude of 200–300 μV, occurring as singlets and clusters (Figure 1). These spikes during REM sleep period are in parallel to eye saccades and are observed not only in cats [5][25][27][28][29][5,25,27,28,29] but also in rats [30][31][30,31], mice [32], in non-human primates such as macaques [33][34][33,34], baboons [35], and in humans [7][36][37][38][7,36,37,38]. Cholinergic/glutamatergic neurons in the pontine brainstem have been shown to generate PGO waves by burst firing. PGO waves look just like a message sent from the pons to the lateral geniculate nucleus (LGN), occipital cortex, and other brain regions, including the temporal and prefrontal cortices and the amygdala. The dedicated visual pathway of PGO waves could be interpreted by the brain as visual information, thereby leading to the visual hallucinosis of dreams. These waves are not only associated with dream production but also limit the cortex’s ability to process external inputs [39][40][39,40].Figure 1. PGO waves in cats. PGO waves occur just before the onset of REM sleep, i.e., during the transition from NREM to REM, and during REM sleep period. PGO wave singlets occur predominantly during the transition from NREM to REM sleep and are not time-locked to rapid eye movements (REMs), but PGO wave clusters (≥3 waves) occur predominantly during REM sleep and correlate strongly with REMs and typical hippocampal theta oscillations. Abbreviations: EEG, electroencephalogram of the neocortex; EMG, electromyogram; EOG, electro-oculogram; Hipp, EEG of the hippocampus; LGN, EEG of the lateral geniculate nucleus.
In humans, many invasive and non-invasive studies provide insight into how PGO waves occur during REM sleep and how they contribute to dreaming. For example, in two invasive studies in Parkinson’s disease patients, deep brain stimulation electrodes were surgically implanted in the pedunculopontine nucleus of the pontine tegmentum [36] and the subthalamic nucleus (STN) [7]. The results showed that PGO-like waves were observed during REM sleep. Much like feline models, PGO wave singlets and clusters were recorded within STN during pre-REM and REM sleep [7].
Overall, much of the detailed understanding of PGO waves and their mechanisms still depends on animal studies. However, direct translation to humans may require further investigation and validation. Using noninvasive methods such as fMRI and magnetoencephalogram, the future of PGO wave research undoubtedly lies in broad studies that combine behavioral, pharmacological, physiological, and cognitive experiments in human subjects [38].
2.2. Activation–Synthesis Hypothesis
Hobson and McCarley, following the pioneering work of Jouvet [41][45], proposed the activation–synthesis hypothesis in 1977 [19]. This hypothesis is based primarily on microelectrode recordings of PGO waves in cats, which were found to occur primarily during REM sleep. The dreaming process consists of activation and synthesis. In short, the neural activity in the pons activates the brain, especially the LGN and visual cortex, to generate information during REM sleep. Under these conditions, in the absence of external stimuli, internally generated inputs randomly activate sensorimotor information, and the passive synthesis of this information (perceptual, conceptual, and emotional) creates dreams (Figure 2). During REM sleep, these internal inputs generated by PGO waves continuously activate the forebrain via the
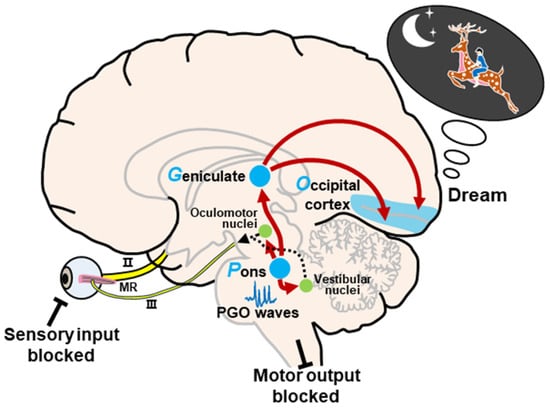
Figure 2. Schematic representation of the activation–synthesis model. Neural activity of PGO waves in the pons activates the lateral geniculate nucleus of the visual thalamus and the visual cortex. The passive synthesis of information (perceptual, conceptual, and emotional) generates dreams in the cortex. Meanwhile, the neural activity in the pons also activates the vestibular nucleus and oculomotor-related nuclei, such as the oculomotor nucleus, the trochlear nucleus, and the abducens nucleus, to induce REMs during REM sleep. In this state, external input and motor output are blocked. Abbreviations: II, optic nerve; III, oculomotor nerve; MR, medial rectus.
2.3. Activation, Input, and Modulation (AIM) Model
The activation–synthesis hypothesis of dreaming was updated and extended by Hobson et al. [21][22][21,22], and transformed into the AIM state-space model of the brain–mind isomorphism, where a conscious state can be understood as a point in a three-dimensional state space. In the AIM model, the variables consist of activation (A), input (I), and modulation (M) (Figure 3). (A) The level of brain activation can be defined as the average firing frequency of brainstem neurons as reflected by high and low frequency levels in the EEG. The high levels of A in REM sleep are a correlate of the mind’s ability to access and manipulate significant amounts of stored information from the brain during dream synthesis. (I) Input source is a measure of how much of the sensory data being processed are from external or internal sources. During REM sleep, internally generated PGO waves replace blocked external sensory input, activating sensory and affective centers that then prime the cortex for dream construction by stimulating the visual cortex with PGO waves. This can be estimated from the frequency of REMs in REM sleep, which is thought to reflect brainstem PGO and motor generator activity. (M) The brainstem neuromodulators that they release exert a broad chemical influence on the brain. It is well known that REM sleep is cholinergic potentiated and monoaminergic suppressed (Figure 3A).
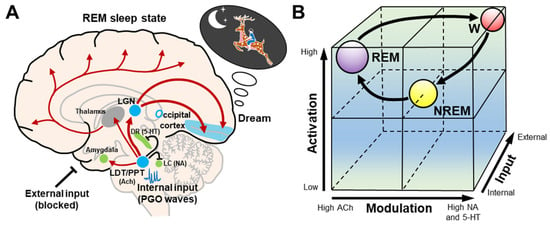
Figure 3. Schematic representation of the AIM model. (A) During REM sleep dreams, cholinergic neurons in the LDT/PPT are involved in the generation of internal input PGO waves, while serotonergic dorsal raphe nucleus and noradrenergic locus coeruleus neurons and external inputs are inhibited. These internal inputs activate the LGN and occipital cortex, and produce dreams. The LDT/PPT–thalamus–cerebral cortex pathway causes the desynchronization of the cerebral cortex. The LDT/PPT–amygdala pathway may be involved in mood regulation during dreams. (B) The cubic 3-dimensional model shows normal transitions within the 3-dimensional parameters (activation, input, and modulation) from wakefulness to NREM and then to REM sleep. Abbreviations: 5–HT, serotonergic; ACh, cholinergic; DR, dorsal raphe nucleus; LC, locus coeruleus; LDT/PPT, laterodorsal tegmental and pedunculopontine nuclei; LGN, lateral geniculate nucleus; NA, noradrenergic; NREM, non-rapid eye movement; REM, rapid eye movement; W, wakefulness.
2.4. Neuronal Mechanisms of REM Sleep Regulation
REM sleep is generally thought to be mediated by a neural network located primarily in the brainstem. More recently, the concept of REM sleep regulation has evolved. Several hypothalamic and forebrain networks, including newly identified neuropeptides such as orexin and melanin-concentrating hormone (MCH), have been implicated in both the control and the final expression of this behavioral state [14][16][17][42][43][44][45][46][47][14,16,17,49,50,51,52,53,54]. Firstly, a reciprocal interaction between REM on and REM off states has been proposed to occur in the brainstem. In this model, the cholinergic laterodorsal and pedunculopontine tegmental neurons (LDT/PPT) are REM-on cells. The serotonergic dorsal raphe nucleus and noradrenergic locus coeruleus neurons are REM-off cells [22][48][49][22,55,56] (Figure 3A). However, this reciprocal interaction may not be sufficient to produce REM sleep, as suggested by experimental evidence over the past decade [16][46][16,53]. The glutamateric sublaterodorsal tegmental nucleus (SLD) and the GABAergic lateral paragigantocellular nucleus (LPGi) have been subsequently found to act as REM-on neurons, whereas GABAergic ventrolateral periaqueductal gray matter (vlPAG) and lateral pontine tegmentum (LPT) act as REM-off neurons [16][46][16,53].