Cardiac regeneration is an ancestral trait in vertebrates, a general capacity that seems to be inversely correlated with evolutionary complexity across the animal kingdom. To better understand the biology of MI, as well as to develop different therapeutic strategies, in vitro, ex vivo and in vivo models have been developed.
- cardiac disease
- myocardial infarction
- cell cycle
- heart regeneration
1. Introduction
2. Experimental Models of Cardiac Regeneration
Cardiac regeneration is an ancestral trait in vertebrates, a general capacity that seems to be inversely correlated with evolutionary complexity across the animal kingdom [14][15][14,15]. For example, the human heart has a very limited capacity for cardiac regeneration in contrast to fish and amphibian organisms [16][17][16,17]. Some years ago, Field’s lab evidenced that a low percentage of mouse ventricular CMs have proliferative capacity in normal conditions in mice, becoming higher after injury [18]. In the same line, it has been demonstrated that 1% of human CMs are renewed each year [12]. In a cardiac injured scenario, this low percentage of CM renewal is not enough to repair the damaged myocardium. Some laboratories, within the cardiovascular field, are focusing their efforts on increasing the proliferative ability of CMs in order to enhance the regenerative capacity of mammalian hearts. To better understand the biology of MI, as well as to develop different therapeutic strategies, in vitro, ex vivo and in vivo models have been developed. The first approach is the in vitro cardiac model which implies primary cultures and/or cell lines. Concretely, CMs can be obtained from animal or human hearts or by the differentiation process from stem cells. In general terms, CMs from primary culture have some limitations for the study of cardiac physiology, such as low proliferative capacity and the absence of spontaneous beat, among others [19][20][21][22][23][19,20,21,22,23]. As an alternative, differentiated CMs derived from pluripotent cells, mesenchymal stem cells (MSCs), embryonic stem cells (ESCs) or induced pluripotent stem cells (iPSCs), are essential for the development of MI models [24][25][26][27][28][24,25,26,27,28]. On the other hand, ex vivo models involve keeping the heart outside of the body in a normal or retrograde perfusion system. This kind of model offers the opportunity to generate ischemic and reperfusion situations to analyze the effects of MI [29][30][31][36,37,38]. The main advantage of this model is that the infarct size can be measured and left ventricular function can be easily assessed [32][33][34][35,39,40]. Last but not least are the in vivo models: they represent the most used model to test MI effects as well as for drug and safety studies. This model offers the opportunity of analyzing the vast majority of physiological changes generated in response to MI, i.e., inflammatory processes and scar formation as well as the possibility of identifying blood biomarkers [32][35]. In the in vivo scenario, researchers have the possibility of employing different strategies that enable the study of different molecular mechanisms, but need to take into consideration the variability associated to injury degree (mild, moderate and severe), as well as surgical and postoperative mortality [32][35].3. Injury Models to Study Cardiac Healing
Nowadays, the most important goal of cardiac researchers is to decipher the mechanisms that control heart regeneration in distinct animal models with the intention of stimulating cardiac repair in adult mammals. To perform this challenging goal, several different methods are applied to simulate a cardiac injury that is similar to the damage generated by MI. The cryoinjury technique is based on the exposition of the cardiac apical left ventricle area to a liquid nitrogen cooled cryoprobe [35][48] (Figure 1A). By using this technique, it is noteworthy that, depending on the damage severity, the cardiac regenerative response can be different. Transmural injury is the highest degree of damage with this technique, where the full wall diameter of the ventricle is affected, whereas non-transmural injury, where the cryoprobe does not penetrate the wall ventricle, inflicts mild damage [36][49].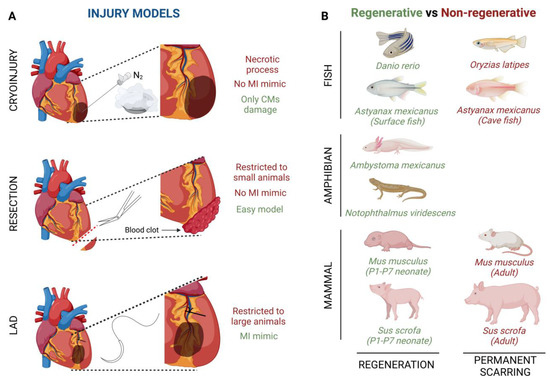
4. Heart Regeneration after Injury in Different Animal Models
4.1. Invertebrate Models
The ability to replace lost body parts or tissues is a phenomenon that is peculiar to a few organisms from different clades in the animal kingdom. Within the invertebrate group, hydras and planarians stand out, due to their ability to completely regenerate their bodies after an amputation [40][56]. These models allow us a first approach to the morphological and cellular dynamic changes involved in regeneration [41][57]. Hydras and planarians are two in vivo models with simple cellular organization that make them useful model systems to better study and understand stem cell behavior and regenerative processes in higher evolutionary models.4.2. Vertebrate Models
Fish models
Several fish species have shown the ability to repair an injured heart through the induction of CM proliferation (Figure 1B). For example, zebrafish (Danio rerio) have the ability of total heart regeneration 60 days after ~20% apical resection of the ventricle, by activating the proliferation rate of the CMs present in the surrounding injured area [16][42][16,58]. Something similar happens when ~25% of the zebrafish’s ventricle is damaged by using a cryoprobe. In this type of heart injury model, although the regeneration process is much slower, the heart achieves restoration in approximately ~180 days [43][44][45][59,60,61]. Finally, Poss’ lab developed a tamoxifen-inducible genetic ablation model that allowed the promotion of 60% of CM cell death. In this case, 30-day post tamoxifen induction they observed a complete re-muscularized ventricle [46][62]. Moreover, a few years ago, Gonzalez-Rosas et al. (2018) [47][63] reported that most of the zebrafish’s CMs contain only two sets of homologous chromosomes. These diploid CMs have the ability to proliferate, inducing heart regeneration [48][64]. Finally, considering that zebrafish resides in a hypoxic environment, this restrictive O2 condition enables CM dedifferentiation and proliferation leading to heart regeneration [42][58].
Amphibian models
Similar to fishes, some urodele amphibians have the ability to regenerate several tissues and organs [49][50][71,72] (Figure 1B). For example, axolotls (Ambystoma mexicanus), which are neotenic aquatic urodele amphibians, are capable of regenerating the heart after injury, resection and cryo-injury, but only in the larval phase [49][50][51][71,72,73]. However, when axolotls undergo metamorphosis and get into the adulthood stage, regeneration, in general terms, is reduced and some morphological defects are still observed [52][74]. This animal group loses its overall regenerative capacity as it undergoes metamorphosis, concomitantly with maturation of the immune system, which is comparable to that of mammals [53][54][75,76]. There are no studies about heart regeneration in adult axolotls, however it is important to highlight that thyroid hormone, which is necessary in this species to undergo metamorphosis into a land-dwelling adult, impairs heart regeneration in zebrafish [51][55][73,77]. In these terms, newts as Notophthalmus viridescens, bear heart regeneration capacity dependent on the type of injury [17][56][57][58][59][17,42,78,79,80] (Figure 1B).
Chicken models Avian cardiac regenerative capacity has not been thoroughly investigated. Burn lesions in chicken (Gallus gallus) myocardium resolve as regeneration in 3- and 5-day old chick embryos [60][61][83,84]. Furthermore, a study carried out by Novikov and Khloponin in 1984 demonstrated that chickens have the ability to repair cardiac damage at early embryonic stages. This process takes about 7 to 10 days after injury with the intervention of all three layers of the embryonic heart, i.e., epicardium, endocardium and myocardium [61][84]. Mammal models Lastly, it is widely known that the mammalian heart does not have the ability to regenerate after an injury process (Figure 1B). However, more than a decade ago, Porrello et al. (2011), revealed that heart regeneration was achieved in mice (Mus musculus) when apical resection was performed at postnatal day 1 (P1). This regenerative event lasted a period of 21 days and was carried out by proliferation of the existing CMs in the surrounding injured area [62][63][45,85]. Similarly, a full cardiac recovery is observed in mice after ligation of the left anterior descending (LAD) coronary artery in P1 [64][86]. On the other hand, the heart behavior after ventricle cryoinjury is different depending on severity; i.e., non-transmural cryoinjury in P1 mice undergoes healing, while P1 mice with transmural cryoinjury do not attain complete regeneration [36][65][66][49,87,88].