Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Charalampos Proestos and Version 2 by Peter Tang.
Radical oxygen species formed in human tissue cells by many endogenous and exogenous pathways cause extensive oxidative damage which has been linked to various human diseases.
- LDL-oxidation
- DNA-damage
- antioxidant vitamins
- oxidative stress
1. Introduction to Lipid Peroxidation and Antioxidants
Peroxidation of lipids, particularly of polyunsaturated fatty acids (PUFAs) is a process with marked implications: it shortens the shelf-life of food and drugs, it causes fragmentation of DNA, it damages cellular membranes and it promotes the genesis of many human diseases [1]. Lipid peroxidation is a complex biological process, initiated by free radicals, that results in the formation of conjugated dienes and lipid hydroperoxides [2]. These are usually degraded to a variety of products including alkanals, hydroxyalkenals, ketones, alkanes etc. [3]. Figure 1 reflects the lipid peroxidation process along with a few common oxidative biomarkers.
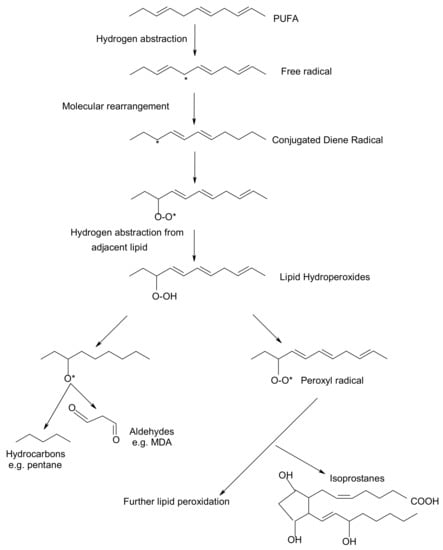
Figure 1.
Overview of lipid peroxidation process and oxidative biomarkers (* indicates the presence of free radical).
Free radicals and oxidants play a dual role as both toxic and beneficial compounds, since they can be either harmful or helpful to the body [4][5][4,5]. They are formed in tissue cells by various endogenous and exogenous pathways [6]. The ability of free radicals to structurally modify cellular components, gene expression and protein production has led to the implication of their involvement in a variety of pathological conditions, including inflammation, aging, carcinogenesis and cardiovascular diseases [7][8][7,8]. Oxidative stress exerts an adverse impact on human health. Oxygen free radicals (such as hydroxyl radicals, superoxide radicals and other active oxygen species including also singlet oxygen) adversely alter lipids, proteins, and DNA [9]. A role of lipid peroxidation and oxidative stress in the association between thyroid diseases and breast cancer has been claimed by Dominguez and Castelao (2008) [10]. Actually, overproduction of free radicals in vivo and the consequent damage to biological molecules is increasingly regarded as an important event in the development of human diseases, including arthritis, thyroid, cancer, and atherosclerosis [11].
Bhattacharyya et al. (2014) [12] noted that reactive oxygen species (ROS) are produced within the gastrointestinal (GI) tract, since ingested materials and microbial pathogens can induce oxidative injury and GI inflammatory responses involving the epithelium and immune/inflammatory cells. Therefore, further investigation on how the ROS can contribute to diverse gastrointestinal dysfunction, or manifest dual roles in cancer promotion or cancer suppression would enhance understanding of inflammation-based GI diseases and facilitate the development of new therapies [13].
Certain oxidative biomarkers have linked oxidative stress and the development of health diseases. A substantial body of evidence indicates that measurement of prostaglandine (PG)-like compounds provides a direct and reliable approach to assess oxidative damage in vivo compared with other methods such as thiobarbituric acid reacting substances (TBARS) that have been also widely studied [14]. According to Barocas et al. (2011) [15] oxidative stress measured by urine F2-isoprostane level is associated with prostate cancer. In recent years, development of immunochemical detection of 4-Hydroxynonenal (HNE)-histidine adducts opened more advanced methodological possibilities for qualitative and quantitative detection of lipid peroxidation in various human and animal tissues [16].
To control and reduce in vivo oxidative damage, nature makes use of several types of antioxidants or radical trapping agents operating at different stages of the process [17][18][17,18]. These compounds, also known as biological antioxidants, react rapidly with free radicals and slow down the oxidative damage [19][20][19,20]. A body of evidence indicates that certain dietary compounds of plant origin can act as radical scavengers in model biological systems and in the human organism, thereby acting as dietary antioxidants [21][22][21,22]. Increased plasma total antioxidant capacity has been associated with a high consumption of fruits and vegetables rich in these vitamins, although limited information is available on whether this reflects the dietary intake of antioxidants [23]. The commonly used assays for ranking antioxidants share a common problem. Most estimates are based on methods conducted in solution and are, therefore, not necessarily relevant to processes that occur at the lipid–water interfaces in both membranes and micro emulsions, e.g., lipoproteins [24]. This resviearchw focuses on several natural compounds the levels of which in human body can be manipulated by supplements and dietary modifications. More specifically, the following ones have been reported to exert in vitro and in vivo antioxidant activities:
(i) Tocopherols and tocotrienols (vitamin E). Tocopherols and tocotrienols comprise a group of eight chromanol homologs extracted from natural sources (e.g., oils, nuts, leafy vegetables) that possess vitamin E activity in the diet [25]. They are natural monophenolic compounds with well-established antioxidant activities in food and biological systems [26]. The α-, β-, γ- and δ-tocopherols are characterized by a saturated side chain consisting of three isoprenoid units, whereas their corresponding tocotrienols have double bonds at the 3′, 7′ and 11′ position of the isoprenoid side chain [27] (structures are presented in Figure 2).
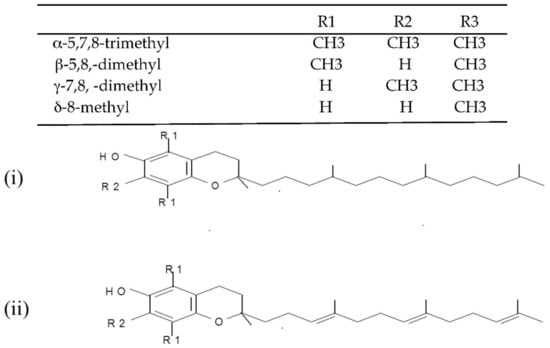
Figure 2.
Chemical structures of the E vitamers (tocopherols-i and tocotrienols-ii).
(ii) L-Ascorbic acid (vitamin C). Vitamin C, also known as ascorbic acid, occurs in all tissues of living organisms where it is responsible for the normal functioning of important metabolic processes [28]. It is very widespread in nature (e.g., in oranges, green peppers, watermelon, grapefruit) and recognized as an antioxidant nutrient with multi-functional effects depending on the conditions of the food and biological systems [29]. L-Ascorbic acid is a six-carbon weak acid with a pKa of 4.2, which is reversibly oxidized due to its enediol structure with the loss of an electron to form the free radical semihydroascorbic acid [21].
(iii) Carotenoids (provitamins A). The carotenoids are natural pigments extracted from many sources (e.g., in carrots, plums, apricots, tomatoes, spinach) that are used for various food applications. Carotenoids have been increasingly studied in the last decade for their potential to act as in vitro and in vivo antioxidants [30][31][30,31]. Dietary supplementation with certain carotenoids possessing provitamin A activity (such as β-carotene and lycopene, Figure 3) has been associated in literature with a protective role against diseases (including aging, types of cancer, cardiovascular disease, cataracts, and age-related macular degeneration) [32].
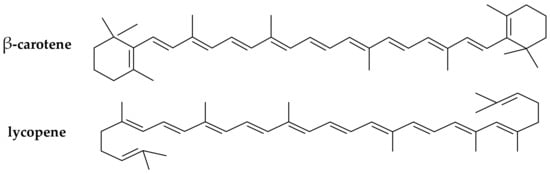
Figure 3.
Chemical structure of the main provitamin A carotenes (β-carotene, lycopene).
(iv) Flavonoids and phenolic acids. The current work also reports the dietary antioxidant effects of various phenolic acids available in many natural sources (e.g., in olive oil, herbs, fruits) that have been widely explored in food systems [33][34][33,34]. Flavonoids, in particular, comprise a class of phenolic compounds with well established antioxidant properties strongly related to their structure [35].
2. Low-Density Lipoprotein (LDL) Oxidative Damage and Antioxidation
2.1. Link of LDL Oxidation to Atherogenesis and Common Monitoring Methods
Link of LDL Oxidation to Atherogenesis and Common Monitoring Methods
Human low-density lipoprotein (LDL) is defined as the population of lipoproteins, which can be isolated from plasma by ultra centrifugation within a density gradient of 1019–1063 g/L [36][37]. Elevated plasma concentration of LDL is a risk factor for atherosclerosis and coronary artery disease [37][38]. A recent body of literature has reported that atherosclerosis develops following free radical processes that cause oxidative modification of LDL [38][39][39,40]. Atherosclerosis is a progressive disease of the arterial tree that involves deposition of cholesterols in the arterial intima leading finally to a thickening of the arterial wall and reduced luminal blood flow [40][41]. The lipid deposited is mainly LDL, derived from the circulation. Amarowicz and Pegg [41][42] claimed that the exact mechanism(s) of atherosclerosis in humans remains elusive, but one theory hypothesizes that this deleterious process results from the oxidative modification of LDL. The oxidation of LDL is a complex process during which both the proteins and the lipids undergo oxidative changes and form complex products [42][43]. It is a lipid peroxidation reaction driven by free radicals. Reactive oxygen species, or thiols may be released and thus participate directly in the initiation of LDL oxidation [43][44]. Oxidation presumably begins when a reactive radical abstracts hydrogen from a PUFA of surface phospholipids or bulk lipids in the core of the LDL particle, a reaction which in the absence of sufficient concentrations of antioxidants results in the propagation of lipid peroxidation [44][45]. There are various ways to measure the effect of a diet on LDL oxidation, including: the level of thiobarbituric acid reactive substances (TBARS) and lipid hydroperoxides in plasma; anion exchange chromatography and electrophoresis motility of the LDL particles; the formation of conjugated dienes (CD) at 232 nm and fluorescence spectra on the oxidation of native LDL with a chemical inducer; and the uptake of oxidised LDL by macrophages [45][46][36,46]. Given their responsiveness to targeted nutritional interventions, markers of LDL oxidation have been employed in a rapidly growing number of clinical studies for more than two decades [47]. The evaluation of LDL oxidation in vivo is difficult, and most of the investigations deal with in vitro oxidised LDL, a process accompanied by characteristic changes of physicochemical and biological properties [38][48][39,48]. The most common method for the determination of antioxidant properties of natural phenolic compounds is the LDL oxidation assay. LDL is isolated from human plasma, and oxidation is induced by Cu2+ ions and is monitored spectrophotometrically via the change of CD absorption at 232 nm [41][45][36,42]. Subsequently, the initiators break down the existing lipid hydroperoxides and initiate the propagation stage according to the following reactions: (a) Cu+ + LOOH → Cu2+ +OH− + LO·(b) Cu+ + HOOH → Cu2+ +OH− + HO