1. Introduction
“Pigeon” is the collective name for hundreds of breeds of birds in the
Columbidae family
[1]. As one of the earliest domesticated birds, it is important economically and for entertainment due to its ornamental and racing value, as well as its use for meat and eggs. At present, it has become an ideal animal model for analyzing bird behavior and the physiological regulation of reproduction due to its well-timed breeding cycle and biparental care
[2][3][2,3].
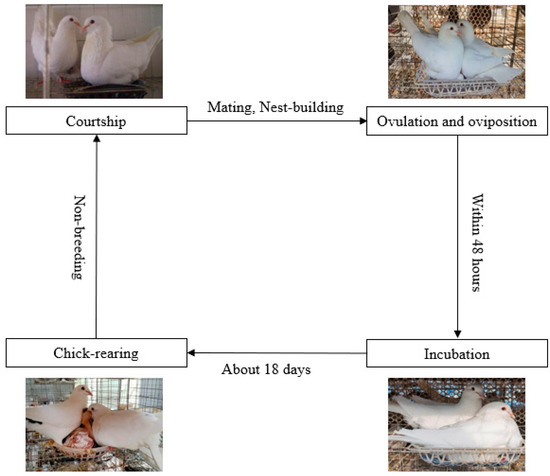
Pigeons are non-seasonal breeders, and their breeding cycle includes non-breeding, courtship and mating; nest-site selection and nest building; incubation; and feeding of the squabs
[4]. The birds pair off in a pattern of one female and one male for their entire life. The female usually lays two eggs within 48 h, and then the parents take turns to hatch the egg
[5]. There is a clear time difference between male and female incubation activities, with males generally incubating at noon and females incubating for the rest of the day
[6]. After about 18 days of incubation, the squabs are taken care of by their parents, and the adults regurgitate food to their squabs in a mouth-to-mouth manner (
Figure 1)
[7][8][9][10][7,8,9,10]. Similar situations also occur among flamingos and emperor penguins
[11][12][11,12]. Amazingly, the growth rate of pigeons is much higher than that of other poultry. The relative growth index of squabs is 3.79 and 1.96 times higher than that of large fast-growing chickens and quails, respectively
[10].
Figure 1.
Breeding cycle of pigeons.
2. The Formation Mechanism of Pigeon Milk
The formation of pigeon milk is involved in a very complex regulatory network. Two biological processes are involved. One is the proliferation and shedding of crop epidermal cells, and the other is the accumulation of nutrients in cells. Under the stimulation of hormones, especially prolactin, crop cells proliferate massively, and the proliferation of the crop epidermis is also affected by non-coding RNAs. Meanwhile, a large number of nutrients, such as proteins and lipids, are synthesized in crop epidermal cells. When nutrients have accumulated to a certain level, apoptosis of epidermal cells occurs, which leads to the shedding of a mass of epithelial cells that are full of nutrients to form pigeon milk.
2.1. The Proliferation of Pigeon Crop Epidermal Cells
2.1. The Proliferation of Pigeon Crop Epidermal Cells
2.1.1. Morphological Changes in the Crop
In birds, the crop is located between the distal esophagus and the proximal end of the proventriculus. In the nonlactating phase, the crop wall of pigeons is thin, and there is no lateral lobe structure
[13][87]. In preparation for lactation, the number and depth of rete pegs increases significantly and the lamina propria becomes progressively more extended and narrow as the crop further differentiates
[9]. This largely increases the surface area of the crop to provide the histological basis for the mass production of crop milk. During the lactation period, in response to prolactin, the rapid proliferation of the germ cell layer of crop tissue leads to a convoluted, highly folded epithelial structure
[14][15][88,89]. The whole crop wall is thickened with two obvious lateral lobe structures, and its weight and volume are significantly enlarged
[8][13][8,87]. The nutritive cell layer is then formed as the vasculatures emerge. Finally, the nutrient-loaded cells are shed to produce crop milk
[9].
2.1.2. The Regulation of Hormones on Crop Proliferation
Prolactin plays an important role in the regulation of reproduction in vertebrates. Riddle et al.
[16][90] first discovered and purified prolactin from the anterior pituitary of pigeons. The level of prolactin rose in adult pigeons during the latter half of the incubation period, and the development of the crop sac directly followed increased hormone secretion
[17][55]. Injection of exogenous prolactin significantly increased crop weight
[18][19][91,92] and levels of DNA and RNA
[19][92], and it also induced expression of specific genes
[19][20][21][92,93,94]. There are at least two modes by which prolactin acts as a mitogen on the epithelium of crop: First, prolactin acts directly on the crop mucosa to enhance the sensitivity of the crop to a somatomedin-like growth factor that functions to promote mitosis; second, prolactin heightens the synthesis and secretion of somatomedin-like growth factor, which then acts synergistically with prolactin to stimulate the proliferation of crop mucosa cells
[22][95]. Studies have shown that ornithine decarboxylase activity is closely related to cell proliferation
[23][24][96,97]. Prolactin increased ornithine decarboxylase activity and promoted DNA synthesis and cell proliferation in human promyelocytic cells
[25][98]. Likewise, prolactin injection significantly increased ornithine decarboxylase (ODC) activity in pigeon crop
[19][92]. Therefore, the rapid proliferation of the crop during lactation may be related to the increased activity of ornithine decarboxylase induced by prolactin stimulation.
Relaxin is mainly produced by the corpus luteum and attains high plasma levels during pregnancy
[26][99]. Studies have shown that relaxin could cause significant development of the mammary gland in rats
[27][28][29][100,101,102]. Likewise, relaxin was reported to enhance the cell growth and differentiation of the crop sac, which induces a significant increase in crop weight
[30][31][103,104]. Bani et al.
[32][105] also found that the changes in the crop sac following prolactin action were similar to those following relaxin injection. Therefore, relaxin may be involved in pigeon milk formation by promoting crop proliferation. Instead, estradiol, progesterone and growth hormone may not contribute to crop milk formation
[33][75]. In addition, the concentrations of EGF and IGF-1 in pigeon crop homogenates were notably elevated during the breeding period
[33][75], and injection of EGF significantly enhanced the local growth of pigeon crop-sac mucosal epithelium
[34][106]. Thus, EGF may also be involved in crop milk formation.
2.1.3. The Regulation of Crop Proliferation by Non-Coding RNA
Regulatory non-coding RNAs consist of small non-coding RNAs (small ncRNAs) and long non-coding RNAs (lncRNAs). Small non-coding RNAs include circular RNAs (circRNAs), microRNAs (miRNAs), and small interfering RNAs (siRNAs). Studies have shown that regulatory non-coding RNAs regulate mammary gland development and proliferation in mammals
[35][36][107,108]. For example, circHIPK3 promoted mammary epithelial cells proliferation in bovine
[37][109]. Comparing the expression profiles of miRNAs in the crop of lactating and nonlactating female pigeons, a total of 71 miRNAs were significantly differentially expressed. The target genes of these miRNAs were mainly involved in cell development and epithelial cell morphogenesis, which indicates that these miRNAs could regulate crop cell proliferation to affect pigeon milk formation
[38][110]. In addition, miR-193-5p bound to a conserved site in the 3′-untranslated region (UTR) of phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit delta (PIK3CD) mRNA to negatively regulate its expression to promote the proliferation of pigeon crop fibrocyte
[39][111].
2.2. Accumulation of Nutrients in Pigeon Crop Epithelial Cells
2.2.1. Synthesis of Protein in Crop Epidermal Cells
Amino acids are the building blocks in the process of protein synthesis that control various metabolic pathways important for whole-body homeostasis
[40][112]. During lactation, the process of amino acid uptake from circulation into the mammary gland was strengthened
[41][113], so amino acid concentrations in plasma are often considered an indicator of milk protein synthesis
[42][114]. Xie et al.
[43][60] found high levels of essential and non-essential amino acids in plasma on Day 1 or 7 of the chick-rearing period, and the expression of amino acid transporters and enzymes related to amino acid synthesis in the crop tissue was also higher at this point in time. This suggests that both amino acid transportation and de novo synthesis are necessary for the synthesis of proteins in crop cells.
Under the stimulation of prolactin and insulin, a series of signaling pathways were activated to enhance protein synthesis in crop epidermal cells, such as the target of rapamycin (TOR), Janus kinases/signal transducer and activator of transcription proteins (JAK/STAT), and insulin receptor substance 1/protein kinase B/target of rapamycin (IRS1/Akt/TOR) signaling pathway. In mammals, the mammalian target of rapamycin (mTOR) signaling pathway has been shown to play an important role in the synthesis of milk proteins by changing the phosphorylation status of downstream effector proteins, such as eukaryotic initiation factor 4E binding protein 1 (4EBP1) and ribosomal protein S6 kinase (70S6K)
[44][45][46][47][48][115,116,117,118,119]. Targeted disruption of the target of rapamycin complex 1 (TORC1) inhibited protein synthesis in pigeon crop, thereby retarding the growth of squabs
[49][120]. Dietary supplementation with leucine promoted the growth of squabs and increased the synthesis of pigeon milk protein through the TOR signaling pathway
[50][51], which is consistent with that in mammary glands
[51][121]. Meanwhile, DL-methionine or DL-methionine-DL-methionine supplementation to the parent diet significantly enhanced protein synthesis in crop tissue by the JAK2/STAT5 signaling pathway
[52]. In addition, the synthesis of protein in crop epidermal cells was also enhanced through activation of the IRS1/Akt/TOR signaling pathway
[53][50]. The avian prolactin receptor consists of two repeats of the extracellular ligand-binding domain. Binding of prolactin to its receptor leads to phosphorylation of one or more STAT proteins through activation of JAK
[17][52][52,55]. Then, the STAT protein is translocated into the nucleus and interacts with sites of the anxIcp35 gene and other potential genes to regulate their transcription. The anxIcp35 protein was engaged in the formation and trafficking of endocytotic vesicles (ECVs) and multivesicular bodies (MVBs), which are very important for the uptake of extracellular nutrients
[17][54][55][55,122,123]. At the same time, anxIcp35 is also involved in protein synthesis in pigeon crop
[21][56][94,124].
In the mammary gland of goats, the AMP-activated protein kinase-mammalian target of rapamycin (AMPK-mTOR) pathway participates in the sensing and utilization of amino acids
[57][125]. A total of 770 differentially expressed circRNAs (DECs) were identified in lactating and nonlactating crops. The Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis showed that the DECs were enriched in the AMPK signaling pathways
[58][126], which suggests that circRNAs may be associated with protein synthesis in crop tissue. The target genes of miRNAs that include miR-20b-5p, miR-146b-5p, miR-21-5p and miR-26b-5p were also engaged in the processes of protein synthesis, uptake and digestion
[38][110]. Furthermore, KEGG analysis of 6166 differentially expressed lncRNAs in pigeon crop showed that the biosynthesis of amino acids was significantly enriched
[59][127].
2.2.2. Synthesis of Lipid in Crop Epidermal Cells
Lipids in crop milk are regarded as an important source of energy for the growth and development of pigeon squabs. In the mammary gland of mammals, lipoprotein lipase was significantly upregulated in the de novo synthesis of triglycerides
[60][61][62][128,129,130]. Injection of prolactin into the crop of adult pigeons significantly increased the activity of lipoprotein lipase
[63][131]. In mammals, de novo lipogenesis requires the activity of key enzymes, including acetyl-CoA carboxylase (ACC) and fatty acid synthase (FAS)
[64][132]. During the peak time of lactation, the mRNA expression of ACC and FAS in pigeon crop showed high levels
[65][62]. In addition, thirty-four genes associated with lipid synthesis were differentially expressed in the lactating crop. Among them, Elovl6 was significantly upregulated in the lactating pigeon crop
[9], which has high elongation activity on C16:0 long-chain fatty acids and some activity on C18:1 and C18:2 long-chain fatty acids
[66][133].
The entry of exogenous long-chain fatty acids into epithelial cells of the mammary gland relies on special transport systems, including fatty acid translocase (FAT/CD36), fatty acid transport proteins and lipoprotein lipase, etc.
[67][68][134,135]. The expression of FAT/CD36, fatty acid-binding protein 5 (EFABP), and acyl-CoA-binding protein (ACBP) in male pigeon crop reached a maximum on Day 17 of incubation
[65][62]. However, in female crop, the mRNA expression levels of FAT/CD36, EFABP and ACBP were highest on Day 14 of incubation
[65][62], suggesting that female pigeons were prepared earlier for lipid synthesis than males. This indicates that fatty acids used for lipid biosynthesis in crop epidermal cells probably originated from exogenous supply at the terminal phases of incubation. At the same time, lipid accumulation in crop epidermal cells induced oxidative stress in mitochondria to inhibit the process of fatty acid β-oxidation, which could, in turn, increase fat deposition
[69][136].
In mammals, regulatory non-coding RNAs can regulate milk fat synthesis. For example, several important circRNAs affecting dairy milk fat synthesis were identified, including circ_0001122, circ_0007367, circ_0018269, and circ_0015179
[70][137]. Additionally, downregulation of miR-29s significantly inhibited the synthesis and secretion of triglycerides in cultured dairy cow mammary epithelial cells
[71][138]. Likewise, Ge et al. determined that miR-20b-5p, miR-146b-5p, miR-21-5p and miR-26b-5p were key miRNAs and found that the target genes of these miRNAs were engaged in lipid synthesis in pigeon crop cells
[38][110]. Although the differential expression profiles of non-coding RNAs in lactating and nonlactating pigeon crops have been revealed
[38][58][110,126], the mechanism by which these non-coding RNAs regulate lipid synthesis in crop cells remains to be further investigated.
2.2.3. Synthesis of Carbohydrates in Crop Epidermal Cells
Although carbohydrates are very important for the growth and development of birds
[72][73][139,140], their contents in pigeon milk are very low. Zhu et al.
[74][67] showed that the mRNA expression level of sodium-dependent glucose transporter 1 (SGLT1) in the crop of male and female pigeons was significantly suppressed from Day 17 of incubation to Day 7 of the chick-rearing period. Similarly, glucose transporter 2 showed the lowest mRNA expression level on Day 17 of incubation
[74][67]. AMP-activated protein kinases (AMPKs) are important cellular energy sensors that regulate glucose metabolism by promoting glucose uptake
[75][76][141,142]. During the peak of lactation, the expression levels of AMPK signaling pathway-related proteins were significantly inhibited
[74][67]. These results suggest that the process of glucose uptake by pigeon crop was severely restricted, thus resulting in a very low carbohydrate content in pigeon milk.
2.3. Shedding of Crop Epidermal Cells
Cell apoptosis is an indispensable factor that finally contributes to the shedding of crop epidermal cells to produce milk
[77][143]. As mentioned above, many nutrients, such as proteins and lipids, are synthesized in the process of crop milk formation. Excessive fat deposition in the mammalian liver leads to lipoapoptosis, fibrosis, and steatohepatitis
[78][144]. In pigeon crop cells, the oxidative stress of mitochondria was brought about by lipid overaccumulation, which ultimately results in apoptosis
[69][136]. The Ca
2+ channel plays an important role in the exchange of Ca
2+ between the endoplasmic reticulum and mitochondria
[79][145]. When unfolded or misfolded proteins accumulate excessively in cells, endoplasmic reticulum stress (ERS) occurs
[80][146]. ERS induced a large influx of Ca
2+ into mitochondria, causing its stress to evoke cell apoptosis
[81][147]. During lactation, the expression of genes related to amino acid transportation and de novo synthesis was significantly elevated in crop tissue
[43][60]. This indicates that crop milk protein is synthesized and accumulates massively in epidermal cells, which may induce ERS to stimulate their apoptosis. In addition, the expression levels of apoptosis-related genes in crop tissue were highest around Day 17 of incubation and Day 1 of the chick-rearing period
[77][143], suggesting that the crop epidermis undergoes a drastic apoptotic response during the formation of pigeon milk.