Aging is the slowest process in a living organism. During this process, mortality rate increases exponentially due to the accumulation of damage at the cellular level. Cellular senescence is a well-established hallmark of aging, as well as a promising target for preventing aging and age-related diseases. Given that the appearance of senescent cells is considered to be a cell fate transition from the proliferative state to the non-proliferative state, similar to the critical transitions that occur during cell differentiation and symptom onset, it can be detectable by the dynamical network biomarkers (DNB) theory, which detects early warning signals just before bifurcation points, such as “the pre-disease state”.
- dynamical network biomarkers theory
- aging
- DNB theory
1. Introduction
2. Dynamical Network Biomarkers Theory
2.1. The Concept of DNB Theory
In view of the bifurcation theory of dynamical systems, when organisms have a high resilience in a homeostatic state (e.g., physiologically young or healthy), a body system is stable, with fast and small fluctuations in terms of the value of a physiological parameter or gene expression. During aging with a gradual loss of resilience, on the other hand, the system enters an allostatic state [11][12], which leads to a dynamic change in the parameter that shows slow and large fluctuations. Then, the system falls into a different state via bifurcation, which is also known as the “critical transition”, and the thresholds at which this transition occurs called “tipping points” [12][13][13,14]. This other state corresponds to the disease state or the state of allostatic overload (discussed below), indicating the breakdown of a physiological network. This phenomenon is a consequence of the accumulation of undetectable elements, and once it occurs, it is often too late and non-reversible, like diabetes. Accordingly, methodology through which scholars can identify a sign of this critical transition or the state just before the critical transition, namely “the pre-disease state”, is needed for disease prediction. It is noteworthy that the concept of the pre-disease state was first proposed in the world’s oldest medical textbook, “Yellow Emperor’s Inner Canon”, in China more than 2200 years ago. In this textbook, the pre-disease state is termed “Weibing” in Chinese, which corresponds to “Mebyo”, meaning “not sick yet” in Japanese. These critical transitions are a sudden and large-scale change in state and occur in many complex systems, including ecosystems, the climate, financial markets, and microorganism populations [14][15][16][17][18][15,16,17,18,19]. The most important indicator of whether a system is getting close to the critical transition is a phenomenon known in dynamical systems theory as “critical slowing down” [12][19][13,20]. Critical slowing down exhibits increases in both fluctuation and autocorrelation, as well as a slow recovery from perturbation just before the critical transition. In general, gene expression is a fundamentally stochastic process, which stems noise from randomness in mRNA synthesis [20][23]. Stochastic gene expression results in a heterogeneous cell population that has beneficial effects in some contexts. In aging, however, noise in gene expression increases with age, and it may mask the expression changes in the genes that, in particular, lead to the critical transition towards the disease state. Notably, a data analysis using the DNB theory (namely a DNB analysis) can extract the cluster of the genes that shows large fluctuations with the strongest correlations. Thus, DNB analysis is applicable to noisy and nonlinear datasets, including multi-omics data.2.2. The Applications of DNB Theory
The DNB theory has been applied to various disease models, from cellular to organismal levels (Table 1). These include predictions of viral infection such as influenza A (H3N2) and COVID-19 [21][22][23][24,25,26], hepatocellular carcinoma metastasis [24][27], drug resistance in breast cancer [25][28], photodamage responses in skin [26][29], and adeno-to-squamous transdifferentiation in lung cancer [27][30].| Models | Cell Types or Species | Datasets | References |
|---|---|---|---|
| Influenza A (H3N2) infection | Human | Microarray of the blood samples | [21][23][24,26] |
| COVID-19 infection | Human | Case reports in five different countries and regions | [22][25] |
| Hepatocellular carcinoma | Xenograft mouse model of HCCLM3 cells | Microarray of the liver samples | [24][27] |
| Breast cancer | Human breast adenocarcinoma MCF-7 cell line | RNA-seq of MCF-7 cells | [25][28] |
| Skin photodamage | The LSE model (3D skin model consisting of normal human keratinocyte and melanocyte) | RNA-seq of the LSE model | [26][29] |
| Lung cancer | KrasLSL-G12D/+; Lkb1flox/flox (KL) mice | RNA-seq of the KL lung samples | [27][30] |
| Hematopoietic stem cell differentiation | Mouse hematopoietic stem cells (mHSCs) | scRNA-seq of mHSCs | [28][31] |
| Embryonic stem cell differentiation | Human embryonic stem cells (hESCs) | scRNA-seq of hESCs | [29][32] |
| Immune cell differentiation | T cells from DO11.10 TCR mice | Raman imaging | [30][33] |
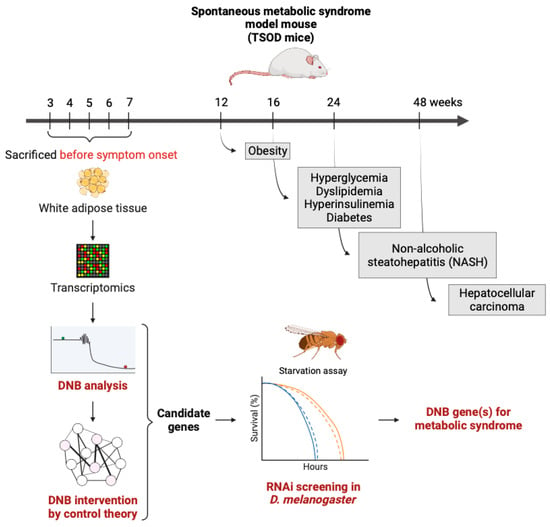
| Metabolic syndrome | Metabolic syndrome model mouse (TSOD mice) | Microarray of the adipose tissues | [31][32][34,35] |
| Type 2 diabetes | Diabetes model rat (GK rats) | Microarray of the adipose tissues | [33][36] |
2.3. Cancer and Cellular Senescence
Cellular senescence is one of the first defense mechanisms against tumor promotion during carcinogenesis. This process is known as oncogene-induced senescence (OIS), which suppress the pro-proliferative effects of oncogenic stimuli by forcing cells to become senescent and prevent the expansion of pre-cancerous cells [34][39]. Senescence is also induced by chemotherapeutic drugs or ionizing radiation during cancer treatment, termed therapy-induced senescence (TIS) [35][36][40,41]. Generally, the mechanisms underlying TIS are connected to the DNA damage response (DDR) that leads to blocking tumor cell proliferation. However, it was found that some senescent tumor cells can escape from cell cycle arrest and acquire stemness properties with a highly aggressive growth potential, which contradicts the dogma of the irreversible arrest of the cell proliferation phenotype in senescent cells [35][36][37][38][39][40,41,42,43,44]. This phenomenon is possibly characterized as “survival at the brink”, which is associated with tumor recovery and cancer relapse [39][44]. During this process, it may display large fluctuations in gene expression with strong correlations, which can be detected using the DNB theory. Notably, Jackson et al. and Huna et al. reported that the dual and heterogeneous up-regulation of two opposing regulators, p21CIP1 for senescence/apoptosis and OCT4A for stemness in the topoisomerase II inhibitor, Etoposide, induced senescent embryonal carcinoma cells [40][41][45,46], suggesting early warning signals of a cell-fate transition towards tumor recovery.3. Senolytics and Senomorphics
The emerging therapeutic strategies for targeting senescent cells are called senotherapies, which include senolytics and senomorphics. Senolytics is the selective elimination of senescent cells by small molecules, while senomorphics is the inhibition of pathological SASPs to cause senostasis (senescent cells stay there but are less harmful) [42][73]. The majority of the senolytics identified to date promote the apoptosis of senescent cells by targeting the key enzymes involved in cellular pro-survival and anti-apoptotic mechanisms, such as SRC kinases, BCL-2 family proteins, HSP90, PI3K-AKT, p53-FOXO4, GLS1, and others.
Senomorphics, on the other hand, is considered to be a safer alternative to senolytics, as it suppresses the unwanted SASP expressions from senescent cells rather than directly removing them. Senomorphics can directly or indirectly attenuate the SASP of senescent cells by inhibiting mTOR, NF-κB, SIRT1, p38MAPK, JAK-STAT, and other signaling pathways. The safety concern associated with senomorphics is the potential suppression of the growth-promoting functions induced by the SASP, similar to those seen in senolytics.
The side effects of senotherapies are due to the high heterogeneity in gene expression and the diverse origins of senescent cells, as well as their beneficial effects on tissue repair and regeneration [43][44][106,107]. An elimination of all senescent cells or a general inhibition of the SASP might cause the detrimental effects; thus, developing universal senotherapeutic drugs is extremely challenging. Further studies are needed to understand the manifestation of senescent cells for the successful development of senotherapeutic interventions. Notably, an M-DNB analysis identified the tipping points of hESC differentiation and found the master regulator genes, which commit to its cell fate determination [29][32]. Thus, scholars speculate that an M-DNB analysis may help in finding the master regulators of senescent cell development at the “pre-senescence state”, and these genes might have the potential to be novel senotherapeutic targets. If scholars could intervene in “pre-senescence cells”, it may be possible to reverse the pre-senescence to the healthy state, preventing senescent cell burden and chronic diseases, as well as delaying multimorbidity and increasing health span.4. DNB Analysis in Metabolism
4.1. Identification of DNB Genes
4.2. Verification of DNB Genes Using a Drosophila Model
In a previous study, authors identified 147 DNB genes from the white adipose tissue of the TSOD mice and determined the pre-disease state in this mouse model [31][34]. Given that the pre-disease state in each disease can be determined using the DNB theory, it is necessary to determine the means by which authors can reverse the physiological state from the pre-disease to healthy state, in order to make this approach clinically available in the near future. Therefore, authors are developing a method called DNB intervention using the control theory, especially the sample covariance matrix, which provides a list of the genes to be targeted, based on the simulation results of gene manipulations (either knockdown or activation). Consequently, authors specified 18 genes using a DNB intervention approach from the original microarray dataset, which did not have known functions relating to metabolism. To evaluate whether these genes played a role in metabolism, scholars took advantage of the fruit fly model system due to its time- and cost-effectiveness compared to mouse models. The authors performed RNAi screening using a fat body-specific (equivalent to the adipose tissue and liver in mammals) knockdown of fly orthologs of the candidate genes to observe their resistance to starvation and found several hit genes (Akagi et al., in preparation) (Figure 13). Moreover, the authors confirmed that the expressions of the mouse and human genes corresponding to the fly orthologs were slightly changed in response to high-fat diet feeding in mice or human subjects with a high body mass index, from a database analysis [47][110]. These slight changes in expression are often disregarded or impossible to detect by using the current average-based detection of the static molecular biomarkers. Using the DNB theory, however, authors can capture those “ignored genes” with a significant biological meaning. These approaches will be the model case for exploiting DNB analyses in research on aging and age-related diseases.