Breast cancer (BC) is a lethal malignancy with high morbidity and mortality but lacks effective treatments thus far. Histone deacetylases 2 (HDAC2) inhibitor (HDAC2i) has been proven to exhibit an anti-cancer effect, can act as a sensitizer for ICIsimmune checkpoint inhibitors (ICIs) therapy. Simultaneously, dietary intervention, as a crucial supportive therapy, has been reported to provide ingredients containing HDAC2 inhibitory activity. Thus, the novel integration of dietary intervention with ICIs therapy may offer promising possibilities for improving treatment outcomes.
- dietotherapy
- breast carcinoma
- HDAC2 suppression
- immune checkpoint inhibitor
1. Introduction
2. Dietary HDAC2i in Breast Cancer
2.1. HDAC2: A Potential Index of Aggressiveness and a Therapeutic Target against BC
HDAC2 is an enzyme involved in the tightening of the chromatin structure and suppression of gene transcription. Evidence has indicated that HDAC2 is overexpressed in breast cancer cells compared to normal breast tissue. Higher levels of HDAC2 have also been associated with more aggressive tumor characteristics, such as increased cell proliferation, invasion, and metastasis [12,13,14,15][12][13][14][15]. For instance, a clinical study that included 226 BC patients reported that expression of HDAC2 protein is significantly higher in breast cancer than in benign tumors and indicates that HDAC2 may be involved in invasion, metastasis, anthracyclines therapy resistance, and poor prognosis of sporadic breast cancer.2.2. HDAC2 Inhibition for Treating Breast Cancer
HDAC2 has been identified as a crucial regulator of epigenetic control in BC and HDAC2 suppression has been further proved to be an effective approach to treating BC by numerous studies [24,25,26][21][22][23]. For instance, HDAC2 inhibition has been observed to inhibit cellular proliferation in a p53-dependent manner in BC cells [27][24]. MiR-155 can also decrease the expression of erythroblastic oncogene B by targeting HDAC2 [28][25]. Moreover, evidence has indicated that PELP1 (proline, glutamate, and leucine-rich protein 1) can bind to miR-200a and miR-141 promoter sequences and modulate the expression of these miRNAs by recruiting HDAC2; therefore, regulating tumorigenic and metastatic potential of BC cells [29][26]. Thus, a molecular network involving HDAC2 has been considered to serve as a target for developing anti-cancer drugs [29][26]. Inhibition of HDAC2 has also been found to exert a synergistic effect in BC treatment. For instance, evidence indicates that depleting HDAC2 can sensitize breast cancer cells to apoptosis induced by epirubicin. This finding highlighted the potential of HDAC2 as a therapeutic target and a biomarker in treating breast cancer [30][27]. Moreover, the modulation of estrogen receptor (ER) signaling is a promising therapeutic approach in ER-expressing breast cancers, and the progesterone receptor (PR) also plays a critical role in this process [31][28].2.3. HDAC2 Inhibition Enhances the Therapeutic Effect of ICIs in BC Treatment
The emergence of immune checkpoint inhibitors (ICI) has revolutionized the treatment of breast cancer [33,34,35,36,37,38,39,40][29][30][31][32][33][34][35][36]. Adjuvant or neoadjuvant immune checkpoint blockades are used for metastatic breast cancer [34][30]. Despite ICI therapy being promising, breast cancer cells often find ways to evade the host’s immune system, necessitating combination therapies to overcome these limitations. HDAC inhibitors (HDACi) have demonstrated potent immunomodulatory activity, making them a rational choice for cancer immunotherapies.
2.3.1. HDAC2 Regulates PD-L1 Nuclear Translocation
As a ligand of PD-1, high PD-L1 levels indicate tumor progression and are associated with poor prognosis in immunotherapy-treated human cancer [35][31]. PD-L1 nuclear translocation has been identified as a key mechanism underlying the immune evasion of BC cells, hindering PD-1 inhibitors [36][32]. Both cytoplasmic and nuclear PD-L1 can exert immunosuppressive functions on BC cells [37][33]. Nuclear PD-L1 has been linked to various cellular processes, for example, increasing the anti-apoptotic capacity of tumor cells, promoting mTOR activity, and upregulating glycolytic metabolism [36][32]. A study has shown that the level of nuclear PD-L1 expression was positively correlated with immune response-related transcription factors, such as STAT3, RelA (p65), and c-Jun [19]. Nuclear PD-L1 interacts with transcription factors, such as RelA and the IFN regulatory factor (IRF), influencing antitumor immunity. Inhibition of nuclear PD-L1 expression led to downregulation of genes involved in evading immune surveillance, such as PDCD1LG2 (encoding PD-L2), VSIR (encoding VISTA), and CD276 (encoding B7-H3), which enhance cytotoxic T-lymphocyte depletion and promote tumor aggressiveness, distant metastasis, and resistance to PD-L1/PD-1 blockade therapy [18,19,38][18][19][34] (Figure 21).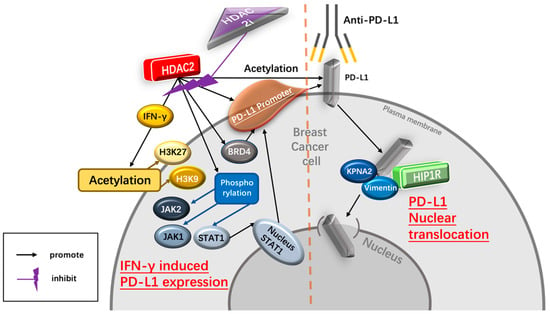
2.3.2. HDAC2 Regulates IFN-γ- Induced PD-L1 Expression
IFN-γ upregulates the expression of PD-L1 Interferon-γ (IFN-γ) is a crucial cytokine in both innate and adaptive immunity and should be considered as an important driving force for PD-L1 expression in tumor microenvironment. It is able to induce PD-L1 expression on BC cells and increase the apoptosis of antigen-specific T cells, such a process is referred to as “adaptive resistance” [43][37]. Depleting IFN-γ receptor 1 has been reported to decrease the expression of PD-L1 expression in BC cells, increase the amount of tumor-infiltrating CD8+ lymphocytes or CTLs, and to inhibit the development of BC cells [44][38]. HDAC2i affects IFN-γ induced PD-L1 HDAC2 can promote PD-L1 induced via IFN-γ stimulation in BC cells [18,19][18][19]. Specifically, IFN-γ can induce gene transcription involving STAT1 binding to the gamma interferon activation site (GAS), recruiting HAT and HDAC for chromatin remodeling [45][39]. Following IFN-γ stimulation, BRD4 is rapidly recruited to the PD-L1 locus, accompanied by increased H3K27ac and RNA Polymerase II (RNA Pol II) occupancy in cancer cells [45][39]. A ChIP-qPCR assay confirmed enhanced HDAC2 binding to the PD-L1 promoter after IFN-γ treatment. HDAC2 knockout led to reduced STAT1 occupancy and bromodomain-containing 4 (BRD4) recruitment to the PD-L1 promoter, attenuated H3K27ac and H3K9ac (markers of active transcription in the PD-L1 promoter) upregulation induced by IFN-γ, highlighting HDAC2’s role in activating PD-L1 expression through IFN-γ induced signaling pathways [18].2.4. Dietary Intervention Is Important for Breast Cancer Patients Receiving Anti-Cancer Immunotherapy
A healthy diet rich in vitamins, minerals, antioxidants, and phytochemicals, provides essential ingredients for building and strengthening a healthy immune system. Thus, the American Cancer Society (ACS) releases the Nutrition and Physical Activity Guideline for Cancer Survivors, which enhances immune function by supporting essential nutrients [47[40][41][42][43],48,49,50], helping to optimize the effectiveness of immunotherapy and improve treatment outcomes. Moreover, anti-cancer immunotherapy also possesses toxicities and can lead to various adverse effects on breast cancer patients, such as fatigue, nausea, loss of appetite, and gastrointestinal issues [51][44]. However, proper dietary interventions have been found to help alleviate these side effects by providing proper nutrition, promoting hydration, and supporting gastrointestinal health [52,53,54][45][46][47].2.5. Dietary HDAC2i
Compared to commonly used anti-cancer therapies, dietary interventions are safer and more cost-effective [65,66][48][49]. Even if dietary interventions cannot be considered a replacement for conventional cancer treatments, their role in improving the outcomes of cancer treatment also cannot be ignored [67,68][50][51] (Figure 32).
2.6. Selected Candidates of HDAC2i
2.6.1. Genistein (GE)
GE is the most predominant bioactive isoflavone found mainly in soybean products and other food sources such as lupin, fava beans, kudzu, and psoralea [84][68]. GE has been reported to act as a potent chemo-preventive and therapeutic agent against various types of cancers including breast, prostate, and lung cancer [85][69]. For instance, GE suppresses the growth of BC cells in patient-derived tumor xenograft (PDX) [85][69]. Moreover, GE has been reported to modify the expression levels and activities of key epigenetic-associated genes, including HDAC2, DNA methyltransferases (DNMT3b) and ten-eleven translocation (TET3) methylcytosine dioxygenases. These genes are involved in epigenetic modifications, such as DNA methylation and histone methylation, which can regulate gene expression and impact cellular behavior [85][69]. By modulating the activities of these epigenetic regulators, GE may influence the epigenetic landscape of breast cancer cells, leading to changes in gene expression patterns that can affect cancer-related pathways.2.6.2. Sulforaphane (SFN)
SFN is a natural compound and is abundant in cruciferous vegetables such as broccoli sprouts (BSp) and kale [111][70]. It has gained attention for its potential health benefits, including its ability to inhibit HDAC2 activity [112,113][71][72]. In the context of breast cancer, a dietary regimen of genistein and BSp in combination has been shown effective in reducing mammary tumor incidence and delaying tumor latency in a spontaneous breast cancer mouse model [86][73]. The combination of GE and SFN downregulated HDAC2 protein levels in breast cancer cells.2.6.3. Chrysin and Its Analogues
Chrysin and its analogues are a group of polyphenolic compounds found in various dietary sources such as fruits, vegetables, olive oil, tea, and red wine [114][74]. The cytotoxic effects of chrysin have been shown against a wide range of cancer cell lines, including BC (MCF-7, MDA-MB-231), colon cancer (Lovo, DLD-1), and prostate cancer cells [87,88][75][76]. It is able to induce G1 cell cycle arrest and inhibit the activity of HDACs, specifically HDAC2 [89][77].2.6.4. Resveratrol (RSV)
RSV is also a polyphenol abundant in grape skin and seeds. It also presents in other food sources such as apples, blueberries, mulberries, peanuts, pistachios, plums, and red wine [90][78]. RSV has numerous beneficial properties of anti-glycosylation, anti-inflammation, anti-neurodegeneration, and antioxidation in various types of cancer [91][79]. One intriguing aspect of RSV is its proposed potential as a pan-HDAC inhibitor [92][80].2.6.5. Oleuropein (OLE)
OLE is a polyphenolic compound in virgin olive oil with antineoplastic properties and it is well tolerated by humans [94][81]. Studies have shown that OLE can reduce progression, invasion, and proliferation of breast cancer cells by suppressing the activity of both HDAC2 and HDAC3 [95][82]. However, OLE exhibits little negative effect on normal breast epithelial cells, suggesting a potential selectivity towards BC cells and its potential for BC patients receiving ICIs therapy [95][82].2.6.6. Curcumin
Curcumin, a lipophilic polyphenol derived from turmeric (Curcuma longa), has been extensively studied for its diverse health-promoting properties, including antioxidant, anti-inflammatory, hepatoprotective, anti-atherosclerotic, and antidiabetic effects [96][83].2.6.7. Valeric Acid
Valerian (Valeriana officinalis) is a medicated diet that has been commonly used in cooking soup by some ethnic minorities in China for hundreds of years for restoring and balancing body energy [98][84]. Valeric acid, a major active component of valerian, has been identified as a potential HDAC inhibitor with anti-cancer effects on liver and breast cancer [98][84].2.6.8. Rh4
Ginseng is also a typical medicated diet item and is commonly used for making cuisine mainly in Asia. It is also a traditional Chinese herb with multiple biological effects. One of its components, Rh4, has been identified as a rare ginsenoside with potential inhibitive effects on the development of various cancers [100][85]. Rh4 can inhibit the expression of PD-L1 by regulating HDAC2-mediated JAK/STAT in breast cancer cells [101][86].2.6.9. Butyrate (NaB)
NaB, a short-chain fatty acid generated via the fermentation of dietary fiber by the colonic microbiota, has shown anticancer activities mediated through HDACi [102][87]. NaB is primarily derived from undigested dietary carbohydrates, such as resistant starch and dietary fiber and, to a lesser extent, from dietary and endogenous proteins [103,104][88][89]. Studies have demonstrated that treatment with NaB, when combined with retinoids, enhances the inhibition of breast cancer cell proliferation [105][90].2.6.10. Other Potential Candidates
Some other dietary compounds have also been identified as HDAC2i in other cancer types. For instance, green tea and its bioactive components, especially polyphenols, possess many health-promoting and disease-preventing benefits with anti-inflammatory, antimutagenic, antioxidant, and anticancer properties, but have no significant toxicity on normal cells in vivo. It has the potential as an effective chemotherapeutic agent for cancer prevention and treatment through various cellular, molecular, and biochemical mechanisms [116][91]. The major polyphenol components of green tea are (-)-epigallocatechin-3-gallate (EGCG), (-)-epigallocatechin (EGC), (-)-epicatechin-3-gallate (ECG) and (-)-epicatechin (EC) [116][91]. One of the molecular mechanisms underlying the anticancer effects of green tea polyphenols (GTPs) is HDAC2 inhibition [107][92].2.7. Potential Approaches of Taking Bioactive Compound
For enhancing absorption efficiency and therefore improving the potential health benefits and biological activities of certain bioactive compounds, many means of application have been developed [119][93]. Some of them might provide a better way for BC patients receiving ICIs to gain HDAC2i efficiently. However, it is important to emphasize that thorough exploration in this area is still a pressing necessity. Nutraceuticals and Dietary Supplements Many bioactive compounds with antioxidant or anti-inflammatory properties, found in certain fruits or vegetables, have been produced as nutraceuticals and dietary supplements for taking them more conveniently and easily [65][48]. Moreover, nutraceuticals and dietary supplements have been found to maintain excellent safety levels [120][94]. Nanotechnology and Drug Delivery Bioactive compounds can also be incorporated into well-designed nanoparticles for targeted drug delivery, enhancing drug efficacy and reducing side effects [122][95]. For instance, theracurmin, a curcumin formulation consisting of dispersed curcumin with colloidal nanoparticles, possesses significantly improved bioavailability and therapeutic efficacy for treating osteoarthritis, compared to turmeric powder monotherapy [123,124,125][96][97][98]. Pharmaceuticals and Medicinal Products Some bioactive compounds can be isolated and developed into pharmaceutical drugs to efficiently improve their therapeutic effect [129][99]. For instance, curcumin, a bioactive compound that has been found to possess multiple biological regulatory functions, has been successfully isolated from plant curcuma aromatica salisb for treating different types of cancer, including BC [130,131][100][101]. Phytotherapy and Traditional Medicine Phytotherapy and traditional medicine have been widely applied in treating various of diseases [133,134][102][103]. They are natural, with relatively low irritation and side effects on the human body, and can also be utilized in combination with other treatment [135,136][104][105].2.8. Nutrients That May Impair the Therapeutic Effect of ICIs
2.8. Nutrients That May Impair the Therapeutic Effect of ICIs
Omega-3 Fatty Acids Omega-3 fatty acids, commonly present in fish oil and certain plant sources, possess anti-inflammatory properties and are essential for synthesizing hormones and endogenous substances [139][106]. Natural killer (NK) cells are innate lymphocytes responsible for orchestrating immune responses against tumors and viruses [140][107]. Fish oil supplementation was found to decrease NK cell activity, which rebounded after supplementation ceased [141][108]. Vitamins Vitamins are a type of trace organic substance obtained from food that can maintain normal physiological functions in humans [142][109]. Vitamins participate in the biochemical reactions of the human body and regulate metabolic functions, including immunity [143][110]. Deficiency or over intake of certain vitamins has been found to impair anti-cancer immunity, therefore affecting the efficiency of ICIs [138][111]. Probiotics Probiotics, including bacteria and yeast, are living microorganisms [149,150][112][113]. Some of them have been commonly utilized to promote gut health, closely intertwined with immune function [149,150][112][113]. Recent evidence has newly pointed out that an excessive immune response in the gut induced by overconsumption of probiotics might constrain the systemic immune reaction necessary for the optimal efficacy of ICIs [151,152][114][115]. High-Fiber Diets Fiber-rich diets primarily comprise two essential elements: soluble fiber and insoluble fiber. These vital components are found in an array of plant-based foods, including legumes, whole grains, cereals, vegetables, fruits, nuts, and seeds. Dietary fiber is composed of non-starch polysaccharides and various plant constituents like cellulose, resistant starch, and resistant dextrins [153][116]. High-fiber diets have been considered to modulate the gut microbiota and influence immune responses [154][117]. While a diverse gut microbiome is generally associated with better health, certain bacterial metabolites produced from high-fiber diets could potentially hamper the efficiency of ICIs [155][118]. Ketogenic diet The ketogenic diet (KD) is characterized by high fat, low to moderate protein, and very low carbohydrate intake [156][119]. Evidence has shown that KD can lead to a downregulation of CTLA-4 and PD-1 expression on tumor-infiltrating lymphocytes (TILs), as well as PD-L1 expression on glioblastoma cells in animal models [157][120]. Protein-restricted diet A low-protein diet serves as a therapeutic approach for managing inherited metabolic disorders like phenylketonuria and homocystinuria. Additionally, it can be employed in the treatment of kidney or liver ailments. Furthermore, a reduced intake of protein has been observed to potentially lower the risk of bone fractures, likely due to alterations in calcium [159][121].References
- Tao, X.; Li, T.; Gandomkar, Z.; Brennan, P.C.; Reed, W.M. Incidence, mortality, survival, and disease burden of breast cancer in China compared to other developed countries. Asia-Pac. J. Clin. Oncol. 2023.
- Bazzi, T.; Al-Husseini, M.; Saravolatz, L.; Kafri, Z. Trends in Breast Cancer Incidence and Mortality in the United States From 2004-2018: A Surveillance, Epidemiology, and End Results (SEER)-Based Study. Cureus 2023, 15, e37982.
- Pedersini, R.; di Mauro, P.; Bosio, S.; Zanini, B.; Zanini, A.; Amoroso, V.; Turla, A.; Vassalli, L.; Ardine, M.; Monteverdi, S.; et al. Changes in eating habits and food preferences in breast cancer patients undergoing adjuvant chemotherapy. Sci. Rep. 2021, 11, 12975.
- Taylor, C.; McGale, P.; Probert, J.; Broggio, J.; Charman, J.; Darby, S.C.; Kerr, A.J.; Whelan, T.; Cutter, D.J.; Mannu, G.; et al. Breast cancer mortality in 500 000 women with early invasive breast cancer in England, 1993–2015: Population based observational cohort study. BMJ 2023, 381, e074684.
- Bertucci, F.; Gonçalves, A. Immunotherapy in Breast Cancer: The Emerging Role of PD-1 and PD-L1. Curr. Oncol. Rep. 2017, 19, 64.
- Lu, L.; Bai, Y.; Wang, Z. Elevated T cell activation score is associated with improved survival of breast cancer. Breast Cancer Res. Treat. 2017, 164, 689–696.
- Jacob, S.L.; Huppert, L.A.; Rugo, H.S. Role of Immunotherapy in Breast Cancer. JCO Oncol. Pract. 2023, 19, 167–179.
- Hattori, M.; Masuda, N.; Takano, T.; Tsugawa, K.; Inoue, K.; Matsumoto, K.; Ishikawa, T.; Itoh, M.; Yasojima, H.; Tanabe, Y.; et al. Pembrolizumab plus chemotherapy in Japanese patients with triple-negative breast cancer: Results from KEYNOTE-355. Cancer Med. 2023, 12, 10280–10293.
- Downs-Canner, S.; Mittendorf, E.A. Correction: Preoperative Immunotherapy Combined with Chemotherapy for Triple-Negative Breast Cancer: Perspective on the KEYNOTE-522 Study. Ann. Surg. Oncol. 2023, 30, 3286.
- Khalid, A.B.; Calderon, G.; Jalal, S.I.; Durm, G.A. Physician Awareness of Immune-Related Adverse Events of Immune Checkpoint Inhibitors. Breast Cancer Res. Treat 2022, 20, 1316–1320.
- Gumusay, O.; Callan, J.; Rugo, H.S. Immunotherapy toxicity: Identification and management. Breast Cancer Res. Treat. 2022, 192, 1–17.
- Liu, S.; Zhao, S.; Dong, Y.; Wang, T.; Niu, X.; Zhao, L.; Wang, G. Antitumor activity and mechanism of resistance of the novel HDAC and PI3K dual inhibitor CUDC-907 in pancreatic cancer. Cancer Chemother. Pharmacol. 2021, 87, 415–423.
- Garmpis, N.; Damaskos, C.; Dimitroulis, D.; Kouraklis, G.; Garmpi, A.; Sarantis, P.; Koustas, E.; Patsouras, A.; Psilopatis, I.; Antoniou, E.A.; et al. Clinical Significance of the Histone Deacetylase 2 (HDAC-2) Expression in Human Breast Cancer. J. Pers. Med. 2022, 12, 1672.
- Shan, W.; Jiang, Y.; Yu, H.; Huang, Q.; Liu, L.; Guo, X.; Li, L.; Mi, Q.; Zhang, K.; Yang, Z. HDAC2 overexpression correlates with aggressive clinicopathological features and DNA-damage response pathway of breast cancer. Am. J. Cancer Res. 2017, 7, 1213–1226.
- Zhao, H.; Yu, Z.; Zhao, L.; He, M.; Ren, J.; Wu, H.; Chen, Q.; Yao, W.; Wei, M. HDAC2 overexpression is a poor prognostic factor of breast cancer patients with increased multidrug resistance-associated protein expression who received anthracyclines therapy. Jpn. J. Clin. Oncol. 2016, 46, 893–902.
- Maccallini, C.; Ammazzalorso, A.; De Filippis, B.; Fantacuzzi, M.; Giampietro, L.; Amoroso, R. HDAC Inhibitors for the Therapy of Triple Negative Breast Cancer. Pharmaceuticals 2022, 15, 667.
- Li, Y.; Seto, E. HDACs and HDAC Inhibitors in Cancer Development and Therapy. Cold Spring Harb. Perspect. Med. 2016, 6, a026831.
- Xu, P.; Xiong, W.; Lin, Y.; Fan, L.; Pan, H.; Li, Y. Histone deacetylase 2 knockout suppresses immune escape of triple-negative breast cancer cells via downregulating PD-L1 expression. Cell Death Dis. 2021, 12, 779.
- Gao, Y.; Nihira, N.T.; Bu, X.; Chu, C.; Zhang, J.; Kolodziejczyk, A.; Fan, Y.; Chan, N.T.; Ma, L.; Liu, J.; et al. Acetylation-dependent regulation of PD-L1 nuclear translocation dictates the efficacy of anti-PD-1 immunotherapy. Nat. Cell Biol. 2020, 22, 1064–1075.
- Bassett, S.A.; Barnett, M.P.G. The Role of Dietary Histone Deacetylases (HDACs) Inhibitors in Health and Disease. Nutrients 2014, 6, 4273–4301.
- Muller, B.M.; Jana, L.; Kasajima, A.; Lehmann, A.; Prinzler, J.; Budczies, J.; Winzer, K.-J.; Dietel, M.; Weichert, W.; Denkert, C. Differential expression of histone deacetylases HDAC1, 2 and 3 in human breast cancer—Overexpression of HDAC2 and HDAC3 is associated with clinicopathological indicators of disease progression. BMC Cancer 2013, 13, 215.
- Long, M.; Hou, W.; Liu, Y.; Hu, T. A Histone Acetylation Modulator Gene Signature for Classification and Prognosis of Breast Cancer. Curr. Oncol. 2021, 28, 928–939.
- Choi, S.R.; Hwang, C.Y.; Lee, J.; Cho, K.-H. Network Analysis Identifies Regulators of Basal-Like Breast Cancer Reprogramming and Endocrine Therapy Vulnerability. Cancer Res. 2022, 82, 320–333.
- Harms, K.L.; Chen, X. Histone Deacetylase 2 Modulates p53 Transcriptional Activities through Regulation of p53-DNA Binding Activity. Cancer Res. 2007, 67, 3145–3152.
- He, X.-H.; Zhu, W.; Yuan, P.; Jiang, S.; Li, D.; Zhang, H.-W.; Liu, M.-F. miR-155 downregulates ErbB2 and suppresses ErbB2-induced malignant transformation of breast epithelial cells. Oncogene 2016, 35, 6015–6025.
- Jo, H.; Shim, K.; Kim, H.-U.; Jung, H.S.; Jeoung, D. HDAC2 as a target for developing anti-cancer drugs. Comput. Struct. Biotechnol. J. 2023, 21, 2048–2057.
- Biçaku, E.; Marchion, D.C.; Schmitt, M.L.; Münster, P.N. Selective Inhibition of Histone Deacetylase 2 Silences Progesterone Receptor–Mediated Signaling. Cancer Res. 2008, 68, 1513–1519.
- Marchion, D.C.; Bicaku, E.; Turner, J.G.; Schmitt, M.L.; Morelli, D.R.; Munster, P.N. HDAC2 regulates chromatin plasticity and enhances DNA vulnerability. Mol. Cancer Ther. 2009, 8, 794–801.
- Masoumi, E.; Tahaghoghi-Hajghorbani, S.; Jafarzadeh, L.; Sanaei, M.-J.; Pourbagheri-Sigaroodi, A.; Bashash, D. The application of immune checkpoint blockade in breast cancer and the emerging role of nanoparticle. J. Control. Release 2021, 340, 168–187.
- Isaacs, J.; Anders, C.; McArthur, H.; Force, J. Biomarkers of Immune Checkpoint Blockade Response in Triple-Negative Breast Cancer. Curr. Treat. Options Oncol. 2021, 22, 38.
- Lu, L.; Risch, E.; Halaban, R.; Zhen, P.; Bacchiocchi, A.; Risch, H.A. Dynamic changes of circulating soluble PD-1/PD-L1 and its association with patient survival in immune checkpoint blockade-treated melanoma. Int. Immunopharmacol. 2023, 118, 110092.
- Fan, Z.; Wu, C.; Chen, M.; Jiang, Y.; Wu, Y.; Mao, R.; Fan, Y. The generation of PD-L1 and PD-L2 in cancer cells: From nuclear chromatin reorganization to extracellular presentation. Acta Pharm. Sin. B 2022, 12, 1041–1053.
- Xiong, W.; Gao, Y.; Wei, W.; Zhang, J. Extracellular and nuclear PD-L1 in modulating cancer immunotherapy. Trends Cancer 2021, 7, 837–846.
- Koh, Y.W.; Han, J.-H.; Haam, S.; Lee, H.W. HIP1R Expression and Its Association with PD-1 Pathway Blockade Response in Refractory Advanced NonSmall Cell Lung Cancer: A Gene Set Enrichment Analysis. J. Clin. Med. 2020, 9, 1425.
- Muenst, S.; Schaerli, A.R.; Gao, F.; Däster, S.; Trella, E.; Droeser, R.A.; Muraro, M.G.; Zajac, P.; Zanetti, R.; Gillanders, W.E.; et al. Expression of programmed death ligand 1 (PD-L1) is associated with poor prognosis in human breast cancer. Breast Cancer Res. Treat. 2014, 146, 15–24.
- Lotfinejad, P.; Kazemi, T.; Safaei, S.; Amini, M.; Baghbani, E.; Shotorbani, S.S.; Niaragh, F.J.; Derakhshani, A.; Shadbad, M.A.; Silvestris, N. PD-L1 silencing inhibits triple-negative breast cancer development and upregulates T-cell-induced pro-inflammatory cytokines. Biomed. Pharmacother. 2021, 138, 111436.
- Minn, A.J.; Wherry, E.J. Combination Cancer Therapies with Immune Checkpoint Blockade: Convergence on Interferon Signaling. Cell 2016, 165, 272–275.
- Abiko, K.; Matsumura, N.; Hamanishi, J.; Horikawa, N.; Murakami, R.; Yamaguchi, K.; Yoshioka, Y.; Baba, T.; Konishi, I.; Mandai, M. IFN-γ from lymphocytes induces PD-L1 expression and promotes progression of ovarian cancer. Br. J. Cancer 2015, 112, 1501–1509.
- Bouhet, S.; Lafont, V.; Billard, E.; Gross, A.; Dornand, J. The IFNgamma-induced STAT1-CBP/P300 association, required for a normal response to the cytokine, is disrupted in Brucella-infected macrophages. Microb. Pathog. 2009, 46, 88–97.
- Calder, P.C. Foods to deliver immune-supporting nutrients. Curr. Opin. Food Sci. 2022, 43, 136–145.
- Calder, P.C.; Carr, A.G.; Gombart, A.F.; Eggersdorfer, M. Reply to Comment on: Optimal Nutritional Status for a Well-Functioning Immune System Is an Important Factor to Protect against Viral Infections. Nutrients 2020, 12, 1181.
- Chen, O.; Mah, E.; Dioum, E.; Marwaha, A.; Shanmugam, S.; Malleshi, N.; Sudha, V.; Gayathri, R.; Unnikrishnan, R.; Anjana, R.M.; et al. The Role of Oat Nutrients in the Immune System: A Narrative Review. Nutrients 2021, 13, 1048.
- Noor, S.; Piscopo, S.; Gasmi, A. Nutrients Interaction with the Immune System. Arch. Razi. Inst. 2021, 76, 1579–1588.
- Kichloo, A.; Albosta, M.; Dahiya, D.; Guidi, J.C.; Aljadah, M.; Singh, J.; Shaka, H.; Wani, F.; Kumar, A.; Lekkala, M. Systemic adverse effects and toxicities associated with immunotherapy: A review. World J. Clin. Oncol. 2021, 12, 150–163.
- Ticinesi, A.; Nouvenne, A.; Chiussi, G.; Castaldo, G.; Guerra, A.; Meschi, T. Calcium Oxalate Nephrolithiasis and Gut Microbiota: Not just a Gut-Kidney Axis. A Nutritional Perspective. Nutrients 2020, 12, 548.
- Taraszewska, A. Risk factors for gastroesophageal reflux disease symptoms related to lifestyle and diet. Rocz Panstw Zakl Hig 2021, 72, 21–28.
- Jadhav, A.; Bajaj, A.; Xiao, Y.; Markandey, M.; Ahuja, V.; Kashyap, P.C. Role of Diet–Microbiome Interaction in Gastrointestinal Disorders and Strategies to Modulate Them with Microbiome-Targeted Therapies. Annu. Rev. Nutr. 2023, 43, 355–383.
- Martínez-Garay, C.; Djouder, N. Dietary interventions and precision nutrition in cancer therapy. Trends Mol. Med. 2023, 29, 489–511.
- Vernieri, C.; Casola, S.; Foiani, M.; Pietrantonio, F.; De Braud, F.; Longo, V. Targeting Cancer Metabolism: Dietary and Pharmacologic Interventions. Cancer Discov. 2016, 6, 1315–1333.
- Bose, S.; Allen, A.E.; Locasale, J.W. The Molecular Link from Diet to Cancer Cell Metabolism. Mol. Cell 2020, 78, 1034–1044.
- Locasale, J.W. Diet and Exercise in Cancer Metabolism. Cancer Discov. 2022, 12, 2249–2257.
- Morita, M.; Kudo, K.; Shima, H.; Tanuma, N. Dietary intervention as a therapeutic for cancer. Cancer Sci. 2021, 112, 498–504.
- Patra, S.; Pradhan, B.; Nayak, R.; Behera, C.; Das, S.; Patra, S.K.; Efferth, T.; Jena, M.; Bhutia, S.K. Dietary polyphenols in chemoprevention and synergistic effect in cancer: Clinical evidences and molecular mechanisms of action. Phytomedicine 2021, 90, 153554.
- Zhou, Y.; Li, H. Neurological adverse events associated with PD-1/PD-L1 immune checkpoint inhibitors. Front. Neurosci. 2023, 17, 1227049.
- Alturki, N.A. Review of the Immune Checkpoint Inhibitors in the Context of Cancer Treatment. J. Clin. Med. 2023, 12, 4301.
- Zraik, I.M.; Heß-Busch, Y. Management of chemotherapy side effects and their long-term sequelae. Urologe A 2021, 60, 862–871.
- Buchholz, T.A. Radiation Therapy for Early-Stage Breast Cancer after Breast-Conserving Surgery. New Engl. J. Med. 2009, 360, 63–70.
- Zhang, C.; Xu, C.; Gao, X.; Yao, Q. Platinum-based drugs for cancer therapy and anti-tumor strategies. Theranostics 2022, 12, 2115–2132.
- Wu, Q.; Gao, Z.-J.; Yu, X.; Wang, P. Dietary regulation in health and disease. Signal Transduct. Target. Ther. 2022, 7, 252.
- Prado, C.M.; Antoun, S.; Sawyer, M.B.; Baracos, V.E. Two faces of drug therapy in cancer: Drug-related lean tissue loss and its adverse consequences to survival and toxicity. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 250–254.
- Soldati, L.; Di Renzo, L.; Jirillo, E.; Ascierto, P.A.; Marincola, F.M.; De Lorenzo, A. The influence of diet on anti-cancer immune responsiveness. J. Transl. Med. 2018, 16, 75.
- Greathouse, K.L.; Wyatt, M.; Johnson, A.J.; Toy, E.P.; Khan, J.M.; Dunn, K.; Clegg, D.J.; Reddy, S. Diet-microbiome interactions in cancer treatment: Opportunities and challenges for precision nutrition in cancer. Neoplasia 2022, 29, 100800.
- Tao, J.; Li, S.; Gan, R.-Y.; Zhao, C.-N.; Meng, X.; Li, H.-B. Targeting gut microbiota with dietary components on cancer: Effects and potential mechanisms of action. Crit. Rev. Food Sci. Nutr. 2020, 60, 1025–1037.
- Dieli-Conwright, C.M.; Harrigan, M.; Cartmel, B.; Chagpar, A.; Bai, Y.; Li, F.-Y.; Rimm, D.L.; Pusztai, L.; Lu, L.; Sanft, T.; et al. Impact of a randomized weight loss trial on breast tissue markers in breast cancer survivors. npj Breast Cancer 2022, 8, 29.
- Puklin, L.; Cartmel, B.; Harrigan, M.; Lu, L.; Li, F.-Y.; Sanft, T.; Irwin, M.L. Randomized trial of weight loss on circulating ghrelin levels among breast cancer survivors. npj Breast Cancer 2021, 7, 49.
- Thomas, G.A.; Alvarez-Reeves, M.; Lu, L.; Yu, H.; Irwin, M.L. Effect of Exercise on Metabolic Syndrome Variables in Breast Cancer Survivors. Int. J. Endocrinol. 2013, 2013, 168797.
- Zeng, H.; Irwin, M.L.; Lu, L.; Risch, H.; Mayne, S.; Mu, L.; Deng, Q.; Scarampi, L.; Mitidieri, M.; Katsaros, D.; et al. Physical activity and breast cancer survival: An epigenetic link through reduced methylation of a tumor suppressor gene L3MBTL1. Breast Cancer Res. Treat. 2012, 133, 127–135.
- Garbiec, E.; Cielecka-Piontek, J.; Kowalówka, M.; Hołubiec, M.; Zalewski, P. Genistein—Opportunities Related to an Interesting Molecule of Natural Origin. Molecules 2022, 27, 815.
- Sharma, M.; Arora, I.; Chen, M.; Wu, H.; Crowley, M.R.; Tollefsbol, T.O.; Li, Y. Therapeutic Effects of Dietary Soybean Genistein on Triple-Negative Breast Cancer via Regulation of Epigenetic Mechanisms. Nutrients 2021, 13, 3944.
- Çakır, I.; Pan, P.L.; Hadley, C.K.; El-Gamal, A.; Fadel, A.; Elsayegh, D.; Mohamed, O.; Rizk, N.M. Is a corresponding author Masoud Ghamari-Langroudi. Sulforaphane reduces obesity by reversing leptin resistance. Elife 2022, 11, e67368.
- Xie, Q.; Bai, Q.; Zou, L.-Y.; Zhang, Q.-Y.; Zhou, Y.; Chang, H.; Yi, L.; Zhu, J.-D.; Mi, M.-T. Genistein inhibits DNA methylation and increases expression of tumor suppressor genes in human breast cancer cells. Genes Chromosom. Cancer 2014, 53, 422–431.
- Tortorella, S.M.; Royce, S.G.; Licciardi, P.V.; Karagiannis, T.C. Dietary Sulforaphane in Cancer Chemoprevention: The Role of Epigenetic Regulation and HDAC Inhibition. Antioxid. Redox Signal. 2015, 22, 1382–1424.
- Paul, B.; Li, Y.; Tollefsbol, T.O. The Effects of Combinatorial Genistein and Sulforaphane in Breast Tumor Inhibition: Role in Epigenetic Regulation. Int. J. Mol. Sci. 2018, 19, 1754.
- Middleton, E., Jr.; Kandaswami, C.; Theoharides, T.C. The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharmacol. Rev. 2000, 52, 673–751.
- Chang, H.; Mi, M.; Ling, W.; Zhu, J.; Zhang, Q.; Wei, N.; Zhou, Y.; Tang, Y.; Yuan, J. Structurally related cytotoxic effects of flavonoids on human cancer cells in vitro. Arch. Pharmacal Res. 2008, 31, 1137–1144.
- Androutsopoulos, V.; Papakyriakou, A.; Vourloumis, D.; Spandidos, D.A. Comparative CYP1A1 and CYP1B1 substrate and inhibitor profile of dietary flavonoids. Bioorg. Med. Chem. 2011, 19, 2842–2849.
- Pal-Bhadra, M.; Ramaiah, M.J.; Reddy, T.L.; Krishnan, A.; Pushpavalli, S.; Babu, K.S.; Tiwari, A.K.; Rao, J.M.; Yadav, J.S.; Bhadra, U. Plant HDAC inhibitor chrysin arrest cell growth and induce p21 WAF1 by altering chromatin of STAT response element in A375 cells. BMC Cancer 2012, 12, 180.
- Sedlak, L.; Wojnar, W.; Zych, M.; Wyględowska-Promieńska, D.; Mrukwa-Kominek, E.; Kaczmarczyk-Sedlak, I. Effect of Resveratrol, a Dietary-Derived Polyphenol, on the Oxidative Stress and Polyol Pathway in the Lens of Rats with Streptozotocin-Induced Diabetes. Nutrients 2018, 10, 1423.
- Galiniak, S.; Aebisher, D.; Bartusik-Aebisher, D. Health benefits of resveratrol administration. Acta Biochim. Pol. 2019, 66, 13–21.
- Venturelli, S.; Berger, A.; Böcker, A.; Busch, C.; Weiland, T.; Noor, S.; Leischner, C.; Schleicher, S.; Mayer, M.; Weiss, T.S.; et al. Resveratrol as a pan-HDAC inhibitor alters the acetylation status of histone proteins in human-derived hepatoblastoma cells. PLoS ONE 2013, 8, e73097.
- Zheng, Y.; Liu, Z.; Yang, X.; Liu, L.; Ahn, K.S. An updated review on the potential antineoplastic actions of oleuropein. Phytother. Res. 2022, 36, 365–379.
- Bayat, S.; Derakhshan, S.M.; Derakhshan, N.M.; Khaniani, M.S.; Alivand, M.R. Downregulation of HDAC2 and HDAC3 via oleuropein as a potent prevention and therapeutic agent in MCF-7 breast cancer cells. J. Cell. Biochem. 2019, 120, 9172–9180.
- Jabczyk, M.; Nowak, J.; Hudzik, B.; Zubelewicz-Szkodzińska, B. Curcumin in Metabolic Health and Disease. Nutrients 2021, 13, 4440.
- Han, R.; Yang, H.; Li, Y.; Ling, C.; Lu, L. Valeric acid acts as a novel HDAC3 inhibitor against prostate cancer. Med. Oncol. 2022, 39, 213.
- Wu, Y.; Duan, Z.; Qu, L.; Zhang, Y.; Zhu, C.; Fan, D. Gastroprotective effects of ginsenoside Rh4 against ethanol-induced gastric mucosal injury by inhibiting the MAPK/NF-κB signaling pathway. Food Funct. 2023, 14, 5167–5181.
- Dong, F.; Qu, L.; Duan, Z.; He, Y.; Ma, X.; Fan, D. Ginsenoside Rh4 inhibits breast cancer growth through targeting histone deacetylase 2 to regulate immune microenvironment and apoptosis. Bioorganic Chem. 2023, 135, 106537.
- Liu, H.; Wang, J.; He, T.; Becker, S.; Zhang, G.; Li, D.; Ma, X. Butyrate: A Double-Edged Sword for Health? Adv. Nutr. 2018, 9, 21–29.
- Fan, P.; Li, L.; Rezaei, A.; Eslamfam, S.; Che, D.; Ma, X. Metabolites of Dietary Protein and Peptides by Intestinal Microbes and their Impacts on Gut. Curr. Protein Pept. Sci. 2015, 16, 646–654.
- Canani, R.B.; Di Costanzo, M.; Leone, L.; Pedata, M.; Meli, R.; Calignano, A. Potential beneficial effects of butyrate in intestinal and extraintestinal diseases. World J. Gastroenterol. 2011, 17, 1519–1528.
- Andrade, F.; Nagamine, M.; De Conti, A.; Chaible, L.; Fontelles, C.; Junior, A.J.; Vannucchi, H.; Dagli, M.; Bassoli, B.; Moreno, F.; et al. Efficacy of the dietary histone deacetylase inhibitor butyrate alone or in combination with vitamin A against proliferation of MCF-7 human breast cancer cells. Braz. J. Med. Biol. Res. 2012, 45, 841–850.
- Wang, S.; Li, Z.; Ma, Y.; Liu, Y.; Lin, C.-C.; Li, S.; Zhan, J.; Ho, C.-T. Immunomodulatory Effects of Green Tea Polyphenols. Molecules 2021, 26, 3755.
- Thakur, V.S.; Gupta, K.; Gupta, S. Green tea polyphenols causes cell cycle arrest and apoptosis in prostate cancer cells by suppressing class I histone deacetylases. Carcinogenesis 2012, 33, 377–384.
- Dima, C.; Assadpour, E.; Nechifor, A.; Dima, S.; Li, Y.; Jafari, S.M. Oral bioavailability of bioactive compounds; modulating factors, in vitro analysis methods, and enhancing strategies. Crit. Rev. Food Sci. Nutr. 2023, 1–39.
- Rosenfeld, R.M.; Juszczak, H.M.; Wong, M.A. Scoping review of the association of plant-based diet quality with health out-comes. Front. Nutr. 2023, 10, 1211535.
- Li, B.; Shao, H.; Gao, L.; Li, H.; Sheng, H.; Zhu, L. Nano-drug co-delivery system of natural active ingredients and chemotherapy drugs for cancer treatment: A review. Drug Deliv. 2022, 29, 2130–2161.
- Nakagawa, Y.; Mukai, S.; Yamada, S.; Matsuoka, M.; Tarumi, E.; Hashimoto, T.; Tamura, C.; Imaizumi, A.; Nishihira, J.; Nakamura, T. Short-term effects of highly-bioavailable curcumin for treating knee osteoarthritis: A randomized, double-blind, placebo-controlled prospective study. J. Orthop. Sci. 2014, 19, 933–939.
- Kanai, M.; Imaizumi, A.; Otsuka, Y.; Sasaki, H.; Hashiguchi, M.; Tsujiko, K.; Matsumoto, S.; Ishiguro, H.; Chiba, T. Dose-escalation and pharmacokinetic study of nanoparticle curcumin, a potential anticancer agent with improved bioavailability, in healthy human volunteers. Cancer Chemother. Pharmacol. 2012, 69, 65–70.
- Sasaki, H.; Sunagawa, Y.; Takahashi, K.; Imaizumi, A.; Fukuda, H.; Hashimoto, T.; Wada, H.; Katanasaka, Y.; Kakeya, H.; Fujita, M.; et al. Innovative Preparation of Curcumin for Improved Oral Bioavailability. Biol. Pharm. Bull. 2011, 34, 660–665.
- Li, Z.; Wang, L.; Lin, X.; Shen, L.; Feng, Y. Drug delivery for bioactive polysaccharides to improve their drug-like properties and curative efficacy. Drug Deliv. 2017, 24 (Suppl. 1), 70–80.
- Prasad, S.; Gupta, S.C.; Tyagi, A.K.; Aggarwal, B.B. Curcumin, a component of golden spice: From bedside to bench and back. Biotechnol. Adv. 2014, 32, 1053–1064.
- Passos, C.L.A.; Polinati, R.M.; Ferreira, C.; dos Santos, N.A.N.; Lima, D.G.V.; da Silva, J.L.; Fialho, E. Curcumin and melphalan cotreatment induces cell cycle arrest and apoptosis in MDA-MB-231 breast cancer cells. Sci. Rep. 2023, 13, 13446.
- Nootim, P.; Kapol, N.; Bunchuailua, W.; Poompruek, P.; Tungsukruthai, P. Current state of cancer patient care incorporating Thai traditional medicine in Thailand: A qualitative study. J. Integr. Med. 2020, 18, 41–45.
- Yazdi, N.; Salehi, A.; Vojoud, M.; Sharifi, M.H.; Hoseinkhani, A. Use of complementary and alternative medicine in pregnant women: A cross-sectional survey in the south of Iran. J. Integr. Med. 2019, 17, 392–395.
- Ouyang, W.; Meng, Y.; Guo, G.; Zhao, C.; Zhou, X. Efficacy and safety of traditional Chinese medicine in the treatment of osteonecrosis of the femoral head. J. Orthop. Surg. Res. 2023, 18, 600.
- Bu, Z.-J.; Liu, Y.-N.; Shahjalal, M.; Zheng, Y.-Y.; Liu, C.-J.; Ye, M.-M.; Xu, J.-Y.; Peng, X.-Y.; Wang, X.-H.; Chen, X.; et al. Comparative effectiveness and safety of Chinese medicine belly button application for childhood diarrhea: A Bayesian network meta-analysis of randomized controlled trials. Front. Pediatr. 2023, 11, 1180694.
- Kromhout, D.; Yasuda, S.; Geleijnse, J.M.; Shimokawa, H. Fish oil and omega-3 fatty acids in cardiovascular disease: Do they really work? Eur. Heart J. 2012, 33, 436–443.
- Björkström, N.K.; Strunz, B.; Ljunggren, H.-G. Natural killer cells in antiviral immunity. Nat. Rev. Immunol. 2022, 22, 112–123.
- Thies, F.; Nebe-von-Caron, G.; Powell, J.R.; Yaqoob, P.; Newsholme, E.A.; Calder, P.C. Dietary supplementation with eicosapentaenoic acid, but not with other long-chain n-3 or n-6 polyunsaturated fatty acids, decreases natural killer cell activity in healthy subjects aged >55 y. Am. J. Clin. Nutr. 2001, 73, 539–548.
- Fortmann, S.P.; Burda, B.U.; Senger, C.A.; Lin, J.S.; Whitlock, E.P. Vitamin and Mineral Supplements in the Primary Prevention of Cardiovascular Disease and Cancer: An Updated Systematic Evidence Review for the U.S. Preventive Services Task Force. Ann. Intern. Med. 2013, 159, 824–834.
- Beck, K.L.; von Hurst, P.R.; O’Brien, W.J.; Badenhorst, C.E. Micronutrients and athletic performance: A review. Food Chem. Toxicol. 2021, 158, 112618.
- Zhang, X.; Li, H.; Lv, X.; Hu, L.; Li, W.; Zi, M.; He, Y. Impact of Diets on Response to Immune Checkpoint Inhibitors (ICIs) Therapy against Tumors. Life 2022, 12, 409.
- Kim, S.-K.; Guevarra, R.B.; Kim, Y.-T.; Kwon, J.; Kim, H.; Cho, J.H.; Kim, H.B.; Lee, J.-H. Role of Probiotics in Human Gut Microbiome-Associated Diseases. J. Microbiol. Biotechnol. 2019, 29, 1335–1340.
- Bedada, T.L.; Feto, T.K.; Awoke, K.S.; Garedew, A.D.; Yifat, F.T.; Birri, D.J. Probiotics for cancer alternative prevention and treatment. BioMedicine 2020, 129, 110409.
- Suez, J.; Zmora, N.; Zilberman-Schapira, G.; Mor, U.; Dori-Bachash, M.; Bashiardes, S.; Zur, M.; Regev-Lehavi, D.; Brik, R.B.-Z.; Federici, S.; et al. Post-Antibiotic Gut Mucosal Microbiome Reconstitution Is Impaired by Probiotics and Improved by Autologous FMT. Cell 2018, 174, 1406–1423.e16.
- Spencer, C.N.; Gopalakrishnan, V.; McQuade, J.; Andrews, M.C.; Helmink, B.; Khan, M.W.; Sirmans, E.; Haydu, L.; Cogdill, A.; Burton, E.; et al. Abstract 2838: The gut microbiome (GM) and immunotherapy response are influenced by host lifestyle factors. Cancer Res 2019, 79 (Suppl. 13), 2838.
- Kuang, R.; Binion, D.G. Should high-fiber diets be recommended for patients with inflammatory bowel disease? Curr. Opin. Gas-Troenterol. 2022, 38, 168–172.
- Marques, F.Z.; Nelson, E.; Chu, P.-Y.; Horlock, D.; Fiedler, A.; Ziemann, M.; Tan, J.K.; Kuruppu, S.; Rajapakse, N.W.; El-Osta, A.; et al. High-Fiber Diet and Acetate Supplementation Change the Gut Microbiota and Prevent the Development of Hypertension and Heart Failure in Hypertensive Mice. Circulation 2017, 135, 964–977.
- Kocher, F.; Amann, A.; Zimmer, K.; Geisler, S.; Fuchs, D.; Pichler, R.; Wolf, D.; Kurz, K.; Seeber, A.; Pircher, A. High indoleamine-2,3-dioxygenase 1 (IDO) activity is linked to primary resistance to immunotherapy in non-small cell lung cancer (NSCLC). Transl. Lung Cancer Res. 2021, 10, 304–313.
- Weber, D.D.; Aminzadeh-Gohari, S.; Tulipan, J.; Catalano, L.; Feichtinger, R.G.; Kofler, B. Ketogenic diet in the treatment of cancer—Where do we stand? Mol. Metab. 2020, 33, 102–121.
- Lussier, D.M.; Woolf, E.C.; Johnson, J.L.; Brooks, K.S.; Blattman, J.N.; Scheck, A.C. Enhanced immunity in a mouse model of malignant glioma is mediated by a therapeutic ketogenic diet. BMC Cancer 2016, 16, 310.
- Orillion, A.R.; Damayanti, N.P.; Shen, L.; Adelaiye-Ogala, R.; Affronti, H.C.; Elbanna, M.; Chintala, S.; Ciesielski, M.J.; Fontana, L.; Kao, C.; et al. Dietary Protein Restriction Reprograms Tumor-Associated Macrophages and Enhances Immunotherapy. Clin. Cancer Res. 2018, 24, 6383–6395.