1. Introduction
Malaria is a mosquito-borne infection caused by an apicomplexan parasite of the genus
Plasmodium and is transmitted through the bite of a mosquito vector of the genus
Anopheles. In humans, malaria is caused by six
Plasmodium species, including
Plasmodium falciparum,
Plasmodium vivax,
Plasmodium malariae,
Plasmodium ovale curtisi,
Plasmodium ovale wallikeri, and the zoonotic parasite
Plasmodium knowlesi. Of the human malaria parasites,
P. falciparum causes the most significant morbidity and mortality. In 2021, clinical malaria affected 247,000 million individuals worldwide, leading to 619,000 deaths, with the highest burden observed in sub-Saharan Africa
[1]. Over the last two decades, artemisinin-based combination therapies (ACTs), intermittent preventive treatments (IPTs), and vector control approaches have significantly reduced the burden of malaria in some areas
[1]. However, despite tremendous efforts to control malaria, the emergence of resistant parasites to current antimalarials and the resistance of mosquitos to insecticides are significant obstacles to the global malaria eradication/elimination program
[1].
With the renewed interest in malaria elimination and eradication, it has been recognized that intervention strategies should target all stages of the parasite life cycle with a particular interest in preventing transmission
[2].
Plasmodium transmission from the human host to the mosquito vector and the parasite stages within the mosquito are promising targets for successfully interrupting the parasite life cycle and reducing the malaria burden in endemic areas
[3].
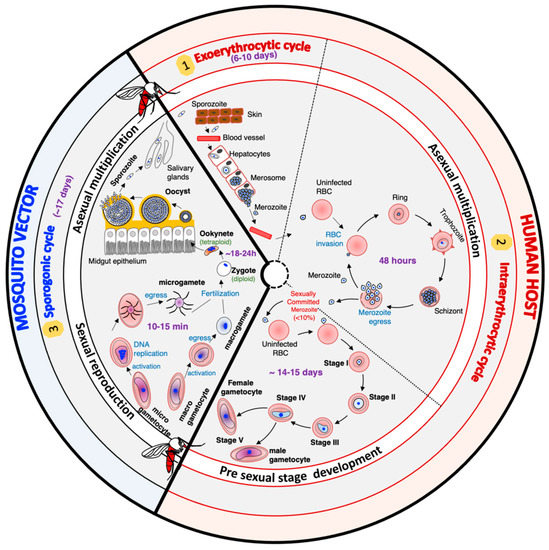
The life cycle of
Plasmodium falciparum is very complex. It includes different hosts, different cell types within the same host, asexual multiplication phases, and a sexual replication phase (
Figure 1). In humans, the cycle begins with the inoculation of the sporozoite forms of the parasite by an infected mosquito during its blood meal. These sporozoites migrate through the bloodstream, reach the liver, and enter the hepatocytes, where they multiply asexually. After six to ten days, the rupture of infected hepatocytes releases thousands of merozoites, a new form of the parasite, into the bloodstream. The merozoites invade the red blood cells (RBCs) to initiate the intraerythrocytic cycle. After two days, new merozoites are released, and these offspring enter new RBCs to perpetuate the intraerythrocytic cycle responsible for the clinical manifestations of malaria in humans. A small percentage of merozoites differentiate into sexual forms called gametocytes, which mediate transmission. Gametocytes are morphologically (shape, size, and flexibility)
[4][5][6][4,5,6] and functionally different from asexual parasites
[6][7][8][9][6,7,8,9]. Mosquitos ingest male and female gametocytes during a blood meal, which results in a wave of differentiation processes and developmental stages within the mosquito
[6][10][11][12][6,10,11,12]. In the mosquito’s gut lumen, gametocytes transform into male and female gametes. These gametes then fuse to produce a zygote that differentiates into a highly motile and invasive ookinete
[6][11][13][6,11,13]. The ookinete invades the mosquito epithelial cell wall to form an oocyst, which in turn will ultimately produce thousands of sporozoites through a process known as sporogony
[6][12][14][6,12,14]. The sporozoites will migrate and reside in the mosquito salivary gland
[12], waiting to be transmitted to the vertebrate host during a blood meal and perpetuating the parasite’s life cycle.
Figure 1.
The life cycle of
Plasmodium falciparum
parasites.
2. Gametocyte Development
The development of male and female gametocytes within the vertebrate host and the sexual recombination in the mosquito vector are critical steps in the
Plasmodium falciparum life cycle. These biological stages ensure parasite virulence and transmission. During the erythrocytic cycle, 1–5% of asexual parasites switch from the asexual multiplication pathway to a sexual differentiation pathway. The differentiation of sexually committed parasites into mature gametocytes is divided into five developmental stages (stage I–V) over 14 days. The differentiation process is characterized by dramatic alterations in parasite size, shape, and flexibility. Only early stages (stage I) and mature stage V are detectable in the peripheral blood, as the other stages are sequestered in the bone marrow and possibly other tissues
[15][93]. The differentiation process underlying asexual to stage V gametocyte formation is governed by specific expression mechanisms extensively described in other reviews
[16][17][18][94,95,96]. The following paragraphs summarize gene expression regulation mechanisms involved in asexual to early gametocyte formation.
Several environmental and metabolic factors that trigger sexual commitment, including antimalarial drugs
[19][20][97,98], lysophosphatidylcholine
[21][22][99,100], host immune response
[23][24][101,102], and parasite factors
[25][26][103,104], have been reported. ApiAP2-G was the first transcription factor shown to govern the transcription of gametocyte-associated genes
[27][28][24,25], and its regulation requires interaction with a second transcription factor,
PfAP2-I
[29][105]. The cell surface receptors involved in signal transduction and the signaling cascade leading to the activation of ApiAp2-G transcription factors still need to be characterized.
While the
P. falciparum genome is predominantly acetylated during the asexual developmental cycle, ApiAP2-G appears to be repressed, and its locus was shown to be associated with histone heterochromatin marks, including H3K4ac, H3K9me1, H3K36me2,
PfHP1, and the gametocyte essential factor
PfAP2-G5
[30][106].
PfHP1-dependent gene silencing is antagonized by a sexual commitment activator protein called
PfGDV1 (
Plasmodium falciparum gametocyte development 1)
[31][32][107,108].
PfGDV1 targets heterochromatin and triggers the eviction of
PfHP1. Interestingly,
PfGDV1 activation is, in turn, controlled by a multi-exon long non-coding
gdv1 antisense RNA (
Pfgdv1asRNA) that initiates downstream of the
gdv1 locus
[31][107].
The gametocyte essential factor AP2-G5 further prevents sexual commitment
[32][108]. Indeed,
PfAP2-G5 binds upstream of the
Pfap2-g locus and other exogenic regions, hence suppressing ap2-g expression
[32][108]. Although findings strongly suggest that ApiAP2 genes are under epigenetic control, histone post-translational modification linked to parasite conversion remains to be characterized. In addition, the signaling cascade events leading to the removal of the repressive state by
PfGDV1 and the activation of
PfApiAP2-G remain unknown.
Sexual conversion can occur by two possible routes following specific histone modification marks that promote changes in gene expression and sexual differentiation
[29][33][105,109]. One route suggests that the decision to undergo gametocyte development occurs during the asexual cycle preceding the erythrocyte reinvasion event that leads to gametocyte formation. In this model, the decision to undergo gametocyte development is linked to
PfAP2-G expression in the schizont of the preceding cycle
[33][109]. The second route implies that the decision occurs within the same cycle, during the initial
PfAP2-G expression in ring stages
[33][109]. Recently, Li et al. identified an ApiAP2 transcription factor in
P. yoelii, AP2-O3, implicated in a gender-specific transcription program
[34][110]. AP2-O3, expressed predominantly in female gametocytes, represses the expression of male-specific genes. Genetic studies have shown that the depletion of either HP1 or the histone deacetylase enzyme 2 (Hda2) resulted in the activation of
ap2-g transcription and many heterochromatic genes
[34][110].
Gametocyte development from stage I to stage V is characterized by euchromatic post-translational modifications and an abundance of repressive methylation marks on histone 3. Ealy gametocyte stages (I to III) are characterized by the H3K9me3, H3K27me2, H3K27me3, H3K36me2, H3K37me1, H3R17me1, and H3R17me2 modifications
[30][35][46,106]. The role of arginine methylation as a key feature for the epigenetic regulation of gametocyte development and maturation was suggested by Von Grüning et al.
[30][106].
Three-dimensional structure analyses of the
P. falciparum genome revealed that the localization and interaction of sexual differentiation genes in a repressive center are critical for regulating sexual conversion
[36][44]. Further studies are needed to unravel the machinery regulating genome localization and the signaling pathway governing sexual commitment. The repertoire of environmental triggers for sexual conversion is far from being exhaustive. Characterizing proteins interacting with the histone-associated protein
PfHP1 and
PfGDV1 will undoubtedly unravel the molecular player involved in parasite conversion.
Despite their similitude with human enzymes, histone-modifying enzymes could be potential targets for developing antimalarial drugs with transmission-blocking activities. Stenzel et al.
[37][111] designed, synthesized, and tested the biological activity of thirteen terephthalic-acid-based HDAC inhibitors. A subset of these compounds had moderate activity against
P. falciparum gametocytes but showed sub-micromolar transmission-blocking activity against the rodent malaria parasite
P. berghei. Hence, histone-modifying enzymes are potential targets to prevent malaria infection in the mosquito.
3. Gamete Development
Establishing a successful malaria infection in the
Anopheles mosquito and the subsequent spread of competent parasites depend on the coordinated development of the
Plasmodium parasite in the mosquito midgut. The differentiation process in the mosquito midgut, lasting almost 20 h, is defined by the rapid conversion of mature stage V female and male gametocytes into female macrogametes and male microgametes, respectively
[6][10][11][6,10,11]. This step is followed by the sexual reproduction between macro- and microgametes and, finally, the conversion of newly formed zygotes into ookinetes
[6][11][6,11]. The transition in the mosquito midgut constitutes a significant bottleneck in the
Plasmodium life cycle as there is a 316-fold loss in parasite abundance from the gametocyte to ookinete stage and a 100-fold loss from the ookinete to oocyst stage
[38][112]. Due to this considerable reduction in the parasite population, the plasmodial midgut stages are attractive targets for developing transmission-blocking interventions. Hence, malaria parasite developmental stages in the mosquito midgut have been extensively studied, and several key mechanisms and proteins orchestrating the differentiation process have been described, providing potential targets for vaccines and drug development
[39][113].
Gamete formation is morphologically characterized by a rounding-up of the cell, followed by the parasitophorous vacuole membrane (PVM) rupture and parasite egress from the erythrocyte
[10][40][10,114]. The differentiation of male gametocytes into male gametes involves three successive mitotic DNA replications that will produce eight motile microgametes through a process called exflagellation. Upon completion of DNA replication, the microgametes egress from the host cell
[10]. In contrast, the differentiation of female gametocytes results in a rounding-up of the parasite with no DNA replication and the emergence of the forming gamete from the infected red blood cell
[6].
3.1. Gene Expression Regulation Controlling Gametogenesis
Transcriptomic analysis of
P. falciparum gametocytes and gametes revealed that small transcriptome changes characterize gametogenesis compared to other life-stage differentiation processes
[41][115]. Most studies investigating gamete to sporozoite formation rely on
Plasmodium berghei, causing rodent malaria. The rodent parasites are not of direct practical concern to humans. However, the easy access to
Plasmodium life stages in the mosquito and the practicability for the experimental study of human malaria made them a good model for human malaria.
In
Plasmodium berghei, the change in gene expression necessary for female gametogenesis is determined by the translation of a large number of mRNAs maintained in a repressive translational state
[42][43][116,117]. Indeed, in female gametocytes, many transcripts are synthesized but translationally repressed until needed for macrogamete formation and zygote-to-ookinete transformation. Using the
Plasmodium berghei model system, Guerreiro et al. revealed that approximately 50% of the transcriptome is maintained in this translationally repressed state by a messenger ribonucleoprotein (mRNP) composed of 16 major factors, including the RNA helicase DOZI (development of zygote inhibited) and the Sm-like factor CITH (homolog of worm CAR-I and fly Trailer Hitch)
[44][118]. Using an in vitro translation assay, Tarique et al. characterized
P. falciparum DOZI (
PfDZ50) and demonstrated that the protein inhibits translation
[45][119], suggesting a function similar to that reported for
P. berghei female gametogenesis.
The expression of stage-specific genes that are necessary for microgamete formation is regulated by transient and reversible protein phosphorylation followed by the de novo synthesis of genes involved in DNA replication, axoneme assembly and motility, chromatin condensation, cytokinesis, and exflagellation
[46][47][120,121]. Proteomic studies in
P. berghei gametes revealed that proteins implicated in RNA translation, protein biosynthesis, glycolysis, environmental stress response, and tubulin-associated cytoskeleton dynamics are predominantly regulated during gamete formation
[48][122]. Gene expression and protein synthesis regulation during both male and female gametogenesis depend highly on the signaling cascade orchestrated by stage-specific kinases and phosphatases
[46][49][50][120,123,124].
3.2. Signaling Cascade Controlling Gametogenesis
Gametocyte activation is stimulated by environmental stimuli, including a temperature drop by approximately 5 °C, the presence of mosquito-derived xanthurenic acid (XA), and an increase in extracellular pH from 7.2 to about 8
[51][52][53][125,126,127].
Plasmodium receptors responsible for sensing temperature drops and binding to XA have not been identified yet. The exposure of
P. falciparum gametocytes to XA has been shown to increase cyclic GMP (cGMP) levels in the gametocyte, suggesting parasite guanylyl cyclase (GC) activation
[54][128]. In the rodent malaria parasites, the increase in cGMP eventually leads to an increase in the cytoplasmic calcium level necessary to trigger a calcium-dependent stage-specific gene expression and regulation pathway
[55][56][57][129,130,131].
The
Plasmodium falciparum genome encodes two guanylyl cyclase proteins (GCα and GCβ) that are only expressed in sexual-stage parasites. Functional studies in
P. falciparum revealed that these proteins are active guanylyl cyclases and likely important in gametocyte activation
[58][132]. Pharmacological and genetic studies investigating the signaling cascade of
P. falciparum and
P. berghei gamete formation have shown that GC activation leads to the formation of a secondary messenger cyclic GMP (cGMP), which in turn activates the protein kinase G (PKG), the unique effector identified in
Plasmodium [54][59][128,133]. Taylor et al. revealed that the tight regulation of cGMP concentration is critical for
P. falciparum gametocyte conversion and that premature high cGMP levels are deleterious for gamete formation
[60][134]. Bennink et al. suggested that activation of PKG leads to the hydrolysis of phosphatidylinositol-(4,5)-bisphosphate (PIP2) by the phosphoinositide-specific phospholipase C (PI-PLC) and the production of two secondary messengers, diacylglycerol (DAG) and inositol triphosphate (IP3)
[61][62][135,136]. Although IP3 induces the opening of calcium channels on the
Plasmodium endoplasmic reticulum membrane, no ortholog of an IP3 receptor channel has yet been identified. A recent biochemical approach of an IP3 affinity chromatography column combined with bioinformatics has revealed a potential transporter associated with multidrug resistance in
P. falciparum [63][137]. Additional work will elucidate the function of this novel protein.
The rapid release of calcium in mature gametocytes mediates a stage-specific calcium-dependent effector pathway in all gametogenesis steps, including gamete formation, gamete egress, microgamete mitotic maturation, and exflagellation
[54][59][61][128,133,135]. In
Plasmodium, the intracellular calcium level is sensed by calcium-dependent protein kinases (CDPKs)
[64][138]. Macro- and microgamete-specific CDPKs that regulate the expression and activation of genes necessary for converting gametocytes into gametes have been identified
[47][65][66][121,139,140].
In mature female gametocytes, the increase in cytoplasmic Ca
2+ is sensed by CDPK1
[65][139]. CDPK1 has been identified as the key protein that regulates the translation of mRNAs in a temporal and stage-specific manner during macrogamete formation
[65][139].
The differentiation of male gametocytes is regulated by a male-specific calcium-dependent protein kinase CDPK4, which initiates DNA replication, axoneme assembly, and cell motility
[47][67][121,141]. In
P. berghei, the lysis of the host cell membrane(s) surrounding the microgametocyte is mediated by CDPK1
[65][139]. In addition, CDPK1 translationally activates mRNA species in the developing zygote that remain repressed in macrogametes
[65][139]. The cell-division cycle protein 20 (CDC20) appears to regulate male gametocyte mitotic division but not the one during schizogony
[68][142]. Whether CDC20 works with CDPK1 and anaphase-promoting complex 3 (APC3) to modulate chromosome condensation and cytokinesis for microgamete formation is not yet determined
[69][143]. In
P. berghei, a histone chaperone protein termed FACT-L (facilitates chromatin transcription), which facilitates chromatin transcription, was shown to be involved in male gametocyte DNA replication and the production of fertile microgametes
[70][144]. The formation of the flagella requires the formation of basal bodies and the assembly of axonemes.
Upon completion of DNA replication, axonemes become motile, facilitating the egress of the microgametes from the host cell. Genetic studies in
P. berghei have revealed that axoneme assembly and motility require several proteins, including the stage-specific Actin II
[71][145], the armadillo-repeat motif protein
Pf16
[72][146], the spindle-assembly-related protein SAS6
[73][147], the SR protein kinase (SRPK)
[74][148], and a gametocyte-specific mitogen-activated protein kinase 2 (MAP2)
[75][76][149,150]. The
Plasmodium falciparum kinome includes four NIMA-related kinases (
PfNEK 1 to 4). While
PfNEK-1 is expressed in asexual and sexual stages, the mRNA transcripts of
PfNEK-2,
PfNEk-3, and
PfNEK-4 are exclusively expressed in gametocytes. NEK1 and NEK3 have been shown to activate the atypical MAP2 protein through phosphorylation
[77][78][151,152].
PfNEK-2 and
PfNEK-4 are required for meiosis completion in the ookinete
[79][80][153,154]. A metallo-dependent protein phosphatase, PPM1, also plays an important role in
P. berghei male gametocyte exflagellation
[50][124].
Kinases and phosphatases involved in gamete formation can be considered promising targets for drug development. A high-throughput screening of
P. falciparum-cGMP-dependent protein kinase identified a thiazole scaffold that kills erythrocytic and sexual-stage parasites
[81][155]. Since the malarian PKG differs from the mammalian PKGs, this scaffold represents a good starting point for developing a new class of antimalarial drugs. Recently, Xitong et al.
[82][156] assessed the activity of 25 phosphatase inhibitors against
Plasmodium berghei sexual development and transmissibility to the mosquito. Two compounds from the panel effectively inhibited different development stages, from gametogenesis to ookinete maturation. These examples highlight that in silico modeling and screening combined with in vivo and ex vivo approaches using mouse models and human malaria could help identify parasite kinase and phosphatase inhibitors. The kinases regulating male gamete exflagellation (CDPK4 and atypical MAP-2) and DNA replication (Nek-2, Nek-4) should be of great interest in the screening.
Although rodent malaria parasites are sound model systems to understand P. falciprum gametogenesis, more studies relying on human malaria parasites are necessary to grasp the complexity of the early stages of malaria transmission. Those studies should consider including field studies to better understand the biology of transmission during natural infection.
4. Zygote to Ookinete Development
4.1. Gamete Fusion and Zygote Formation
The zygote results from the fusion between a fertile male microgamete and a female macrogamete. Fertilization of a macrogamete by a microgamete is mediated by stage- and sex-specific proteins synthesized in the respective gamete before the fusion event. The proteins involved in gamete fusion also mediate the prerequisite recognition and attachment steps. Genetic studies of
P. falciparum and
P. berghei genes have revealed that P48/45 and P230, two members of a protein family defined by a disulfide bonding pattern of six conserved cysteine residues, are essential for male gamete fertility and fusion with macrogametes
[83][84][85][157,158,159]. In
P. falciparum, P48/45 and P230 are localized on the gamete surface and have been shown to form a complex necessary for the fusion of the microgamete to the macrogamete. In vitro and in vivo studies in the mouse model have demonstrated the critical implication of P48/45 and P230 in fertilization. These findings led to their use as targets for transmission-blocking vaccines
[39][86][113,160]. In macrogamete
Pfs47, a paralog of P48/45 was identified
[87][161]. Functional studies demonstrated the expression of
Pfs47 exclusively on the surface of female gametes, but the protein did not appear crucial for fertilization
[87][161].
The evolutionarily conserved class II gamete fusogen HAP2/GCS1 (hapless 2/Generative-cell-specific 1) has also been implicated in the fusion of the plasma membranes of two haploid gametes
[88][162]. Gene disruption studies of
p48/45 or
hap2/gcs1 resulted in sterility due to the inability of the male gamete to either attach or fuse to fertile female gametes
[85][88][159,162]. Studies investigating gamete fusion in
Plasmodium revealed that a histone chaperone protein named FACT (facilitates chromatin transcription) plays an essential role in fertilization. However, the mechanism of action of this nuclear protein is still unclear
[70][144].
The fusion of the two gametes requires the prior disassembly of a peculiar organelle located underneath the parasite plasma membrane and is named the inner membrane complex (IMC)
[5]. The IMC will be reassembled later to ensure the pellicle’s integrity and parasite polarity
[5][89][5,163]. The IMC is a membranous scaffold in the Alveolata, a group of diverse unicellular eukaryotes, including
Plasmodium spp.,
Toxoplasma gondii, the ciliates, and dinoflagellates. This membranous patchwork is anchored to the subpellicular microtubule network. It also interacts with various parasite-specific proteins to coordinate parasite morphological changes, the segmentation of daughter cells during asexual replication, and parasite motility. Interestingly, the localization and interaction of many IMC proteins are regulated by post-translational S-palmitoylation mediated by the palmitoyl-acyl-transferase DHHC2 (the ortholog of
PfDHHC1)
[90][164]. Inhibitor studies revealed that DHHC2 palmitoylation is critical to zygote differentiation during the initial mosquito infection with
P. berghei [91][165]. Hence, stage-specific palmitoylation enzymes could be novel targets for disrupting IMC assembly and zygote formation and differentiation into ookinetes.
The concept of transmission-blocking immunity is mostly antibody-mediated. Therefore, the development of transmission-blocking vaccines (TBVs) focuses on inducing potent antibodies sustained at adequate levels. Extensive efforts toward the clinical development of
P. falciparum TBVs are undertaken, but the functional activities associated with most antibodies remain modest. Further study must be conducted to determine the players involved in gamete fusion and their association with each other to orchestrate gamete fusion. The disassembly of the IMC is a pre-requisite for gamete fusion, and targeting the enzymes modulating this biological process could be an approach to interfere with the establishment of the infection in the mosquito gut.
Plasmodium falciparum is predicted to encode 12 putative palmitoyl acyl-transferases thought to ensure lipid-based palmitoylation of parasite proteins and act as a biological rheostat for protein–protein interactions and subcellular trafficking
[92][166]. Inhibitors of palmitoylation enzymes could constitute a new class of antimalarial drugs targeting multiple parasite life stages and other apicomplexan parasites of medical importance
[92][93][94][166,167,168].
4.2. Molecular and Genetic Mechanisms of Fertilization
Gene expression studies in
P. berghei revealed that zygote development and differentiation into ookinetes require the transcriptional activation of several maternal silenced mRNAs in the zygote. A study investigating the parental contribution of transcripts implicated in zygote differentiation revealed that while inherited maternal mRNAs are activated to drive the early stage of zygote differentiation, the paternal alleles are initially silenced and then reactivated
[95][96][19,169].
The derepression of a maternal transcript post fertilization will drive significant morphological changes defined by the formation of the IMC and the secretory organelles, as well as the elongation and apical polarization of the differentiating zygote
[5][97][98][5,170,171]. The dynamic organization of the IMC and the parasite cytoskeleton is essentially coordinated post-translationally by reversible phosphorylation
[65][74][99][139,148,172] and palmitoylation
[91][100][165,173]. A systematic analysis of the
Plasmodium kinome, combined with genetic studies in
P. berghei, revealed that protein kinase 7 (PK7) and cyclin-G-associated protein (GAK) are essential in ookinete formation
[74][99][148,172]. GAK is predicted to regulate clathrin-mediated vesicle trafficking and membrane fusion, suggesting its involvement in the formation of the secretory organelles or the assembly of the IMC
[74][148]. PK7 mutants were blocked in ookinete formation, but the mechanism by which this kinase regulates ookinete development remains undetermined. Reverse genetics studies revealed that the
Plasmodium phosphatase PPKL (protein phosphatase with kelch-like domains) is essential during ookinete differentiation and is involved in defining ookinete polarity, pellicle morphology and integrity, and ookinete motility
[101][174].
Plasmodium falciparum DHHC1, a palmitoyl-S-acyl-transferase (PAT) containing a conserved DH(H/Y)C motif, was shown to be exclusively localized to the IMC
[91][100][165,173]. DHHC1 is apicomplexan-specific and was implicated in ookinete formation and morphogenesis
[91][100][165,173].
Subsequent regulation of stage-specific gene expression requires a de novo synthesis of ookinete-specific genes by the transcription factor ApiAp2-O
[102][175]. ApiAp2-O appears to be associated with more than 500 genes involved in ookinete development, motility, midgut invasion, and parasite escape from mosquito immunity
[103][78]. The ookinete-specific genes mediate nuclear fusion in the diploid zygote, DNA replication, and meiosis that will produce a motile tetraploid ookinete
[104][176]. The tetraploid state was shown to persist throughout the ookinete stage until the formation of sporozoites in the oocyst
[104][105][176,177]. Like in eukaryotic model systems, meiosis and cell cycle progression in the
Plasmodium parasite are regulated by NIMA-related kinases (Neks). Gene disruption studies in
P. berghei revealed that Nek-4 and Nek-2 are abundantly expressed during the gametocyte stages. Nek-4 and Nek-2 are essential for zygote differentiation into the ookinete stage but not for gamete formation and fertilization
[79][80][153,154]. Given the success in developing drugs targeting human kinases,
Plasmodium kinases are attractive targets for the next generation of antimalarials. Indeed, ongoing efforts attempt to characterize
Plasmodium kinases while evaluating them as antimalarial drug targets
[106][107][178,179].
4.3. Formation and Maturation of Ookinete
Ookinete maturation is completed 19 to 36 h post-gametocyte ingestion in the blood meal. The ookinete will quickly exit the midgut lumen via an intracellular or intercellular route
[108][180]. Ookinete motility is regulated by kinases. The cGMP-dependent protein kinase (PKG) pathway activates the gliding motility apparatus on the IMC, and the calcium-dependent protein kinase 3 (CDPK3) mobilizes the intracellularly stored calcium necessary for signaling
[109][110][181,182]. The traversal of the midgut wall is mediated by an impressive set of microneme proteins discharged by the mature ookinete
[111][183]. Proteomic
[111][183] and genetics studies identified key proteins for midgut traversal, including the circumsporozoite-TRAP-related protein CTRP
[112][184], the membrane-attack ookinete protein (MAOP)
[113][185], the secreted ookinete adhesive protein SOAP
[114][186], the von-Willebrand-Factor-A-domain-related protein WARP
[115][187], the cell-traversal protein for ookinetes and sporozoites CelTOS
[116][188], and the chitinase 1 (CHT1)
[117][118][189,190].
The ookinete stage is an attractive target for transmission-blocking strategies as the mosquito immune system naturally kills many ookinetes. Ookinete proteins involved in the attachment and invasion of the midgut epithelial cells are potential vaccine targets. Two ookinetes surface proteins, P25 and P28, that play a role in ookinete adhesion to the midgut and differentiation into oocyst are candidates for transmission-blocking vaccines
[39][113]. Transmission-blocking strategies comprise gametocytocidal drugs, transmission-blocking vaccines (TBV), and the engineering of genetically modified mosquitos refractory to
Plasmodium infection
[119][191]. The rationale for transmission-blocking drugs is to promote gametocyte clearance in the human host and to block onward parasite transmission by targeting blood stage parasites
[120][192]. On the contrary, transmission-blocking vaccines aim to induce, in the human host, the production of antibodies against specific proteins accessible in the mosquito gut to prevent mosquitoes from carrying and spreading the parasites. Targeted proteins include surface proteins of
Plasmodium stages found in the mosquito
[39][113] or mosquito-specific effectors implicated in the infection
[121][193]. Antibodies will be ingested by the mosquitoes during their blood meal and are expected to interact with the target in the mosquito gut before the ookinete transversal of the midgut wall. Therefore, essential ookinete surface proteins and potentially micronemal proteins secreted by the parasite at the time of gut lumen exit could be targeted by TBV approaches.
As mentioned above, parasite-specific kinases, whose functions are required for parasite motility, are also good targets for Plasmodium multi-stage drugs.
5. Ookinete to Oocyst Development
Following the traversal of the midgut epithelium, ookinetes settle in the basal lamina surrounding the gut and differentiate into oocysts. Ookinetes can be found in the basal lamina 18–24 h after an infective blood meal
[12]. The mechanisms triggering the differentiation process are still unclear, but the extracellular matrices composed of collagen, fibronectin, laminin, and chondroitin sulfate are thought to play a role
[122][194]. With the current proteomic and genomic tools, future studies should further investigate mosquito factors involved in ookinete differentiation.
Compared with other developmental stages, relatively few transcriptomics and proteomics studies assess changes associated with the early stage of oocyst development. Sequencing strand-specific cDNA libraries of seven
Plasmodium stages has revealed an abundance of antisense transcripts in gametocytes and ookinetes
[123][195]. These findings suggest that antisense RNA plays a role in gene expression regulation in sexual stages. However, the roles of these antisense transcripts still have to be investigated
[123][195]. Studies investigating the ookinete to oocyst differentiation are required to understand these processes better.