The intestinal barrier is a precisely regulated semi-permeable physiological structure that absorbs nutrients and protects the internal environment from infiltration of pathological molecules and microorganisms. Bile acids are small molecules synthesized from cholesterol in the liver, secreted into the duodenum, and transformed to secondary or tertiary bile acids by the gut microbiota. Bile acids interact with bile acid receptors (BARs) or gut microbiota, which plays a key role in maintaining the homeostasis of the intestinal barrier.
- bile acid
- bile acid receptor
- gut microbiota
- intestinal barrier
1. Introduction
2. Bile Acids and Intestinal Barrier
The integrity of the intestinal barrier requires constant renewal of the epithelial boundary, maintenance of tight junctions (physical barrier), mucus secretion and normal gut microbiota (biochemical barrier), and a finely regulated intestinal lamina propria immune system (immune barrier). Because of the complexity and heterogeneity of the intestinal barrier, specific mechanisms underlying the dysfunction of the intestinal barrier are still far from clear. That bile acids may play a pivotal role in many aspects of the maintenance of intestinal barrier integrity is a novel concept (Figure 1).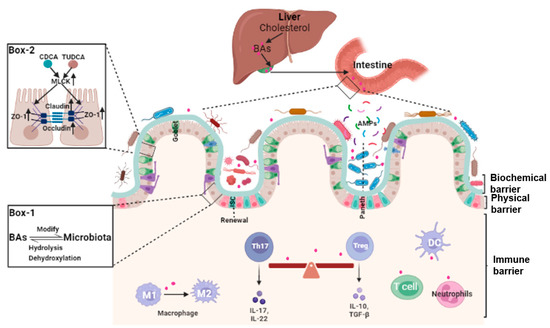
2.1. Bile Acids and Intestinal Epithelial Cells Tight Junctions
2.2. Bile Acids and Gut Microbiota
2.3. Bile Acids, Intestinal Stem Cells (ISCs), and Epithelial Injury Repair
Balance between intestinal epithelial proliferation and cell death from damage, stress, and other pathological conditions maintain normal intestinal barrier function. Intestinal epithelial cells are self-renewed every 3–5 days, in part, from Lgr5+ intestinal stem cell (ISCs) as at counteract intestinal barrier damage and different stress stimuli [17][30]. Lgr5+ ISCs replenish damaged epithelial cells and generate progenitors of goblet and Paneth cells. These cells secrete mucus and antimicrobial peptides to support the integrity of the intestinal mucus layer [18][31]. Data demonstrated that bile acids metabolism plays a potential role in the self-renewal function of Lgr5+ ISCs. Based on pathway synthesis and chemical structures, bile acids are grouped into 12α-hydroxylated (OH) bile acids and non-12α-OH bile acids [3]. What the two groups exert varies, and, at times, they have opposing effects on ISCs. For instance, the 12α-OH bile acid CA can inhibit the activity of peroxisome proliferator-activated receptor alpha (PPARα), impeding fatty acid oxidation (FAO), and the self-renewal of Lgr5+ ISCs [19][32]. An HFD-driven increase in DCA decreased ISC proliferation and differentiate into goblet cells through pathologic endoplasmic reticulum stress [20][33].2.4. Bile Acids and Intestinal Local Immune Homeostasis
The intestinal lamina propria is colonized by a variety of innate and adaptive immune cells and gut-associated lymphoid tissue and is termed the intestinal immune barrier [21][22][23][37,38,39]. Under normal conditions, this microecosystem is tightly and finely regulated. Environmental factors and the gut microbiota and their metabolites (microorganism-associated molecular patterns and pathogen-associated molecular patterns) are recognized by specific receptors (Toll-like receptors, TLRs) on immune cells, leading to intestinal immune homeostasis and self-tolerance [23][39]. Bile acids modulate immune responses in the intestine through BARs, including TGR5, FXR, VDR, CAR, and retinoic-acid-receptor-related orphan nuclear receptor-γt (RORγt). Unique lymphocyte populations function cooperatively to maintain the intestinal immune system, especially the balance between pro-inflammatory T helper 17 (Th17) cells and anti-inflammatory Treg cells [24][40]. Two derivatives of LCA, 3-oxoLCA and isoalloLCA, inhibited RORγt to suppress Th17 cell differentiation and increased the differentiation of Treg cells through the production of mitochondrial reactive oxygen species [25][41]. The secondary bile acid, isoLCA, via VDR, modulated colonic FOXP3 + Treg cells expressing RORγt and ameliorated their susceptibility to inflammatory colitis [26][42].3. Bile Acid and Intestinal-Barrier-Dysfunction-Related Diseases
3.1. Inflammatory Bowel Diseases (IBDs)
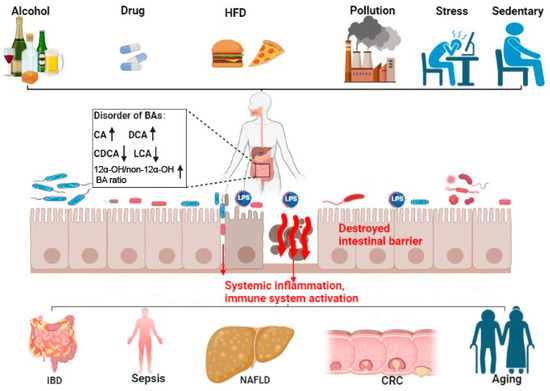
Some bile acids altered the expression of tight junction proteins and the renewal of intestinal stem cells, leading to the intestinal barrier injury, increasing the incidence of IBD [19][27][32,50]. In contrast, some other bile acids may maintain intestinal immune barrier homeostasis by activating BARs such as FXR and TGR5. The DCA-mediated activation of FXR inhibited Paneth cell function and type I interferon signaling in mice with Crohn’s disease [28][51]. Nuclear xenobiotic receptor CAR signaling altered the transcriptome of Teff cells that infiltrated the small intestine lamina propria (siLP) and suppressed Crohn’s disease-like small bowel inflammation [29][80].3.2. Gut Origin of Sepsis
The role of bile acids in intestinal barrier retention is well confirmed. However, there is also a close relationship between the metabolism of bile acids and gut-originating sepsis. Consistent with this view, the inflammatory mediators released during sepsis inhibited hepatobiliary transporter gene expression, resulting in hyperbilirubinemia and cholestasis [30][90]. Serum bile acid concentrations were significantly higher in animals and humans with sepsis. Thus, bile acids may be a potential marker for early sepsis [31][32][52,53]. Of interest is the analysis of plasma from septic individuals found to be glycochenodeoxycholate- and phenylalanine-associated with survival of sepsis [33][91].3.3. Non-Alcoholic Fatty Liver Disease (NAFLD)
Bile acids may also affect the progression of NAFLD through adjusting the intestinal barrier [34][35][36][105,106,107]. Long-term HFD intake can lead to gut dysbiosis and the aberrant metabolism of bile acids. Bidirectional crosstalk between gut microbiota and bile acids could impair the intestinal epithelial function, increasing the translocation of gut-derived endotoxins such as LPS to the blood and lymphatics to activate hepatic TLR-4/NF-κB signaling and promote NAFLD or NASH [37][38][108,109]. HFD-mediated changes in bile acids decreased FXR and TGR5 signaling and degraded the intestinal barrier [39][40][110,111].3.4. Colorectal Cancer (CRC)
The dysregulation of bile acids-BAR pathways also plays a pivotal role in the progression of CRC. Decreased FXR-FGF15 signaling and overexpressed TGR5 were observed in azoxymethane (AOM)/dextran sodium sulfate (DSS)-induced CAC mice [41][68]. In contrast, the activation of FXR protected the intestinal barrier, decreased inflammation, and restricted tumor growth [42][126]. HFD and dysregulated Wnt signaling (APC mutation) altered bile acids profiles, increased the malignant transformation of Lgr5+ cancer stem cells, and promoted adenocarcinoma progression which was counteracted by the activation of FXR [43][127].3.5. Aging
Even though longevity is the people’s long-standing pursuit, to clarify the exact mechanism and pick out the ‘Mr. Key’ in this process is still the most important to success. Fortunately, many lines of evidence have clearly demonstrated that modifying the bile acids profile can delay aging effectively. A Mediterranean diet or calorie restriction altered the gut microbiota and bile acids, which was posited to improve health during aging [44][45][46][47][135,136,137,138]. In rodents, methionine restriction increased macroautophagy/autophagy and altered bile acid conjugation and levels to lengthen lifespan [48][49][139,140]. Modifying the bile acid profile with medications or fecal transplantation also relieved age-associated metabolic dysregulation in mice [50][51][141,142].