Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Akebe Luther King Abia and Version 2 by Lindsay Dong.
The marine environment possesses diverse and complex characteristics, representing a significant challenge for microbial survival. Therefore, bacteria must develop adaptive mechanisms to thrive in such environments. Quorum sensing (QS), a well-established phenomenon in microorganisms, involves the communication between cells through chemical signals, which depends on cell density. Extensive research has been conducted on this microbial ability, encompassing the early stages of understanding QS to the latest advancements in the identification and characterization of its mechanisms.
- quorum sensing
- microbial communication
- biofilm formation
- biogeochemical cycling
1. Introduction
Extensive research has significantly advanced the understanding of bacteria, particularly concerning their lifecycle. From the earliest investigations into basic phenomena, such as food fermentation, research has progressed to uncovering the presence of disease-causing pathogens in diverse ecological niches, including plant rhizosphere soil, deep ocean zones, alpine peaks, and other extreme environments [1]. Among the notable discoveries is the phenomenon of cell-to-cell communication, known as “quorum sensing,” which occurs within bacterial communities, both within and between species. Traditionally, quorum sensing (QS) was understood to predominantly regulate bacterial bioluminescence. The concept of quorum sensing was first observed over 25 years ago in two marine bacterial species: Vibrio fischeri and Vibrio harveyi. In these species, quorum sensing is controlled by various genes and regulons, including Lux genes (LuxCDABEG) and their respective autoinducers. In Vibrio fischeri, N-acyl homoserine lactone (AHL) acts as the autoinducer, while in Vibrio harveyi, autoinducer-2 (AI-2) plays QS role [2]. These autoinducers are synthesized when the bacterial population is low, but once the bacterial density reaches a threshold, these molecules bind to their respective receptor proteins, initiating a signaling cascade. This signaling alters the expression of lux genes, thus triggering the activation of bioluminescence, a phenomenon termed ‘quorum sensing’ [3][4][5][3,4,5]. Since this initial observation, researchers have identified homologs of these genes in various bacterial communities, which play a regulatory role in quorum sensing. This communication process has been found to influence a wide range of biological processes, including bioluminescence [6], biofilm formation [7], cell competency [8], horizontal gene transfer [9], virulence factor expression [10], symbiotic relationships [11], sporulation in fungi [12][13][12,13], pigment production [14], motility [15], toxin production [16], and even antibiotic production [17][18][19][17,18,19]. These observations highlight the intricate communication networks and coordinated behavioral patterns within bacterial communities, underscoring the complexity of microbial ecosystems. Investigations into QS have significantly enriched our understanding of the regulatory mechanisms governing bacterial interactions, thereby offering novel insights into microbial ecology and behavior. Furthermore, the study of QS has sparked interest in the potential for strategic manipulation of bacterial behavior. This field of study opens up promising avenues for application across a broad range of sectors, including healthcare for infection control, environmental management for bioremediation efforts, and various advancements in biotechnology.
To effectively engage in QS, bacteria must possess certain capabilities, including producing and secreting autoinducer signaling molecules, perceiving changes in their concentration, and respond by regulating gene transcription [20][21][20,21]. The process heavily relies on the diffusion mechanism of these signaling molecules, which are typically secreted at low concentrations by individual bacteria. These molecules may simply diffuse away from the cell in environments with low cell density. However, as cell density increases, the local concentration of signaling molecules can surpass a threshold level, triggering a shift in gene expression [22]. Numerous quorum sensing systems have been extensively studied and documented in various taxa and habitats (a PubMed keyword search for “quorum sensing” generates over 12,000 results) [23]. However, many taxa and habitats remain, including deep-sea microbes, where quorum sensing remains unidentified or poorly characterized. Further research is needed to explore and understand the prevalence and specific mechanisms of quorum sensing in these diverse environments
 To date, no AI-2 receptors have been identified in Epsilonproteobacteria. However, two main receptor categories, LuxP in Vibrio spp. And LrsB in enteric bacteria are known. In deep-sea vents, Epsilonproteobacteria act as early-stage colonizers, preparing the stage for colonizing other microbes through QS signaling involving adhesive polymeric secretion. This secretion also serves as a nutrient source, promoting the recruitment of additional microbial species. The extracellular matrix of P. aeruginosa is composed of extracellular DNA (eDNA), which aids biofilm assemblage in a reaction with extracellular calcium by promoting bacterial association when released via QS-dependent and independent pathways. eDNA affects the RNA composition of members of the biofilm, acting as a nutrient source through the contents of the membrane vesicle in which it is enclosed, as well as modulating the expression of genes influencing acidity and antimicrobial resistance [32][34].
To date, no AI-2 receptors have been identified in Epsilonproteobacteria. However, two main receptor categories, LuxP in Vibrio spp. And LrsB in enteric bacteria are known. In deep-sea vents, Epsilonproteobacteria act as early-stage colonizers, preparing the stage for colonizing other microbes through QS signaling involving adhesive polymeric secretion. This secretion also serves as a nutrient source, promoting the recruitment of additional microbial species. The extracellular matrix of P. aeruginosa is composed of extracellular DNA (eDNA), which aids biofilm assemblage in a reaction with extracellular calcium by promoting bacterial association when released via QS-dependent and independent pathways. eDNA affects the RNA composition of members of the biofilm, acting as a nutrient source through the contents of the membrane vesicle in which it is enclosed, as well as modulating the expression of genes influencing acidity and antimicrobial resistance [32][34].
2. Bacterial Quorum Sensing in Oceanic Settings
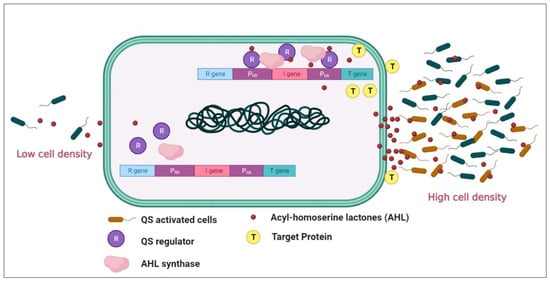
Quorum sensing (QS) is crucial in bacterial intercommunication, particularly in marine ecosystems. Traditionally, QS has been associated with the regulation of bacterial bioluminescence. However, a study by Tanet et al. [24] presented a challenge to this conventional understanding. The researchers analyzed the bioluminescent strain Photobacterium phosphoreum ANT-2200, originating from deep-sea waters of the Mediterranean, and found it exhibited higher light emission rates at lower cell densities rather than higher ones. Additionally, high hydrostatic pressure did not affect QS gene transcription in this bacteria. Consequently, these findings suggest that in this particular strain, bioluminescence regulation may not be density-dependent and could be independent of conventional QS control mechanisms. Quorum sensing (QS) has been discovered in diverse bacterial species and habitats, including marine ecosystems. N-acyl homoserine lactones typically serve as signaling molecules in quorum sensing for gram-negative bacteria, whereas gram-positive bacteria generally employ autoinducing peptides for intercellular communication. However, these broad categories are not all-encompassing. Numerous bacterial species are known to produce unique autoinducers, underlining the diversity and specificity inherent in bacterial communication systems [25][26]. In marine environments, the genera belonging to Proteobacteria that commonly produce autoinducers (Ais) predominantly include Pseudoalteromonas, Thalassomonas, Pseudomonas, Roseobacter, Aeromonas, and Vibrio. QS has also been identified in Epsilonproteobacteria, some of which are human pathogens. The thermal origin of this ability in mesophilic and pathogenic Epsilonproteobacteria has been traced back to ocean hydrothermal vents [26][27]. Among the different classes of signaling molecules, N-acyl homoserine lactones (AHLs), oligopeptides, and LuxS/autoinducer 2 (AI-2) have been extensively studied in marine environments [27][28]. For instance, some marine bacteria, such as Vibrio species, have been shown to employ multiple QS systems simultaneously, indicating a complex hierarchical organization of QS regulation that might differ from most terrestrial bacteria. Additionally, some marine bacteria are known to produce unique QS signal molecules, such as the boronated autoinducer AI-2 (BAI-2) in Vibrio harveyi, which is a derivative of the more common AI-2 signal and is believed to be more stable in the marine environment. Vibrio harveyi, a Gram-negative bacterium renowned for its bioluminescence, thrives predominantly in marine settings as a free-living organism. However, its versatility extends to engaging symbiotic and pathogenic relationships with diverse marine creatures [28][29]. The crux of V. harveyi’s ability to interact with its hosts lies in its quorum sensing system, a regulatory mechanism controlling its bioluminescence, biofilm development, and virulence factor expression. This complex system, which encompasses multiple signaling molecules, enables V. harveyi to adapt and react to changes within its host environment and microbial community. In the quorum sensing (QS) network of Vibrio harveyi, three autoinducers (Ais) are used, which differ based on whether they are for intra-species, intra-genera, or inter-species communication [29][30]. Another common QS network structure is observed in the Pseudomonas genus, particularly in Pseudomonas aeruginosa. In marine ecosystems, P. aeruginosa has been isolated from various niches, including coastal waters, marine sediments, and marine organisms [30][31]. It is known for its robust biofilm formation capabilities, enabling it to survive in challenging marine conditions. This bacterium possesses four known QS pathways that function independently or in a coordinated manner. Two of these pathways are of the LuxI/LuxR type, specifically the LasI/LasR and RhlI/RhlR systems. Additionally, P. aeruginosa uses the quinolone-based QS system (PQS, which uses the 2-heptyl3-hydroxy-4-quinolone signal) and more recently identified integrated QS system (IQS, utilizing the 2-(2-hydroxyphenyl)-thiazole-4-carbaldehyde signal). These QS circuits are arranged hierarchically (Figure 1) [31][32].Figure 1. An illustration of the working system of the autoinducer mechanism in Gram-negative (Pseudomonas aeruginosa). The bacteria secrete AHLs (red dots) that, in threshold concentrations, penetrate the cells, activate the AHL receptor, and induce the QS-regulated gene expression. ‘I’ represents the LuxI gene encoding for the enzyme that synthesizes the AHL signal, and ‘R’ represents the LuxR gene encoding for the protein that binds AHL and modulates the transcription of luciferase genes, quinolone-based QS system (Pqs). (Image created in Bio-render online platform).