Nerve cell death accounts for various neurodegenerative disorders, in which altered immunity to the integrated central nervous system (CNS) might have destructive consequences. This undesirable immune response often affects the progressive neurodegenerative diseases such as Alzheimer’s disease, Parkinson’s disease, schizophrenia and/or amyotrophic lateral sclerosis (ALS). It has been shown that commensal gut microbiota could influence the brain and/or several machineries of immune function. In other words, neurodegenerative disorders may be connected to the gut–brain–immune correlational system. The engrams in the brain could retain the information of a certain inflammation in the body which might be involved in the pathogenesis of neurodegenerative disorders. Tactics involving the use of probiotics and/or fecal microbiota transplantation (FMT) are now evolving as the most promising and/or valuable for the modification of the gut–brain–immune axis.
- gut microbiota
- engram
- neurodegenerative disorders
- Alzheimer’s disease
- Parkinson’s disease
- amyotrophic lateral sclerosis
- schizophrenia
- inflammation
- reactive oxygen species
1. Introduction
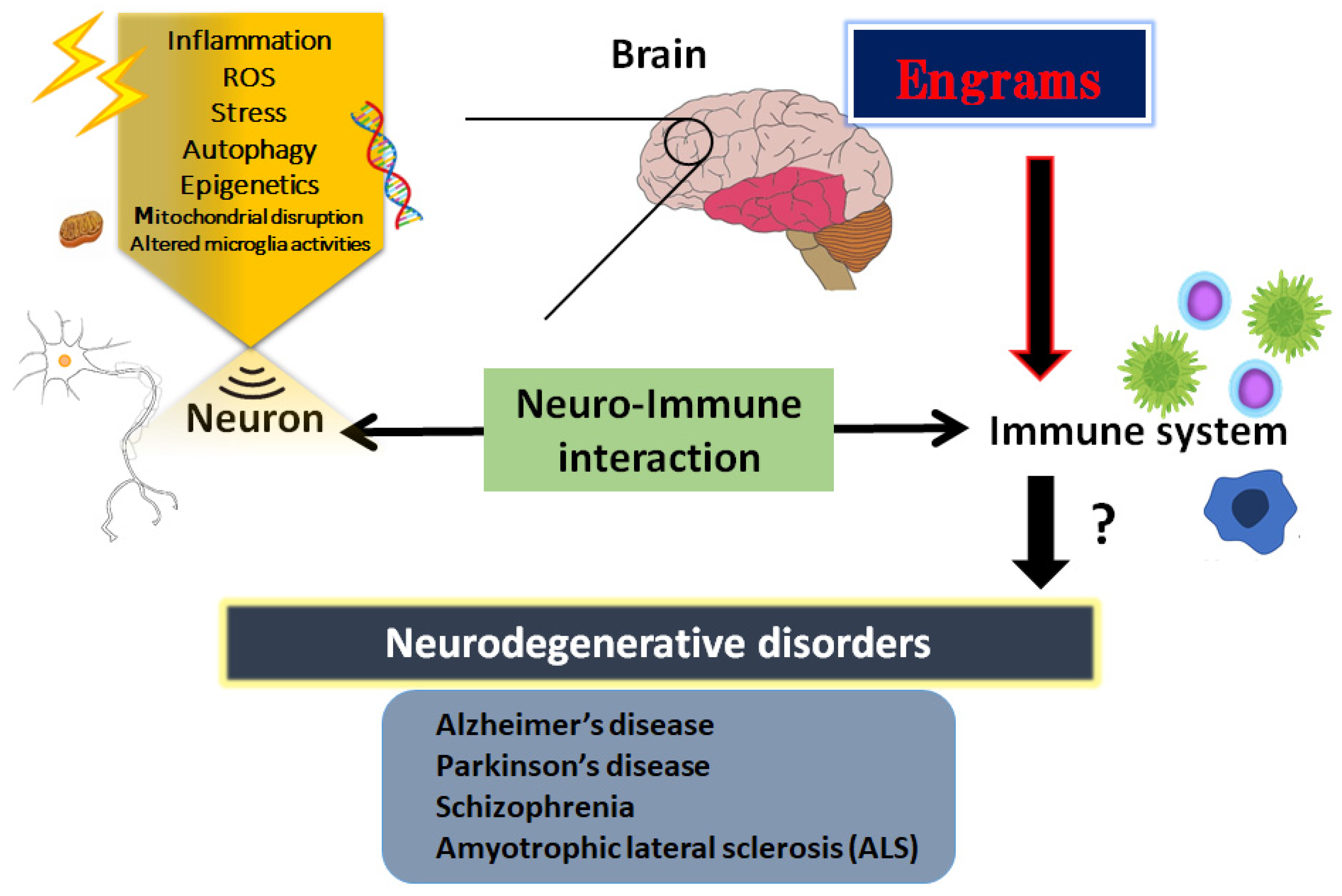
2. Inflammatory Neuro-Immune Responses
Inflammatory progression has a key role in various cellular processes and is suggested as the pathogenesis of neurodegenerative disorders [21][18]. Consistently, it has been revealed that neuro-inflammation triggered by bacterial or viral infections could induce schizophrenia in animal models [22][19]. In addition, it has been described as a reciprocal functional mechanism between the immune system and CNS [23][20]. For example, immune cells could modulate behavior and cognition of the host by direct interactions with the CNS [24][21]. A low-grade neuro-immune/inflammatory response is essential to keep the neurogenesis and/or the homeostasis of brain [5[5][7][22],7,25], suggesting that mild transient immune response might be employed as a restorative role in CNS. Consequently, an array of neuro-immune aberrations related to the chronic activated inflammatory reaction have been identified in patients with neurodegenerative disorders including schizophrenia [25][22]. Generally, an elevated level of inflammation markers in the blood and/or in cerebrospinal fluid (CSF) of the CNS has been detected in patients with neurodegenerative disorders [26][23]. Therefore, prospective treatment with anti-inflammatory medication has been suggested as a secondary treatment in patients with neurodegenerative disorders including schizophrenia or ALS [27][24]. It has been shown that extra prolonged stresses may be a robust risk factor for the development of some psychiatric diseases with a reduced number of mitochondria in the cortex [28][25]. Inflammatory oxidative stress may produce an excess amount of ROS which could be characterized as oxygen-comprising small molecules prone to react with several biological materials such as DNA [29][26]. In addition, an excess amount of ROS production could initiate an activation of autophagy in cells, suggesting an essential role for ROS in the activation of autophagy [30][27]. Generally, autophagy would play a protective role in cells; however, autophagy is also related to apoptotic cell death or necrosis in certain conditions. Additionally, autophagy could regulate the levels of several inflammations [31][28]. Hence, autophagy might be involved in the pathogenesis of neurodegenerative disorders. The significant effect of autophagy may be determined by the type of stimulus, cell types, the microenvironment, and/or other biological factors [32][29]. In intracellular signaling pathways, autophagy could be stimulated by the activated AMP-activated protein kinase (AMPK) during the situation of energy deficit in cells [33][30]. The activity of AMPK is also critical in the cells of the CNS for preserving neuronal integrity and for neuron survival against an excess amount of oxidative stresses [34][31].3. Engrams and Neuro-Immune Responses in the Pathogenesis of Neurodegenerative Disorders
The CNS and the immune system might collaborate on various levels in a body; however, the mechanisms of holding the specific immune-challenge have remained vague. Very lately, it has been clearly shown that the brain keeps the facts of certain inflammation such as inflammatory bowel syndrome occurred in the body [37][32]. This specific memory seems to be an immunological remembrance called “engrams” [38][33]. Here, wthe researchers would like to use this word “engrams” as the meaning of immunological remembrance matching to the meaning of “memory-traces”. The concept of engrams has been fairly hypothetical for the basic units of memory. Now, neuronal assemblies that hold the specific disease engrams have been known in the amygdala, hippocampus, and/or cortex, which may suggest that engrams are distributed among multiple brain regions functionally linking each other as an integrated engrams organization [39][34]. Associations of these engrams are thought to determine the situation of the host, either of health or disease, by engram arrangements, which may be frequently dependent on several environmental conditions [40][35]. Consequently, the immunological engrams could restore the initial inflammatory disease condition, if rebooted [38][33]. Created by stressful and/or repetitive inflammatory occasions, the engrams might commit to a slow progression of chronic diseases including neurodegenerative disorders [41][36]. Epigenetic changes such as DNA methylation or acetylation within the cells of the neuronal assemblies might be important mechanisms of the engram formation [42][37], which is also a significant factor for the fine-tuning of the function in the healthy brain [43][38]. Epigenetics may also stabilize engrams for the effective recovery of fear memory [44][39]. Therefore, engrams and/or epigenetic changes could be related to the immune consequences in the pathogenesis of various neurodegenerative disorders [45][40] (Figure 1). The synergistic arrangement of engrams might bring in the solid progression of several diseases, which involves the concept that any complex neurological and/or immunological consequences could result from the interaction of these engrams with immunity. Additionally, frequent subtle immune challenges might result in the stable formation of multiple engrams positioning independent information [46][41]. Synaptic variations might validate the specific development of engrams during memorizing for supporting memory. Maintenance of the memory might be achieved by a meta-plasticity mechanism that raises the change in neurons within an engram, which may be further encouraged by epigenetic regulators such as histone deacetylases (HDACs) [47][42]. In fact, the HDACs-related signaling pathways have been significantly associated with the alternative expression of several genes related to neurodegenerative disorders [48][43]. Consistently, some kinds of epigenetic regulators modified by environmental factors have been suggested as playing a crucial role in the pathogenesis of various neurodegenerative disorders [49][44]. In short, the brain could hold several specific inflammatory responses as information of pathological neuronal images called “engrams”. This concept could correctly elucidate the pathogenesis of various neurodegenerative disorders and the related CNS disorders, which might contribute to establishing a new strategy for the therapeutic interventions.4. How to Modulate the Engrams
Some engrams could potentially trigger and/or exacerbate the conditions of neurodegenerative disorders [50][45]. Therefore, clearing the bad memory of “engrams” might be favorable for the prevention and/or treatment of neurodegenerative disorders. In the experiment of dextran sulfate sodium-induced colitis, the authors applied the chemo-genetic procedure of the designer receptor exclusively activated by designer drugs (DREADD) system for the inhibition of engram activity [37][32]. However, it seems to be impossible to currently use this system in the clinical treatment of humans. Now, is it possible to clear the memory of engrams without neuronal cell death and/or any brain damage? This is the point for therapeutic interventions. In one possible way, synaptic removal could be achieved by microglia capable of initiating the oblivion of memories with engram cells [51][46]. It is considered that microglia can make synapse elimination a mechanism for forgetting memory retentions [51][46]. In addition, it has been reported that microglia are related to synapse density, learning, and/or memory [52][47]. There are significant associations between gut microbiota and demyelination by the microglia in the brain, suggesting that the crosstalk of gut-microbiota and brain-microglia might play a key role for the clearance of engrams [53][48]. It has been shown that regulation of the microbiota might be connected to the possible therapies of neurodegenerative disorders [54][49]. A gut–brain axis indicates a bidirectional connection between gut microbiota and brain, which is a vital assembly in the pathophysiology of several neurodegenerative disorders [55][50]. This concept might include the associations between gut microbiota and more broad CNS disorders. For example, it has been shown that the composition of gut microbiota might be associated with narcolepsy type 1 [56][51]. Changes in the conformation of gut microbiota may be accepted by the sympathetic vagal afferent nerve transmitting to the CNS via the microglial action, which in turn could produce and/or modulate the responses of engrams. Studies have proven that some species of bacteria could produce catecholamines and/or acetylcholine, which might contribute to the responses of the sympathetic nerve [57][52].5. Utilization of Gut–Brain Axis for the Treatment of Neurodegenerative Disorders
The dynamic residency of microbes in the gut may play a fundamental role in managing host physiology. In addition, recent advances have emphasized the significance of gut microbiota in neurodevelopment with considerable associations with the onset and/or the progression of neurodegenerative disorders [68,69][53][54]. Furthermore, it has been shown that the dysbiosis of gut microbiota might worsen the symptoms of various neurodegenerative disorders [70][55]. Alterations in the composition of gut microbiota, termed gut dysbiosis, with an increased number of potentially pathological organisms might play a prominent role in the pathogenesis of CNS-related disorders. For example, ALS patients may often demonstrate some changes in their gut microbial communities compared to the paired healthy controls [71][56]. Furthermore, increasing gut dysbiosis has been shown to worsen the symptoms with ALS [72][57]. Evolving evidence also connects the gut dysbiosis to the exacerbation of impaired autophagy in the immune-mediated chronic neuroinflammation [73][58]. Interestingly, it has been reported that a pleiotropic drug modulating AMPK and/or autophagy signaling, such as metformin, could alter the gut microbiota and its metabolic processes [74][59]. Consequently, dietary approach to alter the gut microbiota could be advantageous for the treatment of neurodegenerative disorders [75][60]. Gut microbiota could regulate and/or inhibit the production of ROS to retain the host’s brain health [76][61]. It might be important to diminish the levels of ROS for neuroregeneration with neuronal stem cells [77,78][62][63]. In addition to the unfavorable effects for the stem cells, ROS might skew the function of microglia with the oxidized mitochondria in glial cells (Figure 1) [79][64]. Inflammatory factors, oxidative stress, and/or the alteration of microglia are known to limit neuroplasticity in the CNS [80][65]. In these ways, certain gut microbiota with the inhibition of ROS could probably prevent the incidence and/or attenuate the symptoms of neurodegenerative disorders by regulating the production of ROS and by clearing engram memory via the alteration of functional microglia in the brain (Figure 2).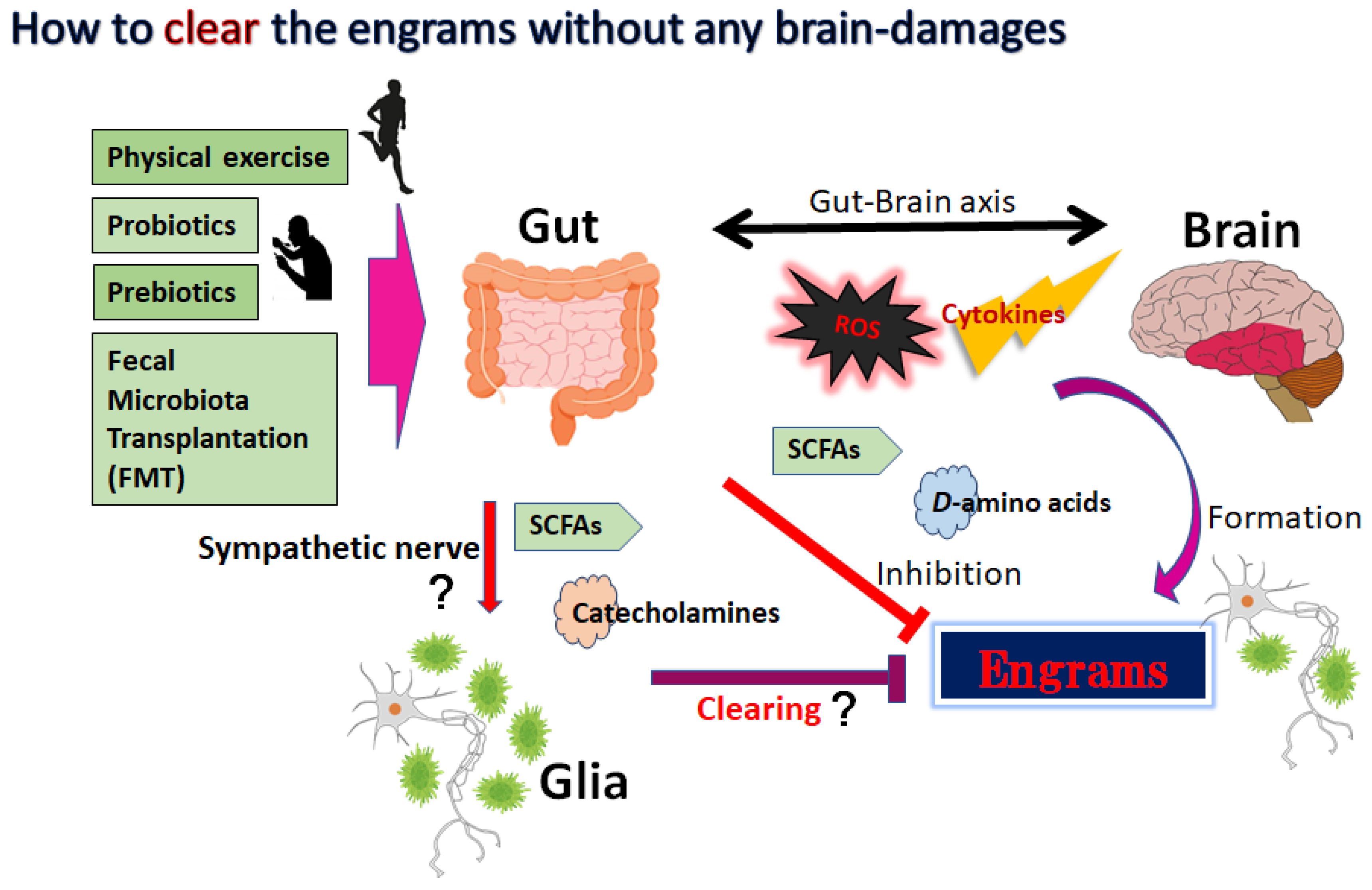
References
- Heneka, M.T.; McManus, R.M.; Latz, E. Inflammasome signalling in brain function and neurodegenerative disease. Nat. Rev. Neurosci. 2018, 19, 610–621.
- Gilodi, M.; Lisi, S.; Dudás, E.F.; Fantini, M.; Puglisi, R.; Louka, A.; Marcatili, P.; Cattaneo, A.; Pastore, A. Selection and Modelling of a New Single-Domain Intrabody Against TDP-43. Front. Mol. Biosci. 2022, 8, 773234.
- Ikeda, Y.; Nagase, N.; Tsuji, A.; Kitagishi, Y.; Matsuda, S. Neuroprotection by dipeptidyl-peptidase-4 inhibitors and glucagon-like peptide-1 analogs via the modulation of AKT-signaling pathway in Alzheimer’s disease. World. J. Biol. Chem. 2021, 12, 104–113.
- Ogino, M.; Ichimura, M.; Nakano, N.; Minami, A.; Kitagishi, Y.; Matsuda, S. Roles of PTEN with DNA Repair in Parkinson’s Disease. Int. J. Mol. Sci. 2016, 17, 954.
- Kitagishi, Y.; Nakanishi, A.; Ogura, Y.; Matsuda, S. Dietary regulation of PI3K/AKT/GSK-3β pathway in Alzheimer’s disease. Alzheimers. Res. Ther. 2014, 6, 35.
- Abbaszadeh, F.; Fakhri, S.; Khan, H. Targeting apoptosis and autophagy following spinal cord injury: Therapeutic approaches to polyphenols and candidate phytochemicals. Pharmacol. Res. 2020, 160, 105069.
- Matsuda, S.; Ikeda, Y.; Murakami, M.; Nakagawa, Y.; Tsuji, A.; Kitagishi, Y. Roles of PI3K/AKT/GSK3 Pathway Involved in Psychiatric Illnesses. Diseases 2019, 7, 22.
- Kitagishi, Y.; Minami, A.; Nakanishi, A.; Ogura, Y.; Matsuda, S. Neuron membrane trafficking and protein kinases involved in autism and ADHD. Int. J. Mol. Sci. 2015, 16, 3095–3115.
- Singh, A.; Kukretim, R.; Saso, L.; Kukreti, S. Mechanistic Insight into Oxidative Stress-Triggered Signaling Pathways and Type 2 Diabetes. Molecules 2022, 27, 950.
- Hitzeroth, A.; Niehaus, D.J.; Koen, L.; Botes, W.C.; Deleuze, J.F.; Warnich, L. Association between the MnSOD Ala-9Val polymorphism and development of schizophrenia and abnormal involuntary movements in the Xhosa population. Prog. Neuropsychopharmacol. Biol. Psychiatry 2007, 31, 664–672.
- Le Gall, L.; Anakor, E.; Connolly, O.; Vijayakumar, U.G.; Duddy, W.J.; Duguez, S. Molecular and Cellular Mechanisms Affected in ALS. J. Pers. Med. 2020, 10, 101.
- Godoy, J.A.; Rios, J.A.; Picón-Pagès, P.; Herrera-Fernández, V.; Swaby, B.; Crepin, G.; Vicente, R.; Fernández-Fernández, J.M.; Muñoz, F.J. Mitostasis, Calcium and Free Radicals in Health, Aging and Neurodegeneration. Biomolecules 2021, 11, 1012.
- Lee, K.H.; Cha, M.; Lee, B.H. Neuroprotective Effect of Antioxidants in the Brain. Int. J. Mol. Sci. 2020, 21, 7152.
- Fakhri, S.; Abbaszadeh, F.; Jorjanim, M. On the therapeutic targets and pharmacological treatments for pain relief following spinal cord injury: A mechanistic review. Biomed. Pharmacother. 2021, 139, 111563.
- Zhong, S.R.; Kuang, Q.; Zhang, F.; Chen, B.; Zhong, Z.G. Functional roles of the microbiota-gut-brain axis in Alzheimer’s disease: Implications of gut microbiota-targeted therapy. Transl. Neurosci. 2021, 12, 581–600.
- Dinan, T.G.; Cryan, J.F. Gut instincts: Microbiota as a key regulator of brain development, ageing and neurodegeneration. J. Physiol. 2017, 595, 489–503.
- Bruce-Keller, A.J.; Salbaum, J.M.; Berthoud, H.R. Harnessing Gut Microbes for Mental Health: Getting from Here to There. Biol. Psychiatry 2018, 83, 214–223.
- Noss, M.M.; Millwood, S.N.; Kuhlman, K.R. Women with lower systemic inflammation demonstrate steeper cognitive decline with age: Results from a large prospective, longitudinal sample. Brain. Behav. Immun. Health 2022, 22, 100465.
- Arsenault, D.; Coulombe, K.; Zhu, A.; Gong, C.; Kil, K.E.; Choi, J.K.; Poutiainen, P.; Brownell, A.L. Loss of Metabotropic Glutamate Receptor 5 Function on Peripheral Benzodiazepine Receptor in Mice Prenatally Exposed to LPS. PLoS ONE 2015, 10, e0142093.
- Chavan, S.S.; Pavlov, V.A.; Tracey, K.J. Mechanisms and Therapeutic Relevance of Neuro-immune Communication. Immunity 2017, 46, 927–942.
- Clark, S.M.; Vaughn, C.N.; Soroka, J.A.; Li, X.; Tonelli, L.H. Neonatal adoptive transfer of lymphocytes rescues social behaviour during adolescence in immune-deficient mice. Eur. J. Neurosci. 2018, 47, 968–978.
- Theodoropoulou, S.; Spanakos, G.; Baxevanis, C.N.; Economou, M.; Gritzapis, A.D.; Papamichail, M.P.; Stefanis, C.N. Cytokine serum levels, autologous mixed lymphocyte reaction and surface marker analysis in never medicated and chronically medicated schizophrenic patients. Schizophr. Res. 2001, 47, 13–25.
- Lupaescu, A.V.; Iavorschi, M.; Covasa, M. The Use of Bioactive Compounds in Hyperglycemia- and Amyloid Fibrils-Induced Toxicity in Type 2 Diabetes and Alzheimer’s Disease. Pharmaceutics 2022, 14, 235.
- Chang, M.C.; Kwak, S.G.; Park, J.S.; Park, D. The effectiveness of nonsteroidal anti-inflammatory drugs and acetaminophen in reduce the risk of amyotrophic lateral sclerosis? A meta-analysis. Sci. Rep. 2020, 10, 14759.
- Csabai, D.; Sebők-Tornai, A.; Wiborg, O.; Czéh, B. A Preliminary Quantitative Electron Microscopic Analysis Reveals Reduced Number of Mitochondria in the Infralimbic Cortex of Rats Exposed to Chronic Mild Stress. Front. Behav. Neurosci. 2022, 16, 885849.
- Karmakar, J.; Mukherjee, K.; Mandal, C. Siglecs Modulate Activities of Immune Cells Through Positive and Negative Regulation of ROS Generation. Front. Immunol. 2021, 12, 758588.
- Scherz-Shouval, R.; Elazar, Z. Regulation of autophagy by ROS: Physiology and pathology. Trends. Biochem. Sci. 2011, 36, 30–38.
- Zhu, L.; Liu, L. New Insights into the Interplay Among Autophagy, the NLRP3 Inflammasome and Inflammation in Adipose Tissue. Front. Endocrinol. 2022, 13, 739882.
- Marcucci, F.; Bellone, M.; Caserta, C.A.; Corti, A. Pushing tumor cells towards a malignant phenotype: Stimuli from the microenvironment, intercellular communications and alternative roads. Int. J. Cancer. 2014, 135, 1265–1276.
- Yan, L.S.; Zhang, S.F.; Luo, G.; Cheng, B.C.; Zhang, C.; Wang, Y.W.; Qiu, X.Y.; Zhou, X.H.; Wang, Q.G.; Song, X.L.; et al. Schisandrin B mitigates hepatic steatosis and promotes fatty acid oxidation by inducing autophagy through AMPK/mTOR signaling pathway. Metabolism 2022, 131, 155200.
- Nagy, S.; Maurer, G.W.; Hentze, J.L.; Rose, M.; Werge, T.M.; Rewitz, K. AMPK signaling linked to the schizophrenia-associated 1q21.1 deletion is required for neuronal and sleep maintenance. PLoS Genet. 2018, 14, e1007623.
- Koren, T.; Yifa, R.; Amer, M.; Krot, M.; Boshnak, N.; Ben-Shaanan, T.L.; Azulay-Debby, H.; Zalayat, I.; Avishai, E.; Hajjo, H.; et al. Insular cortex neurons encode and retrieve specific immune responses. Cell 2021, 184, 5902–5915.e17.
- Gogolla, N. The brain remembers where and how inflammation struck. Cell 2021, 184, 5851–5853.
- Roy, D.S.; Park, Y.G.; Kim, M.E.; Zhang, Y.; Ogawa, S.K.; DiNapoli, N.; Gu, X.; Cho, J.H.; Choi, H.; Kamentsky, L.; et al. Brain-wide mapping reveals that engrams for a single memory are distributed across multiple brain regions. Nat. Commun. 2022, 13, 1799.
- Sakaguchi, M.; Hayashi, Y. Catching the engram: Strategies to examine the memory trace. Mol. Brain. 2012, 5, 32.
- Gebicke-Haerter, P.J. Engram formation in psychiatric disorders. Front. Neurosci. 2014, 8, 118.
- Fuentes-Ramos, M.; Alaiz-Noya, M.; Barco, A. Transcriptome and epigenome analysis of engram cells: Next-generation sequencing technologies in memory research. Neurosci. Biobehav. Rev. 2021, 127, 865–875.
- Bachmann, S.; Linde, J.; Bell, M.; Spehr, M.; Zempel, H.; Zimmer-Bensch, G. DNA Methyltransferase 1 (DNMT1) Shapes Neuronal Activity of Human iPSC-Derived Glutamatergic Cortical Neurons. Int. J. Mol. Sci. 2021, 22, 2034.
- Gulmez Karaca, K.; Kupke, J.; Brito, D.V.C.; Zeuch, B.; Thome, C.; Weichenhan, D.; Lutsik, P.; Plass, C.; Oliveira, A.M.M. Neuronal ensemble-specific DNA methylation strengthens engram stability. Nat. Commun. 2020, 11, 639.
- Niemi, M.B.; Härting, M.; Kou, W.; Del Rey, A.; Besedovsky, H.O.; Schedlowski, M.; Pacheco-López, G. Taste-immunosuppression engram: Reinforcement and extinction. J. Neuroimmunol. 2007, 188, 74–79.
- Pacheco-López, G.; Niemi, M.B.; Kou, W.; Baum, S.; Hoffman, M.; Altenburger, P.; del Rey, A.; Besedovsky, H.O.; Schedlowski, M. Central blockade of IL-1 does not impair taste-LPS associative learning. Neuroimmunomodulation 2007, 14, 150–156.
- Kyrke-Smith, M.; Williams, J.M. Bridging Synaptic and Epigenetic Maintenance Mechanisms of the Engram. Front. Mol. Neurosci. 2018, 11, 369.
- Manea, S.A.; Vlad, M.L.; Fenyo, I.M.; Lazar, A.G.; Raicu, M.; Muresian, H.; Simionescu, M.; Manea, A. Pharmacological inhibition of histone deacetylase reduces NADPH oxidase expression, oxidative stress and the progression of atherosclerotic lesions in hypercholesterolemic apolipoprotein E-deficient mice; potential implications for human atherosclerosis. Redox Biol. 2020, 28, 101338.
- Qing, L.; Liu, L.; Zhou, L.; Zhang, F.; Gao, C.; Hu, L.; Nie, S. Sex-dependent association of mineralocorticoid receptor gene (NR3C2) DNA methylation and schizophrenia. Psychiatry Res. 2020, 292, 113318.
- Bostancıklıoğlu, M. An update on memory formation and retrieval: An engram-centric approach. Alzheimers. Dement. 2020, 16, 926–937.
- Wang, C.; Yue, H.; Hu, Z.; Shen, Y.; Ma, J.; Li, J.; Wang, X.D.; Wang, L.; Sun, B.; Shi, P.; et al. Microglia mediate forgetting via complement-dependent synaptic elimination. Science 2020, 367, 688–694.
- Wang, Y.Y.; Deng, Y.S.; Dai, S.K.; Mi, T.W.; Li, R.Y.; Liu, P.P.; Liu, C.; He, B.D.; He, X.C.; Du, H.Z.; et al. Loss of microglial EED impairs synapse density, learning, and memory. Mol. Psychiatry 2022, 27, 2999–3009.
- Wang, X.; Chang, L.; Wan, X.; Tan, Y.; Qu, Y.; Shan, J.; Yang, Y.; Ma, L.; Hashimoto, K. (R)-ketamine ameliorates demyelination and facilitates remyelination in cuprizone-treated mice: A role of gut-microbiota-brain axis. Neurobiol. Dis. 2022, 165, 105635.
- Ghezzi, L.; Cantoni, C.; Pinget, G.V.; Zhou, Y.; Piccio, L. Targeting the gut to treat multiple sclerosis. J. Clin. Investig. 2021, 131, e143774.
- Klann, E.M.; Dissanayake, U.; Gurrala, A.; Farrer, M.; Shukla, A.W.; Ramirez-Zamora, A.; Mai, V.; Vedam-Mai, V. The Gut-Brain Axis and Its Relation to Parkinson’s Disease: A Review. Front. Aging Neurosci. 2022, 13, 782082.
- Lecomte, A.; Barateau, L.; Pereira, P.; Paulin, L.; Auvinen, P.; Scheperjans, F.; Dauvilliers, Y. Gut microbiota composition is associated with narcolepsy type 1. Neurol. Neuroimmunol. Neuroinflamm. 2020, 7, e896.
- Wiley, N.C.; Cryan, J.F.; Dinan, T.G.; Ross, R.P.; Stanton, C. Production of Psychoactive Metabolites by Gut Bacteria. Mod. Trends Psychiatry 2021, 32, 74–99.
- Caputi, V.; Popov, J.; Giron, M.C.; O’Mahony, S. Gut Microbiota as a Mediator of Host Neuro-Immune Interactions: Implications in Neuroinflammatory Disorders. Mod. Trends. Psychiatry 2021, 32, 40–57.
- Benakis, C.; Martin-Gallausiaux, C.; Trezzi, J.P.; Melton, P.; Liesz, A.; Wilmes, P. The microbiome-gut-brain axis in acute and chronic brain diseases. Curr. Opin. Neurobiol. 2020, 61, 1–9.
- Castanon, N.; Luheshi, G.; Layé, S. Role of neuroinflammation in the emotional and cognitive alterations displayed by animal models of obesity. Front. Neurosci. 2015, 9, 229.
- Hertzberg, V.S.; Singh, H.; Fournier, C.N.; Moustafa, A.; Polak, M.; Kuelbs, C.A.; Torralba, M.G.; Tansey, M.G.; Nelson, K.E.; Glass, J.D. Gut microbiome differences between amyotrophic lateral sclerosis patients and spouse controls. Amyotroph. Lateral. Scler. Frontotemporal. Degener. 2022, 23, 91–99.
- Kim, H.S.; Son, J.; Lee, D.; Tsai, J.; Wang, D.; Chocron, E.S.; Jeong, S.; Kittrell, P.; Murchison, C.F.; Kennedy, R.E.; et al. Gut- and oral-dysbiosis differentially impact spinal- and bulbar-onset ALS, predicting ALS severity and potentially determining the location of disease onset. BMC. Neurol. 2022, 22, 62.
- Chidambaram, S.B.; Essa, M.M.; Rathipriya, A.G.; Bishir, M.; Ray, B.; Mahalakshmi, A.M.; Tousif, A.H.; Sakharkar, M.K.; Kashyap, R.S.; Friedland, R.P.; et al. Gut dysbiosis, defective autophagy and altered immune responses in neurodegenerative diseases: Tales of a vicious cycle. Pharmacol. Ther. 2022, 231, 107988.
- Trujillo-Del Río, C.; Tortajada-Pérez, J.; Gómez-Escribano, A.P.; Casterá, F.; Peiró, C.; Millán, J.M.; Herrero, M.J.; Vázquez-Manrique, R.P. Metformin to treat Huntington disease: A pleiotropic drug against a multi-system disorder. Mech. Ageing Dev. 2022, 204, 111670.
- Matsuda, S.; Nakagawa, Y.; Amano, K.; Ikeda, Y.; Tsuji, A.; Kitagishi, Y. By using either endogenous or transplanted stem cells, which could you prefer for neural regeneration? Neural. Regen. Res. 2018, 13, 1731–1732.
- Taniguchi, K.; Ikeda, Y.; Nagase, N.; Tsuji, A.; Kitagishi, Y.; Matsuda, S. Implications of Gut-Brain axis in the pathogenesis of Psychiatric disorders. AIMS. Bioeng. 2021, 8, 243–256.
- Matsuda, S.; Nakagawa, Y.; Kitagishi, Y.; Nakanishi, A.; Murai, T. Reactive Oxygen Species, Superoxide Dimutases, and PTEN-p53-AKT-MDM2 Signaling Loop Network in Mesenchymal Stem/Stromal Cells Regulation. Cells 2018, 7, 36.
- Ikeda, Y.; Taniguchi, K.; Nagase, N.; Tsuji, A.; Kitagishi, Y.; Matsuda, S. Reactive oxygen species may influence on the crossroads of stemness, senescence, and carcinogenesis in a cell via the roles of APRO family proteins. Explor. Med. 2021, 2, 443–454.
- Zhang, L.; Qian, Y.; Li, J.; Zhou, X.; Xu, H.; Yan, J.; Xiang, J.; Yuan, X.; Sun, B.; Sisodia, S.S.; et al. BAD-mediated neuronal apoptosis and neuroinflammation contribute to Alzheimer’s disease pathology. iScience 2021, 24, 102942.
- Acosta, S.; Jernberg, J.; Sanberg, C.D.; Sanberg, P.R.; Small, B.J.; Gemma, C.; Bickford, P.C. NT-020, a natural therapeutic approach to optimize spatial memory performance and increase neural progenitor cell proliferation and decrease inflammation in the aged rat. Rejuvenation Res. 2010, 13, 581–588.