Early-life stress during critical periods of brain development can have long-term effects on physical and mental health. Oxytocin is a critical social regulator and anti-inflammatory hormone that modulates stress-related functions and social behaviors and alleviates diseases. Oxytocin-related neural systems show high plasticity in early postpartum and adolescent periods. Early-life stress can influence the oxytocin system long term by altering the expression and signaling of oxytocin receptors. Deficits in social behavior, emotional control, and stress responses may result, thus increasing the risk of anxiety, depression, and other stress-related neuropsychiatric diseases. Oxytocin is regarded as an important target for the treatment of stress-related neuropsychiatric disorders.
- early-life stress
- oxytocin
- neural circuit
- neuropsychiatric disorders
1. Introduction
2. The Properties of Oxytocin
Oxytocin is a neuropeptide hormone primarily synthesized in the brain by the parvocellular neurons (parvOT) of the paraventricular nucleus (PVN) and the magnocellular neurons (magnOT) of both the PVN and supraoptic nucleus (SON) [15,16,17,18][11][12][13][14]. The magnOT neurons primarily innervate the forebrain and release oxytocin through the posterior pituitary gland into the bloodstream to supply the body. The parvOT neurons release less oxytocin, mainly to brainstem nuclei, the spinal cord and amygdala, stria bed nucleus, nucleus accumbens (NAc), magnOT neurons of the SON, and other regions [19,20,21][15][16][17]. The oxytocin gene encodes the structural precursor to oxytocin and is expressed in the mammalian hypothalamus; oxytocin is stored in large, dense-core vesicles [22,23][18][19]. The axonal projections of PVN and SON pass through the median eminence and innervate the posterior pituitary lobe, thereby releasing OXT into circulation or through volume transfer to nerve tissue to regulate physiology [24][20]. Oxytocin works through its receptor, OXTR [29[21][22],30], which belongs to the GPCR family and can activate several different G-protein-induced signaling cascades [31,32][23][24]. OXTR activation has been associated with two separate intracellular signaling cascades with dependence on either: Gαi/o or Gαq [28,32][24][25]. OXTR-expressing neurons have different functionality in different brain regions: social reward in the ventral tegmental area (VTA) [33][26], social recognition in the anterior olfactory nucleus (AON) [34][27], and social memory in the hippocampal CA2 region [35][28]. OXTRs are distributed in neurons, astrocytes, and microglia [32,36][24][29]. Neuronal oxytocin signaling is primarily influenced by the amount of locally released oxytocin, OXTR affinity, density, local enzymatic cleavage, and the resulting concentration of oxytocin in the extracellular fluid [28][25]. Oxytocin treatment reduces inflammation and the severity of various diseases. OXTRs have been found on immune cells, including neutrophils, macrophages, and lymphocytes [47][30]. During inflammation, nuclear factor kappa-light-chain-enhancer (NF-κB) mediates increased OXTR expression in macrophages [48][31]. Oxytocin can inhibit the macrophage transition to active inflammatory cells by promoting the expression of β-arrestin 2 [47][30] and peroxisome proliferator-activated receptor gamma [49,50][32][33].3. Oxytocin-Involved Social Behavior
3.1. Positive Social Interactions
Oxytocin signaling is mediated by release from afferent terminals to receptors present in various target regions that impact aspects of behavior. Oxytocin modulates a variety of positive social behaviors via OXTRs in many limbic and reward-related regions of the mammalian brain, such as the PFC, NAc, amygdala, lateral septum, and thalamus [54][34]. The VTA and the NAc of the limbic system of the midbrain cortex are abundant in OXTRs and are linked to dopaminergic reward motivation [33][26]. The VTA PVN neurons release oxytocin to dopamine-secreting neurons of the nerve bundles that stretch from the VTA to the NAc, thus strengthening the social abilities of these mice [33,55][26][35]. Oxytocin may also increase the excitatory drive of VTA dopamine neurons that project to the NAc, thus improving sociability and social pleasure and increasing 5-HT release in the NAc [33,56,57,58][26][36][37][38]. The potent monoamine releaser (±) 3,4-methylenedioxymethamphetamine (MDMA) has potent prosocial effects in humans. Beyond social reward behavior, oxytocin also regulates prosocial behavior and the parent-child relationship. Countless studies have shown that oxytocin signaling regulates social interactions, especially prosocial behaviors associated with altruism [60][39]. MagnOT neurons have long-distance axonal projections to forebrain regions, including the PFC, AON, NAc, lateral septum, hippocampus, and medial and central amygdala, which are found only in higher vertebrates such as mammals and reptiles. These co-evolved regions are implicated in complex social and emotional behaviors [28][25]. In social mammals, OXT mediates prosocial behaviors such as mate preference, social approval and proximity, and parental care, and this effect is more pronounced in monogamous mating systems [61][40]. Social monogamy is a mating system characterized by sharing of territory among partners, mutual care, and preferential mating [62][41]. Neurobiological research on animal social bonds has focused on “social monogamous” species because they have strong long-term bonds, and the formation of this social bond seems to involve oxytocin [63][42]. The density of OXTR is higher in the NAc, mPFC, and amygdala of monogamous meadow voles than in non-monogamous montane (Microtus montanus) and meadow (Microtus pennsylvanicus) voles [62,64][41][43]. OXTR mRNA expression is present in the NAc of humans but absent from non-monogamous rhesus monkeys [62,65,66][41][44][45], and oxytocin is activated in the ACC and amygdala when rats engage in helping behaviors [67][46]. Oxytocin has a crucial role in complex social behaviors such as generosity, empathy, and collaboration [78][47]. Wild-type zebrafish freeze when observing a conspecific in pain in an isolated tank, whereas mutant zebrafish lineages lacking OXT or OXTR or do not exhibit these fear responses, indicating that oxytocin is necessary and sufficient for social fear contagion in zebrafish [79][48]. Interest in the role of oxytocin in social cognition and emotional processing increased after a study found that an oxytocin nasal spray promotes trust and play in social situations [80][49]. Complex social behaviors are closely connected with analgesia, and the role of oxytocin in analgesia and fear is noteworthy. The lactation-induced and oxytocin-dependent lack of social dread is blocked by chemogenetic suppression of DREADD-expressing OXT+ neurons that project to the lateral septum in breastfeeding mice [81][50]. The freezing caused by conventional fear training is reduced by oxytocin in the central amygdala (instead of a maternal context) [21][17]. The lateral portion of the central amygdala (CeL) receives lateral branch axons from magnOT neurons in the PVN and accessory nuclei, and these neurons create glutamatergic synapses with OXTR-expressing GABAergic neurons (Figure 21) [21][17].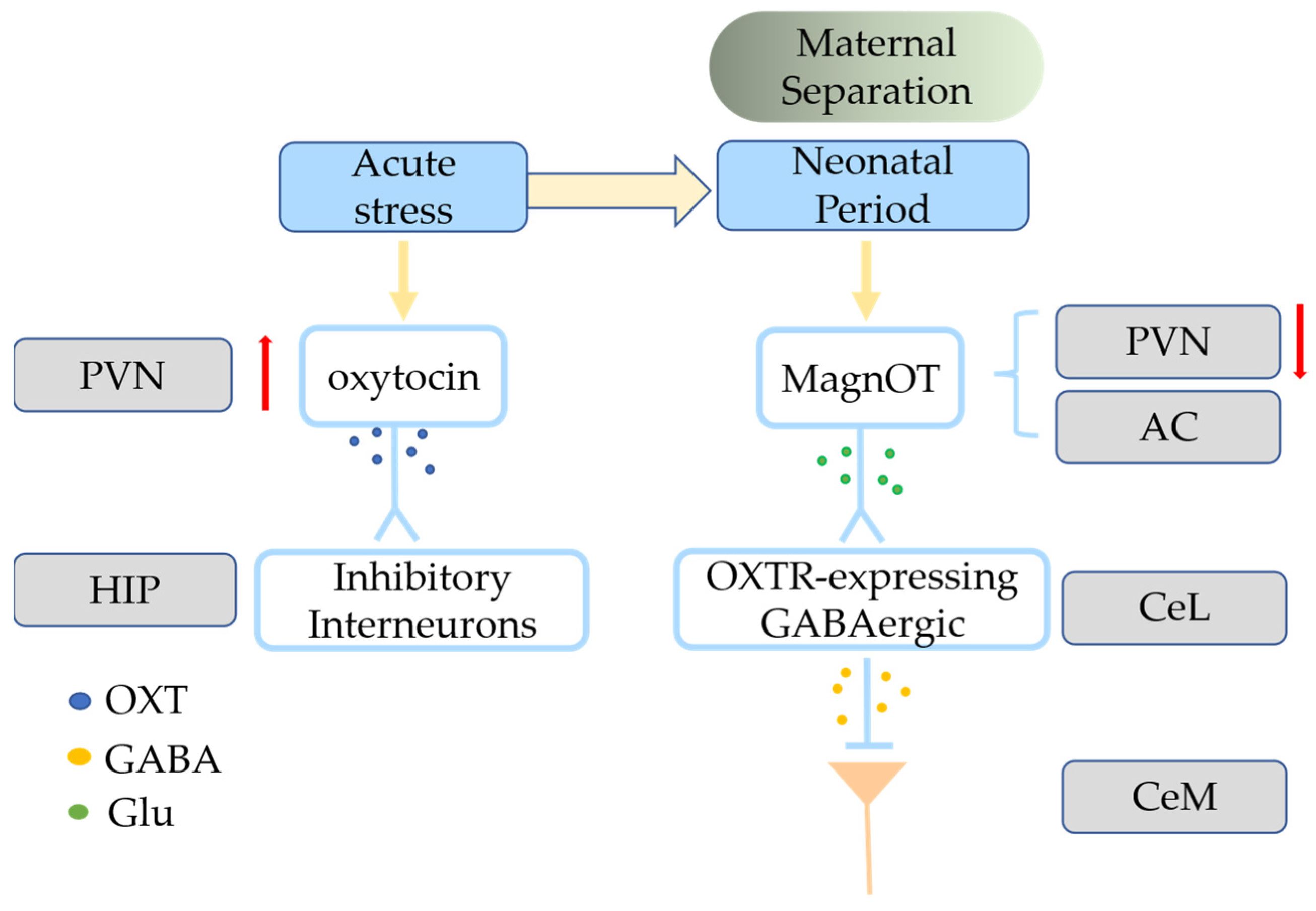
3.2. Negative Social Interactions
3.3. Sexual Dimorphism of Oxytocinin Social Behavior
Sexually dimorphic behaviors may result from sex-specific patterns of activity in the social-behavior-related brain areas. Sexual dimorphism should be considered when investigating the oxytocin system because OXTR expression is regulated by sex hormones, though there are no evident neuroanatomical differences in the distribution of OXT and OXTR expression. There have been reports describing sex-differentiated OXT+ fibers in the MPOAi of Mongolian gerbils, where females have a higher OXT+ cell density than males [96[60][61][62][63],97,98,99], and this sex difference may result from differences in axonal transport or behavioral response speed. The OXTR+ mPFC interneurons have obvious sexual dimorphism in social behavior; activation of these neurons causes anxiety in males but promotes social behavior in females [100,101][64][65].4. Effects of ELS on the Oxytocin System and Central Nervous System
ELS is a stress-induced deficit in social behavior that is closely related to limbic abnormalities that cause chronic activation of physiologic stress responses [56][36]. The term “early life” is frequently used to characterize several developmental periods, such as prenatal, early postnatal (until weaning on postnatal day 21), and puberty (postnatal day 25–35 in rodents). Experience shapes neural circuits during crucial periods of development, allowing individuals to adapt their behaviors to their surroundings in a unique way [102][66]. During this period, the nervous system is extremely sensitive to specific environmental stimuli [103][67], and this sensitivity is necessary for the normal formation of neural circuits and biological learning. In humans, the effects of ELS are similar to those in children born to mothers who experience adverse living circumstances during pregnancy, children in orphanages, and children who are abused in childhood. ELS can be studied more easily by observing animal models, and there are many ELS mouse models; common models include maternal separation/maternal deprivation and limited bedding and nesting. Maternal separation is widely used to simulate ELS through repeated separations from the mother over prolonged periods. Maternal separation can induce depression-like and anxiety-like behaviors in rodents [106,107][68][69]. The ELS pups will activate the HPA axis after long-term separation from their mothers, causing corticosterone levels to rise, and these inflammation episodes may develop over time into chronic inflammation [108][70].4.1. Effects of ELS on the Oxytocin System
The oxytocin system is crucial to social behavior, and its disturbance can have far-reaching implications. In prairie voles, it has been demonstrated that increased parental care causes hypomethylation of the oxytocin receptor gene [111][71]. Female rats raised by attentive mothers who frequently licked or groomed their paws showed significant OXTR binding [112][72]. During development, slow, gentle stroking can elicit pleasurable sensations and social rewards by activating C-tactile fibers [113][73]. This stroking also acutely activates hypothalamic OXT+ neurons and promotes OXT release, and parental care modulates hypothalamic oxytocin concentrations in rat pups [114][74]. Deprivation of touch and social interaction can lead to irreversible deficits in emotional, social, and cognitive behavior [115][75].4.2. Abnormalities in Oxytocin System Altered by ELS Correlated with Glial Cells
The complex mammalian central nervous system results, in part, from the varied cell types formed during development; oxytocin may regulate cell growth, differentiation, and contact with other cells [129][76]. ELS can cause neuroinflammation that damages the developing brain, which then enters an overactivated state that can also be induced by bacterial, environmental chemicals, or neuronal injury or death [130][77]. Astrocytes are the primary cell type implicated in the neuroinflammatory response; they account for 20–40% of all glial cells, and the brain’s balance of nutrient delivery and metabolism is dependent on astrocytes. While astrocytes contain many small fibers that penetrate into the local environment and react to various stimuli, neurons have distinctive dendrites and axons that allow long-distance projections [131][78]. While astrocyte calcium transients can last anywhere from minutes to hours, neuronal electrical activity lasts only a few milliseconds [131][78]. Optogenetic photostimulation of CeA axon terminals causes the release of OXT, which creates calcium transients in adjacent astrocytes [39][79]. In the hippocampus, mPFC, and ACC, ELS decreases the number of astrocytes [132,133][80][81]. Microglia are also important to the neuroinflammatory response; microglia secrete the pro-inflammatory cytokines IL-1, TNF-α, and C1q to activate astrocytes [134][82]. Microglia are highly sensitive to the neural environment, and although there is evidence that OXT affects microglia responsiveness in neuroinflammation, the mechanism of this influence is unclear [36][29]. Glial cells have functions beyond support; astrocytes and microglia release neuromodulators, and oligodendrocytes produce myelin that facilitates neurotransmission and neuronal oscillations [135,136,137,138][83][84][85][86].5. Potential Therapy Strategies of Oxytocin in ELS-Related Neuropsychiatric Disorders
Early-life stress can impair social behavior and contribute to a variety of stress-related diseases. Oxytocin is released into several brain regions closely associated with stress-related disorders, such as the amygdala, hypothalamus, hippocampus, and NAc. Social rewards are important for social interaction, and social reward disorder is closely related to neuropsychiatric disorders in stress-related diseases. Oxytocin is closely connected to social reward and plays an important role in stress-related neurological disorders. At present, oxytocin has been studied in numerous clinical trials of social disorders-related diseases. Dysregulation of dopaminergic signaling is associated with a variety of neuropsychiatric and neurological disorders, including ASD, Parkinson’s disease, and depression [142][87]. For these reasons, oxytocin has been investigated clinically as a treatment for these stress-induced mental disorders, though oxytocin does not easily cross the blood-brain barrier. Delivery of oxytocin has been attempted via intranasal administration, though details have not been confirmed [84][52]. Intranasal oxytocin can reach CSF and blood circulation, though it is unclear that oxytocin can reach concentrations in the brain that would produce clinically meaningful behavioral effects; microdialysis methods are questionable and have not been performed in humans. However, a recently developed mechanism enables oxytocin transport from the periphery to the central nervous system. After intranasal, subcutaneous, or intravenous administration, oxytocin level rises in the amygdala, hypothalamus, and other regions because RAGE, a membrane-associated receptor of advanced glycation end products, binds to and transports peripheral oxytocin via endothelial cells [84,144][52][88].5.1. Autism Spectrum Disorder
ASD is characterized by limitations in repetitive behavioral patterns, poor social interaction, and negligible perception. Both ASD and antisocial disorder have deficits of empathy and social cognition deficits. Because OXTR is distributed primarily in social-behavior-connected brain regions, such as the olfactory bulbs, lateral septum, and piriform cortex, oxytocin may regulate social behavior in ASD [147][89]. Children with ASD have lower plasma oxytocin levels but higher precursor levels than healthy controls, suggesting that oxytocin processing may contribute to ASD [148][90]. Autism may be connected to dysfunction of the amygdala, a major component of the cortico–striatal–thalamo–limbic system and emotional circuit involved in regulating emotional stress; oxytocin treatment can reduce amygdala activity and the fear response [82][91]. Clinical trials have examined the potential of oxytocin as a treatment for ASD, but the findings have been conflicting. In one study, 32 ASD children aged 6–12 years were given intranasal oxytocin for 4 weeks, and this treatment improved the social skills of ASD children [150][92], while a study of 16 ASD patients aged 12–19 showed that oxytocin treatment improved emotion recognition [151][93]. Though clinical studies indicate the therapeutic potential of oxytocin for social-disorder-related diseases such as ASD, the mechanism of these effects is still unclear. Social behaviors can be studied in rodent models to measure defects related to brain function and disease. Numerous animal studies have connected ASD and oxytocin, and clinical studies suggest a connection between ASD and inflammation; ASD patients have increased levels of pro-inflammatory cytokines in the brain (e.g., TNF-α, IFN-γ, and IL-6) [169][94]. In ASD animal models, microglia activation and increased peripheral and central TNF-α, IL-1β, and IL-6 have been observed [170][95], and the anti-inflammatory effects of oxytocin may play a role in this. Plasma oxytocin levels of male ASD patients are negatively correlated with the inflammation-related molecule IFN-γ-induced protein-16 [171,172][96][97]. A disorder of the central oxytocin system was found in Cntnap2 knock-out mice. A central hypothesis of ASD etiology is that long-term developmental disconnection causes abnormal resting-state functional connectivity, and this long-range disconnection may result from developmental events [178][98]. Patients with ASD have reduced functional connectivity in the cerebellum, fusiform gyrus, and occipital brain, and have lower levels of medium- and short-range functional connectivity in the posterior cingulate cortex and mPFC, indicating the distance-dependence of ASD dysfunction [179][99].5.2. Schizophrenia
Schizophrenia is a neurodevelopmental condition with genetic predispositions and an origin connected to stress during critical periods of development [181][100]; the condition has positive symptoms (delusions, hallucinations, thinking abnormalities), negative symptoms (anhedonia, sadness, social isolation, faulty thinking), and cognitive dysfunction [182][101]. Previous studies have identified oxytocinergic dysfunction in people with schizophrenia, and single-nucleotide polymorphisms in the OXT gene contribute to schizophrenia vulnerability [183][102]. Thus, there has been growing interest in the use of oxytocin as a treatment for schizophrenia [181][100]. Social cognitive dysfunction leads to exacerbated delusions, anhedonia, diminished motivation, and disengagement from social interactions, which further leads to comorbid depression [185,186,187][103][104][105]. Mice with knock-outs of either OXT [188,189,190][106][107][108] or OXTR [148,186][90][104] have deficits in social recognition, and oxytocin supplementation to the preoptic region rescues these deficits [189,191][107][109].5.3. Social Anxiety Disorders
Social anxiety disorder (SAD) is characterized by social fear, avoidance, cognitive dysfunction, and life interference, and oxytocin inhibits the amygdala’s response to fear signals, slows maladaptive cognition toward exposed tasks and reduces negative self-evaluation after social stress. In healthy subjects, intranasal administration of synthetic OXT reduces anxiety levels and broadly promotes human social behavior [199][110].6. Conclusions
Early-life stress contributes to many social disorders by altering OXT and OXTR expression in adulthood. Complex social interactions and behaviors are governed by many neural circuits and neuromodulators, and oxytocin plays a crucial role in the mother–infant relationship and stress-induced neuropsychiatric disorders. The impact of oxytocin administration depends on patient gender, brain region, dosage, and experimental paradigms for ELS. Oxytocin is mainly being considered for the treatment of ASD; studies of oxytocin treatment for stress-related neuropsychiatric disorders have produced inconsistent results, but further study of this inconsistency is warranted. Oxytocin signaling may play a more limited role than previously thought in attachment behaviors that may be too essential to rely on simplistic regulation, and other regulatory pathways may compensate for defective oxytocin signaling. This may also explain the mixed results of oxytocin treatment for stress-related neuropsychiatric disorders.References
- Schiavone, S.; Jaquet, V.; Trabace, L.; Krause, K.H. Severe life stress and oxidative stress in the brain: From animal models to human pathology. Antioxid. Redox Signal. 2013, 18, 1475–1490.
- Selye, H. A syndrome produced by diverse nocuous agents. 1936. J. Neuropsychiatry Clin. Neurosci. 1998, 10, 230–231.
- Agorastos, A.; Pervanidou, P.; Chrousos, G.P.; Baker, D.G. Developmental Trajectories of Early Life Stress and Trauma: A Narrative Review on Neurobiological Aspects Beyond Stress System Dysregulation. Front. Psychiatry 2019, 10, 118.
- Makris, G.; Eleftheriades, A.; Pervanidou, P. Early Life Stress, Hormones, and Neurodevelopmental Disorders. Horm. Res. Paediatr. 2022, 96, 70–77.
- Wright, M.O.; Crawford, E.; Del Castillo, D. Childhood emotional maltreatment and later psychological distress among college students: The mediating role of maladaptive schemas. Child. Abus. Negl. 2009, 33, 59–68.
- Baracz, S.J.; Everett, N.A.; Cornish, J.L. The impact of early life stress on the central oxytocin system and susceptibility for drug addiction: Applicability of oxytocin as a pharmacotherapy. Neurosci. Biobehav. Rev. 2020, 110, 114–132.
- Colleluori, G.; Galli, C.; Severi, I.; Perugini, J.; Giordano, A. Early Life Stress, Brain Development, and Obesity Risk: Is Oxytocin the Missing Link? Cells 2022, 11, 623.
- Love, T.M. The impact of oxytocin on stress: The role of sex. Curr. Opin. Behav. Sci. 2018, 23, 136–142.
- Lang, R.E.; Heil, J.W.; Ganten, D.; Hermann, K.; Unger, T.; Rascher, W. Oxytocin unlike vasopressin is a stress hormone in the rat. Neuroendocrinology 1983, 37, 314–316.
- García-Cáceres, C.; Balland, E.; Prevot, V.; Luquet, S.; Woods, S.C.; Koch, M.; Horvath, T.L.; Yi, C.-X.; Chowen, J.A.; Verkhratsky, A.; et al. Role of astrocytes, microglia, and tanycytes in brain control of systemic metabolism. Nat. Neurosci. 2019, 22, 7–14.
- George, J.M. Immunoreactive vasopressin and oxytocin: Concentration in individual human hypothalamic nuclei. Science 1978, 200, 342–343.
- Ohlsson, B.; Truedsson, M.; Djerf, P.; Sundler, F. Oxytocin is expressed throughout the human gastrointestinal tract. Regul. Pept. 2006, 135, 7–11.
- Althammer, F.; Grinevich, V. Diversity of oxytocin neurons: Beyond magno- and parvocellular cell types? J. Neuroendocrinol. 2017, 30, e12549.
- McCormack, S.E.; Blevins, J.E.; Lawson, E.A. Metabolic Effects of Oxytocin. Endocr. Rev. 2020, 41, 121–145.
- Swanson, L.W.; Sawchenko, P.E.; Wiegand, S.J.; Price, J.L. Separate neurons in the paraventricular nucleus project to the median eminence and to the medulla or spinal cord. Brain Res. 1980, 198, 190–195.
- Tang, Y.; Benusiglio, D.; Lefevre, A.; Hilfiger, L.; Althammer, F.; Bludau, A.; Hagiwara, D.; Baudon, A.; Darbon, P.; Schimmer, J.; et al. Social touch promotes interfemale communication via activation of parvocellular oxytocin neurons. Nat. Neurosci. 2020, 23, 1125–1137.
- Knobloch, H.S.; Charlet, A.; Hoffmann, L.C.; Eliava, M.; Khrulev, S.; Cetin, A.H.; Osten, P.; Schwarz, M.K.; Seeburg, P.H.; Stoop, R.; et al. Evoked axonal oxytocin release in the central amygdala attenuates fear response. Neuron 2012, 73, 553–566.
- Baldi, E.; Costa, A.; Rani, B.; Passani, M.B.; Blandina, P.; Romano, A.; Provensi, G. Oxytocin and Fear Memory Extinction: Possible Implications for the Therapy of Fear Disorders? Int. J. Mol. Sci. 2021, 22, 10000.
- Brownstein, M.J.; Russell, J.T.; Gainer, H. Synthesis, transport, and release of posterior pituitary hormones. Science 1980, 207, 373–378.
- Landgraf, R.; Neumann, I.D. Vasopressin and oxytocin release within the brain: A dynamic concept of multiple and variable modes of neuropeptide communication. Front. Neuroendocr. 2004, 25, 150–176.
- Dumais, K.M.; Veenema, A.H. Vasopressin and oxytocin receptor systems in the brain: Sex differences and sex-specific regulation of social behavior. Front. Neuroendocr. 2016, 40, 1–23.
- Stoop, R. Neuromodulation by oxytocin and vasopressin. Neuron 2012, 76, 142–159.
- Gimpl, G.; Fahrenholz, F. The oxytocin receptor system: Structure, function, and regulation. Physiol. Rev. 2001, 81, 629–683.
- Baudon, A.; Clauss Creusot, E.; Althammer, F.; Schaaf, C.P.; Charlet, A. Emerging role of astrocytes in oxytocin-mediated control of neural circuits and brain functions. Prog. Neurobiol. 2022, 217, 102328.
- Jurek, B.; Neumann, I.D. The Oxytocin Receptor: From Intracellular Signaling to Behavior. Physiol. Rev. 2018, 98, 1805–1908.
- Hung, L.W.; Neuner, S.; Polepalli, J.S.; Beier, K.T.; Wright, M.; Walsh, J.J.; Lewis, E.M.; Luo, L.; Deisseroth, K.; Dolen, G.; et al. Gating of social reward by oxytocin in the ventral tegmental area. Science 2017, 357, 1406–1411.
- Oettl, L.L.; Ravi, N.; Schneider, M.; Scheller, M.F.; Schneider, P.; Mitre, M.; da Silva Gouveia, M.; Froemke, R.C.; Chao, M.V.; Young, W.S.; et al. Oxytocin Enhances Social Recognition by Modulating Cortical Control of Early Olfactory Processing. Neuron 2016, 90, 609–621.
- Raam, T.; McAvoy, K.M.; Besnard, A.; Veenema, A.H.; Sahay, A. Hippocampal oxytocin receptors are necessary for discrimination of social stimuli. Nat. Commun. 2017, 8, 2001.
- Knoop, M.; Possovre, M.-L.; Jacquens, A.; Charlet, A.; Baud, O.; Darbon, P. The Role of Oxytocin in Abnormal Brain Development: Effect on Glial Cells and Neuroinflammation. Cells 2022, 11, 3899.
- Mehdi, S.F.; Pusapati, S.; Khenhrani, R.R.; Farooqi, M.S.; Sarwar, S.; Alnasarat, A.; Mathur, N.; Metz, C.N.; LeRoith, D.; Tracey, K.J.; et al. Oxytocin and Related Peptide Hormones: Candidate Anti-Inflammatory Therapy in Early Stages of Sepsis. Front. Immunol. 2022, 13, 864007.
- Tang, Y.; Shi, Y.; Gao, Y.; Xu, X.; Han, T.; Li, J.; Liu, C. Oxytocin system alleviates intestinal inflammation by regulating macrophages polarization in experimental colitis. Clin. Sci. (Lond. Engl. 1979) 2019, 133, 1977–1992.
- Huang, S.; Zhu, B.; Cheon, I.S.; Goplen, N.P.; Jiang, L.; Zhang, R.; Peebles, R.S.; Mack, M.; Kaplan, M.H.; Limper, A.H.; et al. PPAR-γ in Macrophages Limits Pulmonary Inflammation and Promotes Host Recovery following Respiratory Viral Infection. J. Virol. 2019, 93, e00030-19.
- Eckertova, M.; Ondrejcakova, M.; Krskova, K.; Zorad, S.; Jezova, D. Subchronic treatment of rats with oxytocin results in improved adipocyte differentiation and increased gene expression of factors involved in adipogenesis. Br. J. Pharmacol. 2011, 162, 452–463.
- Marlin, B.J.; Froemke, R.C. Oxytocin modulation of neural circuits for social behavior. Dev. Neurobiol. 2017, 77, 169–189.
- Borland, J.M.; Grantham, K.N.; Aiani, L.M.; Frantz, K.J.; Albers, H.E. Role of oxytocin in the ventral tegmental area in social reinforcement. Psychoneuroendocrinology 2018, 95, 128–137.
- Walsh, J.J.; Christoffel, D.J.; Malenka, R.C. Neural circuits regulating prosocial behaviors. Neuropsychopharmacology 2022, 48, 79–89.
- Xiao, L.; Priest, M.F.; Nasenbeny, J.; Lu, T.; Kozorovitskiy, Y. Biased Oxytocinergic Modulation of Midbrain Dopamine Systems. Neuron 2017, 95, 368–384.e365.
- Song, Z.; Borland, J.M.; Larkin, T.E.; O’Malley, M.; Albers, H.E. Activation of oxytocin receptors, but not arginine-vasopressin V1a receptors, in the ventral tegmental area of male Syrian hamsters is essential for the reward-like properties of social interactions. Psychoneuroendocrinology 2016, 74, 164–172.
- Hurlemann, R.; Scheele, D. Dissecting the Role of Oxytocin in the Formation and Loss of Social Relationships. Biol. Psychiatry 2016, 79, 185–193.
- Barraza, J.A.; Zak, P.J. Empathy toward strangers triggers oxytocin release and subsequent generosity. Ann. N. Y. Acad. Sci. 2009, 1167, 182–189.
- Walum, H.; Young, L.J. The neural mechanisms and circuitry of the pair bond. Nat. Rev. Neurosci. 2018, 19, 643–654.
- Perkeybile, A.M.; Bales, K.L. Intergenerational transmission of sociality: The role of parents in shaping social behavior in monogamous and non-monogamous species. J. Exp. Biol. 2017, 220, 114–123.
- Insel, T.R.; Shapiro, L.E. Oxytocin receptor distribution reflects social organization in monogamous and polygamous voles. Proc. Natl. Acad. Sci. USA 1992, 89, 5981–5985.
- Bethlehem, R.A.I.; Lombardo, M.V.; Lai, M.C.; Auyeung, B.; Crockford, S.K.; Deakin, J.; Soubramanian, S.; Sule, A.; Kundu, P.; Voon, V.; et al. Intranasal oxytocin enhances intrinsic corticostriatal functional connectivity in women. Transl. Psychiatry 2017, 7, e1099.
- Freeman, S.M.; Young, L.J. Comparative Perspectives on Oxytocin and Vasopressin Receptor Research in Rodents and Primates: Translational Implications. J. Neuroendocrinol. 2016, 28.
- Yamagishi, A.; Lee, J.; Sato, N. Oxytocin in the anterior cingulate cortex is involved in helping behaviour. Behav. Brain Res. 2020, 393, 112790.
- Barchi-Ferreira, A.M.; Osorio, F.L. Associations between oxytocin and empathy in humans: A systematic literature review. Psychoneuroendocrinology 2021, 129, 105268.
- Akinrinade, I.; Kareklas, K.; Teles, M.C.; Reis, T.K.; Gliksberg, M.; Petri, G.; Levkowitz, G.; Oliveira, R.F. Evolutionarily conserved role of oxytocin in social fear contagion in zebrafish. Science 2023, 379, 1232–1237.
- Kosfeld, M.; Heinrichs, M.; Zak, P.J.; Fischbacher, U.; Fehr, E. Oxytocin increases trust in humans. Nature 2005, 435, 673–676.
- Menon, R.; Grund, T.; Zoicas, I.; Althammer, F.; Fiedler, D.; Biermeier, V.; Bosch, O.J.; Hiraoka, Y.; Nishimori, K.; Eliava, M.; et al. Oxytocin Signaling in the Lateral Septum Prevents Social Fear during Lactation. Curr. Biol. 2018, 28, 1066–1078.e6.
- Liberzon, I.; Young, E.A. Effects of stress and glucocorticoids on CNS oxytocin receptor binding. Psychoneuroendocrinology 1997, 22, 411–422.
- Matsushita, H.; Latt, H.M.; Koga, Y.; Nishiki, T.; Matsui, H. Oxytocin and Stress: Neural Mechanisms, Stress-Related Disorders, and Therapeutic Approaches. Neuroscience 2019, 417, 1–10.
- Owen, S.F.; Tuncdemir, S.N.; Bader, P.L.; Tirko, N.N.; Fishell, G.; Tsien, R.W. Oxytocin enhances hippocampal spike transmission by modulating fast-spiking interneurons. Nature 2013, 500, 458–462.
- Froemke, R.C.; Young, L.J. Oxytocin, Neural Plasticity, and Social Behavior. Annu. Rev. Neurosci. 2021, 44, 359–381.
- Eliava, M.; Melchior, M.; Knobloch-Bollmann, H.S.; Wahis, J.; da Silva Gouveia, M.; Tang, Y.; Ciobanu, A.C.; Triana Del Rio, R.; Roth, L.C.; Althammer, F.; et al. A New Population of Parvocellular Oxytocin Neurons Controlling Magnocellular Neuron Activity and Inflammatory Pain Processing. Neuron 2016, 89, 1291–1304.
- Marsh, N.; Marsh, A.A.; Lee, M.R.; Hurlemann, R. Oxytocin and the Neurobiology of Prosocial Behavior. Neuroscientist 2021, 27, 604–619.
- Ferretti, V.; Maltese, F.; Contarini, G.; Nigro, M.; Bonavia, A.; Huang, H.; Gigliucci, V.; Morelli, G.; Scheggia, D.; Managò, F.; et al. Oxytocin Signaling in the Central Amygdala Modulates Emotion Discrimination in Mice. Curr. Biol. 2019, 29, 1938–1953.e6.
- Hasan, M.T.; Althammer, F.; Silva da Gouveia, M.; Goyon, S.; Eliava, M.; Lefevre, A.; Kerspern, D.; Schimmer, J.; Raftogianni, A.; Wahis, J.; et al. A Fear Memory Engram and Its Plasticity in the Hypothalamic Oxytocin System. Neuron 2019, 103, 133–146.e8.
- Oliveira, V.E.M.; Lukas, M.; Wolf, H.N.; Durante, E.; Lorenz, A.; Mayer, A.L.; Bludau, A.; Bosch, O.J.; Grinevich, V.; Egger, V.; et al. Oxytocin and vasopressin within the ventral and dorsal lateral septum modulate aggression in female rats. Nat. Commun. 2021, 12, 2900.
- Haussler, H.U.; Jirikowski, G.F.; Caldwell, J.D. Sex differences among oxytocin-immunoreactive neuronal systems in the mouse hypothalamus. J. Chem. Neuroanat. 1990, 3, 271–276.
- Qiao, X.; Yan, Y.; Wu, R.; Tai, F.; Hao, P.; Cao, Y.; Wang, J. Sociality and oxytocin and vasopressin in the brain of male and female dominant and subordinate mandarin voles. J. Comp. Physiol. A 2014, 200, 149–159.
- Wang, Y.; Xu, L.; Pan, Y.; Wang, Z.; Zhang, Z. Species differences in the immunoreactive expression of oxytocin, vasopressin, tyrosine hydroxylase and estrogen receptor alpha in the brain of Mongolian gerbils (Meriones unguiculatus) and Chinese striped hamsters (Cricetulus barabensis). PLoS ONE 2013, 8, e65807.
- Xu, L.; Pan, Y.; Young, K.A.; Wang, Z.; Zhang, Z. Oxytocin and vasopressin immunoreactive staining in the brains of Brandt’s voles (Lasiopodomys brandtii) and greater long-tailed hamsters (Tscherskia triton). Neuroscience 2010, 169, 1235–1247.
- Nakajima, M.; Gorlich, A.; Heintz, N. Oxytocin modulates female sociosexual behavior through a specific class of prefrontal cortical interneurons. Cell 2014, 159, 295–305.
- Li, K.; Nakajima, M.; Ibanez-Tallon, I.; Heintz, N. A Cortical Circuit for Sexually Dimorphic Oxytocin-Dependent Anxiety Behaviors. Cell 2016, 167, 60–72.e11.
- Reh, R.K.; Dias, B.G.; Nelson, C.A., 3rd; Kaufer, D.; Werker, J.F.; Kolb, B.; Levine, J.D.; Hensch, T.K. Critical period regulation across multiple timescales. Proc. Natl. Acad. Sci. USA 2020, 117, 23242–23251.
- Molet, J.; Maras, P.M.; Avishai-Eliner, S.; Baram, T.Z. Naturalistic rodent models of chronic early-life stress. Dev. Psychobiol. 2014, 56, 1675–1688.
- Biagini, G.; Pich, E.M.; Carani, C.; Marrama, P.; Agnati, L.F. Postnatal maternal separation during the stress hyporesponsive period enhances the adrenocortical response to novelty in adult rats by affecting feedback regulation in the CA1 hippocampal field. Int. J. Dev. Neurosci. 1998, 16, 187–197.
- Huot, R.L.; Plotsky, P.M.; Lenox, R.H.; McNamara, R.K. Neonatal maternal separation reduces hippocampal mossy fiber density in adult Long Evans rats. Brain Res. 2002, 950, 52–63.
- Derks, N.A.; Krugers, H.J.; Hoogenraad, C.C.; Joels, M.; Sarabdjitsingh, R.A. Effects of Early Life Stress on Synaptic Plasticity in the Developing Hippocampus of Male and Female Rats. PLoS ONE 2016, 11, e0164551.
- Danoff, J.S.; Wroblewski, K.L.; Graves, A.J.; Quinn, G.C.; Perkeybile, A.M.; Kenkel, W.M.; Lillard, T.S.; Parikh, H.I.; Golino, H.F.; Gregory, S.G.; et al. Genetic, epigenetic, and environmental factors controlling oxytocin receptor gene expression. Clin. Epigenetics 2021, 13, 23.
- Francis, D.D.; Young, L.J.; Meaney, M.J.; Insel, T.R. Naturally occurring differences in maternal care are associated with the expression of oxytocin and vasopressin (V1a) receptors: Gender differences. J. Neuroendocrinol. 2002, 14, 349–353.
- Loken, L.S.; Wessberg, J.; Morrison, I.; McGlone, F.; Olausson, H. Coding of pleasant touch by unmyelinated afferents in humans. Nat. Neurosci. 2009, 12, 547–548.
- Kojima, S.; Stewart, R.A.; Demas, G.E.; Alberts, J.R. Maternal contact differentially modulates central and peripheral oxytocin in rat pups during a brief regime of mother-pup interaction that induces a filial huddling preference. J. Neuroendocrinol. 2012, 24, 831–840.
- Hertenstein, M.J.; Verkamp, J.M.; Kerestes, A.M.; Holmes, R.M. The communicative functions of touch in humans, nonhuman primates, and rats: A review and synthesis of the empirical research. Genet. Soc. Gen. Psychol. Monogr. 2006, 132, 5–94.
- Havranek, T.; Lestanova, Z.; Mravec, B.; Strbak, V.; Bakos, J.; Bacova, Z. Oxytocin Modulates Expression of Neuron and Glial Markers in the Rat Hippocampus. Folia Biol. 2017, 63, 91–97.
- Block, M.L.; Zecca, L.; Hong, J.S. Microglia-mediated neurotoxicity: Uncovering the molecular mechanisms. Nat. Rev. Neurosci. 2007, 8, 57–69.
- Bazargani, N.; Attwell, D. Astrocyte calcium signaling: The third wave. Nat. Neurosci. 2016, 19, 182–189.
- Wahis, J.; Baudon, A.; Althammer, F.; Kerspern, D.; Goyon, S.; Hagiwara, D.; Lefevre, A.; Barteczko, L.; Boury-Jamot, B.; Bellanger, B.; et al. Astrocytes mediate the effect of oxytocin in the central amygdala on neuronal activity and affective states in rodents. Nat. Neurosci. 2021, 24, 529–541.
- Roque, A.; Ochoa-Zarzosa, A.; Torner, L. Maternal separation activates microglial cells and induces an inflammatory response in the hippocampus of male rat pups, independently of hypothalamic and peripheral cytokine levels. Brain Behav. Immun. 2016, 55, 39–48.
- Banqueri, M.; Mendez, M.; Gomez-Lazaro, E.; Arias, J.L. Early life stress by repeated maternal separation induces long-term neuroinflammatory response in glial cells of male rats. Stress 2019, 22, 563–570.
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487.
- Allen, N.J.; Lyons, D.A. Glia as architects of central nervous system formation and function. Science 2018, 362, 181–185.
- Perea, G.; Gomez, R.; Mederos, S.; Covelo, A.; Ballesteros, J.J.; Schlosser, L.; Hernandez-Vivanco, A.; Martin-Fernandez, M.; Quintana, R.; Rayan, A.; et al. Activity-dependent switch of GABAergic inhibition into glutamatergic excitation in astrocyte-neuron networks. eLife 2016, 5, e20362.
- Ta, T.T.; Dikmen, H.O.; Schilling, S.; Chausse, B.; Lewen, A.; Hollnagel, J.O.; Kann, O. Priming of microglia with IFN-gamma slows neuronal gamma oscillations in situ. Proc. Natl. Acad. Sci. USA 2019, 116, 4637–4642.
- Hasan, M.; Kanna, M.S.; Jun, W.; Ramkrishnan, A.S.; Iqbal, Z.; Lee, Y.; Li, Y. Schema-like learning and memory consolidation acting through myelination. FASEB J. 2019, 33, 11758–11775.
- Baskerville, T.A.; Douglas, A.J. Dopamine and oxytocin interactions underlying behaviors: Potential contributions to behavioral disorders. CNS Neurosci. Ther. 2010, 16, e92–e123.
- Yamamoto, Y.; Liang, M.; Munesue, S.; Deguchi, K.; Harashima, A.; Furuhara, K.; Yuhi, T.; Zhong, J.; Akther, S.; Goto, H.; et al. Vascular RAGE transports oxytocin into the brain to elicit its maternal bonding behaviour in mice. Commun. Biol. 2019, 2, 76.
- Zhao, F.; Zhang, H.; Wang, P.; Cui, W.; Xu, K.; Chen, D.; Hu, M.; Li, Z.; Geng, X.; Wei, S. Oxytocin and serotonin in the modulation of neural function: Neurobiological underpinnings of autism-related behavior. Front. Neurosci. 2022, 16, 919890.
- Zhang, R.; Jia, M.X.; Zhang, J.S.; Xu, X.J.; Shou, X.J.; Zhang, X.T.; Li, L.; Li, N.; Han, S.P.; Han, J.S. Transcutaneous electrical acupoint stimulation in children with autism and its impact on plasma levels of arginine-vasopressin and oxytocin: A prospective single-blinded controlled study. Res. Dev. Disabil. 2012, 33, 1136–1146.
- Huber, D.; Veinante, P.; Stoop, R. Vasopressin and oxytocin excite distinct neuronal populations in the central amygdala. Science 2005, 308, 245–248.
- Parker, K.J.; Oztan, O.; Libove, R.A.; Sumiyoshi, R.D.; Jackson, L.P.; Karhson, D.S.; Summers, J.E.; Hinman, K.E.; Motonaga, K.S.; Phillips, J.M.; et al. Intranasal oxytocin treatment for social deficits and biomarkers of response in children with autism. Proc. Natl. Acad. Sci. USA 2017, 114, 8119–8124.
- Guastella, A.J.; Einfeld, S.L.; Gray, K.M.; Rinehart, N.J.; Tonge, B.J.; Lambert, T.J.; Hickie, I.B. Intranasal oxytocin improves emotion recognition for youth with autism spectrum disorders. Biol. Psychiatry 2010, 67, 692–694.
- El-Ansary, A.; Al-Ayadhi, L. Neuroinflammation in autism spectrum disorders. J. Neuroinflammation 2012, 9, 265.
- Lee, J.H.; Espinera, A.R.; Chen, D.; Choi, K.E.; Caslin, A.Y.; Won, S.; Pecoraro, V.; Xu, G.Y.; Wei, L.; Yu, S.P. Neonatal inflammatory pain and systemic inflammatory responses as possible environmental factors in the development of autism spectrum disorder of juvenile rats. J. Neuroinflammation 2016, 13, 109.
- Iannucci, A.; Caneparo, V.; Raviola, S.; Debernardi, I.; Colangelo, D.; Miggiano, R.; Griffante, G.; Landolfo, S.; Gariglio, M.; De Andrea, M. Toll-like receptor 4-mediated inflammation triggered by extracellular IFI16 is enhanced by lipopolysaccharide binding. PLoS Pathog. 2020, 16, e1008811.
- Alabdali, A.; Al-Ayadhi, L.; El-Ansary, A. Association of social and cognitive impairment and biomarkers in autism spectrum disorders. J. Neuroinflammation 2014, 11, 4.
- Geschwind, D.H.; Levitt, P. Autism spectrum disorders: Developmental disconnection syndromes. Curr. Opin. Neurobiol. 2007, 17, 103–111.
- Long, Z.; Duan, X.; Mantini, D.; Chen, H. Alteration of functional connectivity in autism spectrum disorder: Effect of age and anatomical distance. Sci. Rep. 2016, 6, 26527.
- Goh, K.K.; Chen, C.Y.; Wu, T.H.; Chen, C.H.; Lu, M.L. Crosstalk between Schizophrenia and Metabolic Syndrome: The Role of Oxytocinergic Dysfunction. Int. J. Mol. Sci. 2022, 23, 7092.
- Winship, I.R.; Dursun, S.M.; Baker, G.B.; Balista, P.A.; Kandratavicius, L.; Maia-de-Oliveira, J.P.; Hallak, J.; Howland, J.G. An Overview of Animal Models Related to Schizophrenia. Can. J. Psychiatry 2019, 64, 5–17.
- Montag, C.; Brockmann, E.M.; Bayerl, M.; Rujescu, D.; Müller, D.J.; Gallinat, J. Oxytocin and oxytocin receptor gene polymorphisms and risk for schizophrenia: A case-control study. World J. Biol. Psychiatry Off. J. World Fed. Soc. Biol. Psychiatry 2013, 14, 500–508.
- Goh, K.K.; Chen, C.H.; Lane, H.Y. Oxytocin in Schizophrenia: Pathophysiology and Implications for Future Treatment. Int. J. Mol. Sci. 2021, 22, 2146.
- Young, J.W.; Geyer, M.A. Developing treatments for cognitive deficits in schizophrenia: The challenge of translation. J. Psychopharmacol. 2015, 29, 178–196.
- Millan, M.J.; Bales, K.L. Towards improved animal models for evaluating social cognition and its disruption in schizophrenia: The CNTRICS initiative. Neurosci. Biobehav. Rev. 2013, 37, 2166–2180.
- Ferguson, J.N.; Young, L.J.; Hearn, E.F.; Matzuk, M.M.; Insel, T.R.; Winslow, J.T. Social amnesia in mice lacking the oxytocin gene. Nat. Genet. 2000, 25, 284–288.
- Ferguson, J.N.; Aldag, J.M.; Insel, T.R.; Young, L.J. Oxytocin in the medial amygdala is essential for social recognition in the mouse. J. Neurosci. Off. J. Soc. Neurosci. 2001, 21, 8278–8285.
- Crawley, J.N.; Chen, T.; Puri, A.; Washburn, R.; Sullivan, T.L.; Hill, J.M.; Young, N.B.; Nadler, J.J.; Moy, S.S.; Young, L.J.; et al. Social approach behaviors in oxytocin knockout mice: Comparison of two independent lines tested in different laboratory environments. Neuropeptides 2007, 41, 145–163.
- Popik, P.; van Ree, J.M. Oxytocin but not vasopressin facilitates social recognition following injection into the medial preoptic area of the rat brain. Eur. Neuropsychopharmacol. 1991, 1, 555–560.
- Kirsch, P.; Esslinger, C.; Chen, Q.; Mier, D.; Lis, S.; Siddhanti, S.; Gruppe, H.; Mattay, V.S.; Gallhofer, B.; Meyer-Lindenberg, A. Oxytocin modulates neural circuitry for social cognition and fear in humans. J. Neurosci. Off. J. Soc. Neurosci. 2005, 25, 11489–11493.