Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Malay Kumar Adak and Version 2 by Lindsay Dong.
Submergence in rice fields creating inundation stress and realizing anoxia or hypoxia is a problem in agriculture. Seeds under this oxygen deficit are faced with fermentative respiration, where the end product would be poisoning the tissue viability. This is more aggravated in direct seeded rice cultivation with the accumulation of lactate as a poison.
- tolerance
- genetic variabilities
- rice
1. Introduction
Rice, a semi-aquatic plant, requires a significant amount of water and tissue hydration throughout its developmental stages. The germination of rice seeds requires more soil moisture than other crops, up to the level of full submergence. Seeds of rice require hypoxia or anoxia under inundation conditions for germination [1][3]. The germination of rice seeds under hypoxic conditions induces one type of carbohydrate metabolism for starch and mobilization via enzymatic induction. The ability of rice seeds to tolerate hypoxia or anoxia also offers direct seeding in agronomic practices [2][4]. Direct seeding is more economic than seedling transplantation, where the availability of complete submergence is frequent. Herein, the chances of weed intolerance from floods are highly possible and obstructed by infestation from pests [3][5]. No matter the species, rice seeds are sensitive to submergence for germination and few cultivars have a better germination rate or seedling tolerance to hypoxia. Tolerant rice seeds exhibit better coleoptile elongation, the induction of carbohydrates, fermentative pathways, and reduced oxidative damages indicating stress tolerance [4][6].
2. Physiological Relevance of Seeds’ Response to Anoxia
The germination of seeds, establishment of seedlings, and development into plants are subjected to flooding stress. Therefore, seed development under anoxic conditions would be sensitive to anoxia or hypoxia [5][12]. When flooding occurs during seed germination (under complete submergence), a distinct shift into anaerobic metabolism is observed. Rice plants that are semi-aquatic in nature can experience withstanding anaerobic respiration mostly for maintaining respiration, meaning seed tissues are viable under the water. Coleoptiles of sprouting seeds are exposed to anoxia, and total carbohydrate metabolism is shifted toward anaerobic respiration, mostly via alcoholic fermentation. A number of genes like alcohol dehydrogenase (ADH) and pyruvate decarboxylase (PDC) are the most important in inducing fermentation metabolism [6][13]. Even rice seeds, which are dormant in nature, also remain non-viable under water. The dormancy of rice seeds and their further progress under water are regulated by an endogenous ratio of abscisic acid (ABA) to gibberellic acid (GA) [7][14]. The major signaling gene(s) for ABA metabolism in rice show their expression with varied modalities through the anaerobic phases. The genes that are predominantly involved include OsABI5, OsDSG1, OsNCED3, OsVP1, OsPDS, and OsZDS for ABA metabolism [8][15]. These are actual key genes governing seed dormancy and germination, and are mostly required for the signaling and metabolism of ABA [9][16].
3. Hormonal Interplay between Seed Dormancy and Germination in Rice under Water
It is evident that seed dormancy and germination play a key role in plant initial survival against submergence. Growth regulators like ABA and GA and their ratio compete with each other, favoring either dormancy or germination depending on the physiological needs [10][20]. ABA suppresses the physiologically assisted genes for germination whereas GAs can erase the effects. Embryo activity at the cellular level overcoming mechanical distances via tests is important. All of the activity of developing embryos is regulated by GA-assisted genes. The latter, with their increasing rates of transcription and translation, may support ABA-suppressed activities of quiescence strategies [11][21]. In seeds, a precise concentration of GA is maintained by a balance between biosynthesis and its turnover via oxidative degradation. Regulation for de novo biosynthesis is also important in the context of GA activity on seedling development. This becomes more complicated given that under water a significant level of reactive oxygen species (ROS) and Ca2+-dependent signaling are prerequisites. This may integrate downstream pathways for anoxia tolerance and the germination of seeds [12][22]. This is explained more by the fact that ROS can upregulate a number of ABA-metabolizing gene (particularly ABAI3 and AP2) transcription factors (TFs). Auxins (Au) are also involved to allow the binding of a few TFs, and thereby they induce downstream genes related to ABA metabolism. Au, like GA involvement, in this aspect may concern other TFs like AP2 and DELLA and other proteins. These often cause GA oxidation pathways involving genes like GA20ox, GA3ox, and PIL5 TF [13][23]. After the ripening of embryos in seeds, a number of genes are involved, like RGA, RGL2, and GAI, from different insensitive mutants to GA. For the up- or downregulation of ABA-inducing genes, growth substances are involved including jasmonic acid (JA), brassinosteroid (BR), ethylene (ET), salicylic acid (SA), etc. [14][24]. ABA, through its auxin inductive cascade, can induce a number of genes via some auxin response factors (ARF10/16). These factors are involved to allow the binding of other TFs. Likewise, ABA can induce MYB96 factor, which otherwise induces TF ABI4. The latter is responsible for the suppression of CYP707A1/2 factor activity, which is likely to bind the α-amylase promoter and promotes seed germination [15][25]. ABA can induce other TFs (ABI5, BIN2, PKS5, etc.) which also inhibit seed germination. All of these are related to the precise concentration of ROS, which collectively are key balancing factors for gene regulation. ABA, under anaerobic conditions, is more active with regard to its regulation of catabolism as well. On the seeds and vegetative parts of plants, a number of ABA receptor components are grouped and these regulate protein phosphatase 2C (PP2C) when ABA is present. PP2C is inactive under aerobic conditions but is activated with sucrose non-fermenting 1 (SNF1)-related kinase 2. The latter allows for specific response element binding TFs [16][26]. Increased dormancy is also related to ABA, where a particular factor, DELAY OF GERMINATION 1 (DOG1), becomes the master regulator of the primary dormancy activator. This is in turn regulated by a specific PP2C, activated by ABA [17][27]. DOG1 modifies ABA signaling where PP2C acts on seeds, particularly regarding those depending on ABA hypersensitive germination. Even after grain development, OsDOG1-like gene expression is also involved for immature seed development. This is co-dominantly expressed with an over expression of OsNCED2 and OsABA8’OH3. Seed germination under water is independent of primary dormancy, where susceptible varieties can germinate but fail through downstream development. Not only embryos under submergence stress but also heat-stressed embryos are dependent on DNA methylation. The DNA methylation of ABA-catabolizing genes and the α-amylase promoter are also subjected to anaerobic stress [18][28]. Another two genes, ABA deficient 4 (ABA4) and Neoxanthin deficient 1 (NXD1), are required for ABA biosynthesis and its regulation [19][29]. Therefore, more studies on ABA-GA cross talk in the germination phenomenon with molecular insights into submergence or anoxic germination are required (Figure 1).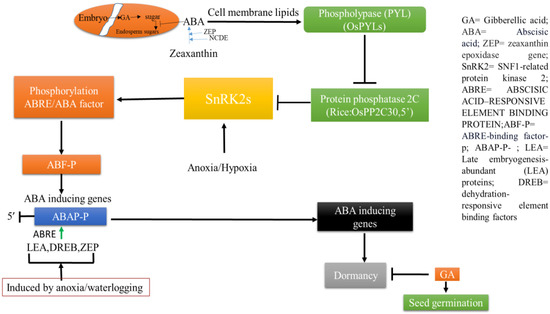
Figure 1. Hormonal interplay between seed dormancy and germination under inundation stress.
4. Development of Rice Seedlings Facing Anoxic Conditions
Soil flooding may be the most vulnerable abiotic constraint in rice cultivation after seed germination. Seedling growth becomes important with the duration and depth of water stagnation. Seeds of taller plants almost fail to germinate under anaerobic conditions whereas rice germinates in oxygen-depleted soil under partial or full submergence [20]. Rice seeds starchy in grain material could tolerate anaerobiosis, where fermentative catabolism is a prime event. Thereby, seeds can maintain a high rate of carbohydrate turnover under oxygen deficiency. A decline in oxygen partial pressure causes negative effects in other cereals like oat and barley, where root emergence is normal [21]. In contrast, even though root growth is suppressed, shoot growth increases where there is oxygen demand under submergence in the case of rice. Even depleted oxygen can increase the final length of coleoptiles that exceeds that of aerobically grown seeds. Under this condition, coleoptiles may be sustained, but the root and primarily the leaves do not grow successfully.
Hormonal interplay between seed dormancy and germination under inundation stress.
4. Development of Rice Seedlings Facing Anoxic Conditions
Soil flooding may be the most vulnerable abiotic constraint in rice cultivation after seed germination. Seedling growth becomes important with the duration and depth of water stagnation. Seeds of taller plants almost fail to germinate under anaerobic conditions whereas rice germinates in oxygen-depleted soil under partial or full submergence [30]. Rice seeds starchy in grain material could tolerate anaerobiosis, where fermentative catabolism is a prime event. Thereby, seeds can maintain a high rate of carbohydrate turnover under oxygen deficiency. A decline in oxygen partial pressure causes negative effects in other cereals like oat and barley, where root emergence is normal [31]. In contrast, even though root growth is suppressed, shoot growth increases where there is oxygen demand under submergence in the case of rice. Even depleted oxygen can increase the final length of coleoptiles that exceeds that of aerobically grown seeds. Under this condition, coleoptiles may be sustained, but the root and primarily the leaves do not grow successfully.
5. Fermentative Mechanism: Pathways for Anaerobic Seeds’ Germination in Rice
Under limited oxygen concentration, mitochondrial respiration is inversely proportional to glycosylate metabolism, which produces NADH. Following entry into mitochondria, NADH undergoes re-oxidation at the start of fermentative pathways. The lactate and toxic substrate ethanol produced with a minimum gain of ATP is a bottleneck for germination under water [22][36]. Lactate, a substrate of lactate dehydrogenase (LDH), is typically toxic to the aleurone membrane in inducing amylase activities by GA, whereas PCD produces ethanol along with the activity of ADH. Seeds may be tolerant to anoxia by reverting the central glycolytic pathways. Ethanol may either be oxidized to acetyl dehydrogenase or diffused out of the seed coat, accumulating within spaces of grains and glumes [23][37]. This also induces dormancy in terms of ecological consideration for submergence tolerance. The minimum amount of ATP (2 moles) over the normal tricarboxylic acid cycle (32 moles) is a limitation for anaerobic respiration [24][38]. Therefore, a specific set of protein expression and their corresponding regulation are selection criteria for the better germination of seeds in the anaerobic mode. Distinct proteins like sucrose-phosphate synthase, PDC, and ADH are the most important in accessing breeding programs where submergence induces anoxia [25][39]. Carbohydrate metabolism in rice seeds under prolonged anoxic conditions creates the most effective screening index. There are specific modalities of the regulation of genes on the anaerobic response elements upstream of the promoters, characterizing the coding of anaerobic proteins [26][40]. These, in turn, become the factors to control the paths of fermentation for specific genes. From the sequence alignment, cis-elements also bear the homology for anoxia-inducible genes in plants, similar to bacteria. Rice plants tolerant to anoxia can accommodate ATPs; however, for a long time, this has occurred via fermentative catabolism, as long as the hypoxia is maintained without affecting the embryo tissues.