Caffeic acid phenethyl ester (CAPE) is a phenylpropanoid naturally found in propolis that shows important biological activities, including neuroprotective activity by modulating the Nrf2 and NF-κB pathways, promoting antioxidant enzyme expression and inhibition of proinflammatory cytokine expression. Its simple chemical structure has inspired the synthesis of many derivatives, with aliphatic and/or aromatic moieties, some of which have improved the biological properties. Moreover, new drug delivery systems increase the bioavailability of these compounds in vivo, allowing its transcytosis through the blood-brain barrier, thus protecting brain cells from the increased inflammatory status associated to neurodegenerative and psychiatric disorders.
- caffeic acid phenethyl ester
- CAPE
1. Introduction
2. Natural Sources of Caffeic Acid Phenethyl Ester
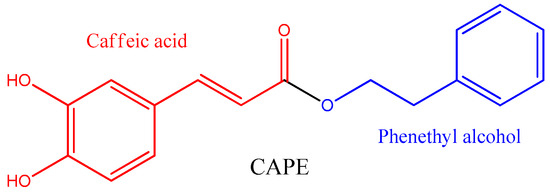
| Source | Concentration of CAPE (mg g−1) | Reference |
|---|---|---|
| Populus buds | 1.611 ± 0.272 | [25] |
| Propolis from Mexico (Ures) | 11.4 | [26] |
| Propolis from Spain (Viveros) | 10.1 | [27] |
| Propolis from Spain (Valencia) | 10.4 | [27] |
| Propolis from Chile | 5.8 | [27] |
| Propolis from China | 4.9 | [27] |
3. Biosynthesis of Caffeic Acid Phenethyl Ester
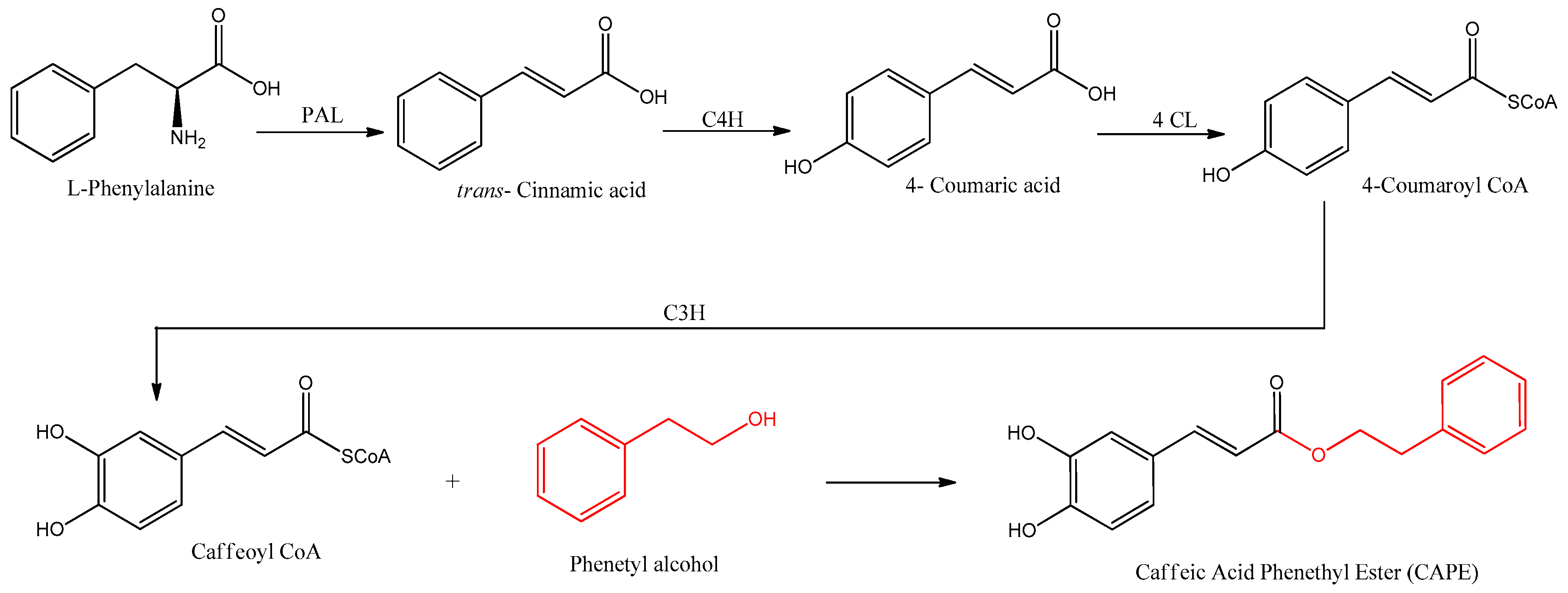
References
- Dubois, B.; Villain, N.; Frisoni, G.B.; Rabinovici, G.D.; Sabbagh, M.; Cappa, S.; Bejanin, A.; Bombois, S.; Epelbaum, S.; Teichmann, M.; et al. Clinical Diagnosis of Alzheimer’s Disease: Recommendations of the International Working Group. Lancet Neurol. 2021, 20, 484–496.
- Fukuhara, S.; Tanigaki, R.; Kimura, K.; Kataoka, T. International Immunopharmacology Kujigamberol Interferes with Pro-in Fl Ammatory Cytokine-Induced Expression of and N-Glycan Modi Fi Cations to Cell Adhesion Molecules at Di Ff Erent Stages in Human Umbilical Vein Endothelial Cells. Int. Immunopharmacol. 2018, 62, 313–325.
- Heppner, F.L.; Ransohoff, R.M.; Becher, B. Immune Attack: The Role of Inflammation in Alzheimer Disease. Nat. Rev. Neurosci. 2015, 16, 358–372.
- Ebadi, M.; Srinivasan, S.K.; Baxi, M.D. Oxidative Stress and Antioxidant Therapy in Parkinson’s Disease. Prog. Neurobiol. 1996, 48, 1–19.
- Markesbery, W.R.; Carney, J.M. Oxidative Alterations in Alzheimer’s Disease. Brain Pathol. 1999, 9, 133–146.
- Barber, S.C.; Shaw, P.J. Oxidative Stress in ALS: Key Role in Motor Neuron Injury and Therapeutic Target. Free Radic. Biol. Med. 2010, 48, 629–641.
- Leiva, A.M.; Martínez-Sanguinetti, M.A.; Troncoso-Pantoja, C.; Nazar, G.; Petermann-Rocha, F.; Celis-Morales, C. Parkinson’s Disease in Chile: Highest Prevalence in Latin America. Rev. Med. Chil. 2019, 147, 535–536.
- Kobayashi, A.; Kang, M.-I.; Okawa, H.; Ohtsuji, M.; Zenke, Y.; Chiba, T.; Igarashi, K.; Yamamoto, M. Oxidative Stress Sensor Keap1 Functions as an Adaptor for Cul3-Based E3 Ligase to Regulate Proteasomal Degradation of Nrf2. Mol. Cell. Biol. 2004, 24, 7130–7139.
- Cullinan, S.B.; Gordan, J.D.; Jin, J.; Harper, J.W.; Diehl, J.A. The Keap1-BTB Protein Is an Adaptor That Bridges Nrf2 to a Cul3-Based E3 Ligase: Oxidative Stress Sensing by a Cul3-Keap1 Ligase. Mol. Cell. Biol. 2004, 24, 8477–8486.
- Sykiotis, G.P.; Bohmann, D. Stress-Activated Cap’n’collar Transcription Factors in Aging and Human Disease. Sci. Signal. 2010, 3, re3.
- Almowallad, S.; Alqahtani, L.S.; Mobashir, M. NF-ΚB in Signaling Patterns and Its Temporal Dynamics Encode/Decode Human Diseases. Life 2022, 12, 2012.
- Borba, R.S.; Wilson, M.B.; Spivak, M. Hidden Benefits of Honeybee Propolis in Hives. In Beekeeping—From Science to Practice; Springer: Cham, Switzerland, 2017; pp. 17–38.
- Przybyłek, I.; Karpiński, T.M. Antibacterial Properties of Propolis. Molecules 2019, 24, 2047.
- Drescher, N.; Klein, A.-M.; Neumann, P.; Yañez, O.; Leonhardt, S.D. Inside Honeybee Hives: Impact of Natural Propolis on the Ectoparasitic Mite Varroa Destructor and Viruses. Insects 2017, 8, 15.
- Machado, B.A.S.; Silva, R.P.D.; Barreto, G.d.A.; Costa, S.S.; Silva, D.F.d.; Brandão, H.N.; Rocha, J.L.C.d.; Dellagostin, O.A.; Henriques, J.A.P.; Umsza-Guez, M.A.; et al. Chemical Composition and Biological Activity of Extracts Obtained by Supercritical Extraction and Ethanolic Extraction of Brown, Green and Red Propolis Derived from Different Geographic Regions in Brazil. PLoS ONE 2016, 11, e0145954.
- Freires, I.A.; Queiroz, V.C.P.P.; Furletti, V.F.; Ikegaki, M.; de Alencar, S.M.; Duarte, M.C.T.; Rosalen, P.L. Chemical Composition and Antifungal Potential of Brazilian Propolis against Candida spp. J. Mycol. Med. 2016, 26, 122–132.
- Pazin, W.M.; Monaco, L.d.M.; Egea Soares, A.E.; Miguel, F.G.; Berretta, A.A.; Ito, A.S. Antioxidant Activities of Three Stingless Bee Propolis and Green Propolis Types. J. Apic. Res. 2017, 56, 40–49.
- Tiveron, A.P.; Rosalen, P.L.; Franchin, M.; Lacerda, R.C.C.; Bueno-Silva, B.; Benso, B.; Denny, C.; Ikegaki, M.; de Alencar, S.M. Chemical Characterization and Antioxidant, Antimicrobial, and Anti-Inflammatory Activities of South Brazilian Organic Propolis. PLoS ONE 2016, 11, e0165588.
- Metzner, J.; Bekemeier, H.; Paintz, M.; Schneidewind, E. On the antimicrobial activity of propolis and propolis constituents (author’s transl). Pharmazie 1979, 34, 97–102.
- Romero, F.; Palacios, J.; Jofré, I.; Paz, C.; Nwokocha, C.R.; Paredes, A.; Cifuentes, F. Aristoteline, an Indole-Alkaloid, Induces Relaxation by Activating Potassium Channels and Blocking Calcium Channels in Isolated Rat Aorta. Molecules 2019, 24, 2748.
- Kurek-Górecka, A.; Rzepecka-Stojko, A.; Górecki, M.; Stojko, J.; Sosada, M.; Swierczek-Zieba, G. Structure and Antioxidant Activity of Polyphenols Derived from Propolis. Molecules 2013, 19, 78–101.
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-Antioxidant Activity Relationships of Flavonoids and Phenolic Acids. Free Radic. Biol. Med. 1996, 20, 933–956.
- Ozdal, T.; Ceylan, F.D.; Eroglu, N.; Kaplan, M.; Olgun, E.O.; Capanoglu, E. Investigation of Antioxidant Capacity, Bioaccessibility and LC-MS/MS Phenolic Profile of Turkish Propolis. Food Res. Int. 2019, 122, 528–536.
- Gómez-Caravaca, A.M.; Gómez-Romero, M.; Arráez-Román, D.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Advances in the Analysis of Phenolic Compounds in Products Derived from Bees. J. Pharm. Biomed. Anal. 2006, 41, 1220–1234.
- Zhu, Y.; Shen, T.; Lin, Y.; Chen, B.; Ruan, Y.; Cao, Y.; Qiao, Y.; Man, Y.; Wang, S.; Li, J. Astragalus Polysaccharides Suppress ICAM-1 and VCAM-1 Expression in TNF-α-Treated Human Vascular Endothelial Cells by Blocking NF-ΚB Activation. Nat. Publ. Gr. 2013, 34, 1036–1042.
- Hernandez, J.; Goycoolea, F.M.; Quintero, J.; Acosta, A.; Castañeda, M.; Dominguez, Z.; Robles, R.; Vazquez-Moreno, L.; Velazquez, E.F.; Astiazaran, H.; et al. Sonoran Propolis: Chemical Composition and Antiproliferative Activity on Cancer Cell Lines. Planta Med. 2007, 73, 1469–1474.
- Borrás, M.J. Universitat Politècnica De València Perfil Fenólico De Propóleos De Diferentes Orígenes Geográficos. Master’s Thesis, Universitat Politècnica de València, València, Spain, 2018.
- Li, N.; Ng, T.B.; Wong, J.H.; Qiao, J.X.; Zhang, Y.N.; Zhou, R.; Chen, R.R.; Liu, F. Separation and Purification of the Antioxidant Compounds, Caffeic Acid Phenethyl Ester and Caffeic Acid from Mushrooms by Molecularly Imprinted Polymer. Food Chem. 2013, 139, 1161–1167.
- Chen, Y.J.; Shiao, M.S.; Hsu, M.L.; Tsai, T.H.; Wang, S.Y. Effect of Caffeic Acid Phenethyl Ester, an Antioxidant from Propolis, on Inducing Apoptosis in Human Leukemic HL-60 Cells. J. Agric. Food Chem. 2001, 49, 5615–5619.
- Watabe, M.; Hishikawa, K.; Takayanagi, A.; Shimizu, N.; Nakaki, T. Caffeic Acid Phenethyl Ester Induces Apoptosis by Inhibition of NFkappaB and Activation of Fas in Human Breast Cancer MCF-7 Cells. J. Biol. Chem. 2004, 279, 6017–6026.
- Xiang, D.; Wang, D.; He, Y.; Xie, J.; Zhong, Z.; Li, Z.; Xie, J. Caffeic Acid Phenethyl Ester Induces Growth Arrest and Apoptosis of Colon Cancer Cells via the Beta-Catenin/T-Cell Factor Signaling. Anticancer. Drugs 2006, 17, 753–762.
- Chen, J.H.; Shao, Y.; Huang, M.T.; Chin, C.K.; Ho, C.T. Inhibitory Effect of Caffeic Acid Phenethyl Ester on Human Leukemia HL-60 Cells. Cancer Lett. 1996, 108, 211–214.
- Murtaza, G.; Sajjad, A.; Mehmood, Z.; Shah, S.H.; Siddiqi, A.R. Possible Molecular Targets for Therapeutic Applications of Caffeic Acid Phenethyl Ester in Inflammation and Cancer. J. Food Drug Anal. 2015, 23, 11–18.
- Jiang, M.; Wu, N.; Xu, B.; Chu, Y.; Li, X.; Su, S.; Chen, D.; Li, W.; Shi, Y.; Gao, X.; et al. Fatty Acid-Induced CD36 Expression via O-GlcNAcylation Drives Gastric Cancer Metastasis. Theranostics 2019, 9, 5359–5373.
- Omar, M.H.; Mullen, W.; Stalmach, A.; Auger, C.; Rouanet, J.-M.; Teissedre, P.-L.; Caldwell, S.T.; Hartley, R.C.; Crozier, A. Absorption, Disposition, Metabolism, and Excretion of Caffeic Acid in Rats. J. Agric. Food Chem. 2012, 60, 5205–5214.
- Celli, N.; Dragani, L.K.; Murzilli, S.; Pagliani, T.; Poggi, A. In Vitro and in Vivo Stability of Caffeic Acid Phenethyl Ester, a Bioactive Compound of Propolis. J. Agric. Food Chem. 2007, 55, 3398–3407.
- de Oliveira, D.M.; Sampaio, G.R.; Pinto, C.B.; Catharino, R.R.; Bastos, D.H.M. Bioavailability of Chlorogenic Acids in Rats after Acute Ingestion of Maté Tea (Ilex Paraguariensis) or 5-Caffeoylquinic Acid. Eur. J. Nutr. 2017, 56, 2541–2556.