Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Jessie Wu and Version 1 by Samah Hamza Osman Zarroug.
Antimicrobial resistance (AMR) due to the prevalence of multidrug-resistant (MDR) pathogens is rapidly increasing worldwide, and the identification of new antimicrobial agents with innovative mechanisms of action is urgently required. Medicinal plants that have been utilised for centuries with minor side effects may hold great promise as sources of effective antimicrobial products. The free-living nematode Caenorhabditis elegans (C. elegans) is an excellent live infection model for the discovery and development of new antimicrobial compounds. However, while C. elegans has widely been utilised to explore the effectiveness and toxicity of synthetic antibiotics, it has not been used to a comparable extent for the analysis of natural products.
- natural products
- antimicrobial activity
- multidrug-resistant pathogens
1. Advantages of C.aenorhabditis elegans as a Model for the Screening of Novel Compounds
To address these issues, in the last decade, antimicrobial drug discovery technology has moved towards the use of whole animal models, such as zebrafish and C. elegans, for the screening of new compounds and the identification of their mechanisms of action.
Caenorhabditis elegans (C. elegans), a bacteria-eating organism, can be infected and killed by a variety of human pathogens, including Gram-negative bacteria such as Pseudomonas aeruginosa and Salmonella enterica; Gram-positive bacteria such as Staphylococcus aureus, Streptococcus pneumoniae, and Enterococcus faecalis; and fungi such as those of the Candida species [16,17,24,25][1][2][3][4].
To screen for novel antimicrobial compounds, the C. elegans natural food source, E. coli OP50, is replaced by a pathogenic bacterium, e.g., Pseudomonas aeruginosa. Survival assays are performed either on agar or in liquid media [16,25,26,27][1][4][5][6]. Initially, worms are incubated with pathogenic organisms to establish infection before transferring them into assay plates (12-, 24-, 96-, or 384-well plates) that are subsequently added with different concentrations of novel compounds. The number of live and dead worms can be counted visually under the microscope or, in automated assays, by using a cell-permeable dye, identifying dead worms, and image analysis software [17,28][2][7] (Figure 1). Notably, automated assays with 10–20 worms per well in 96- or 384-well plates allow for the screening of large numbers of compounds [29][8].
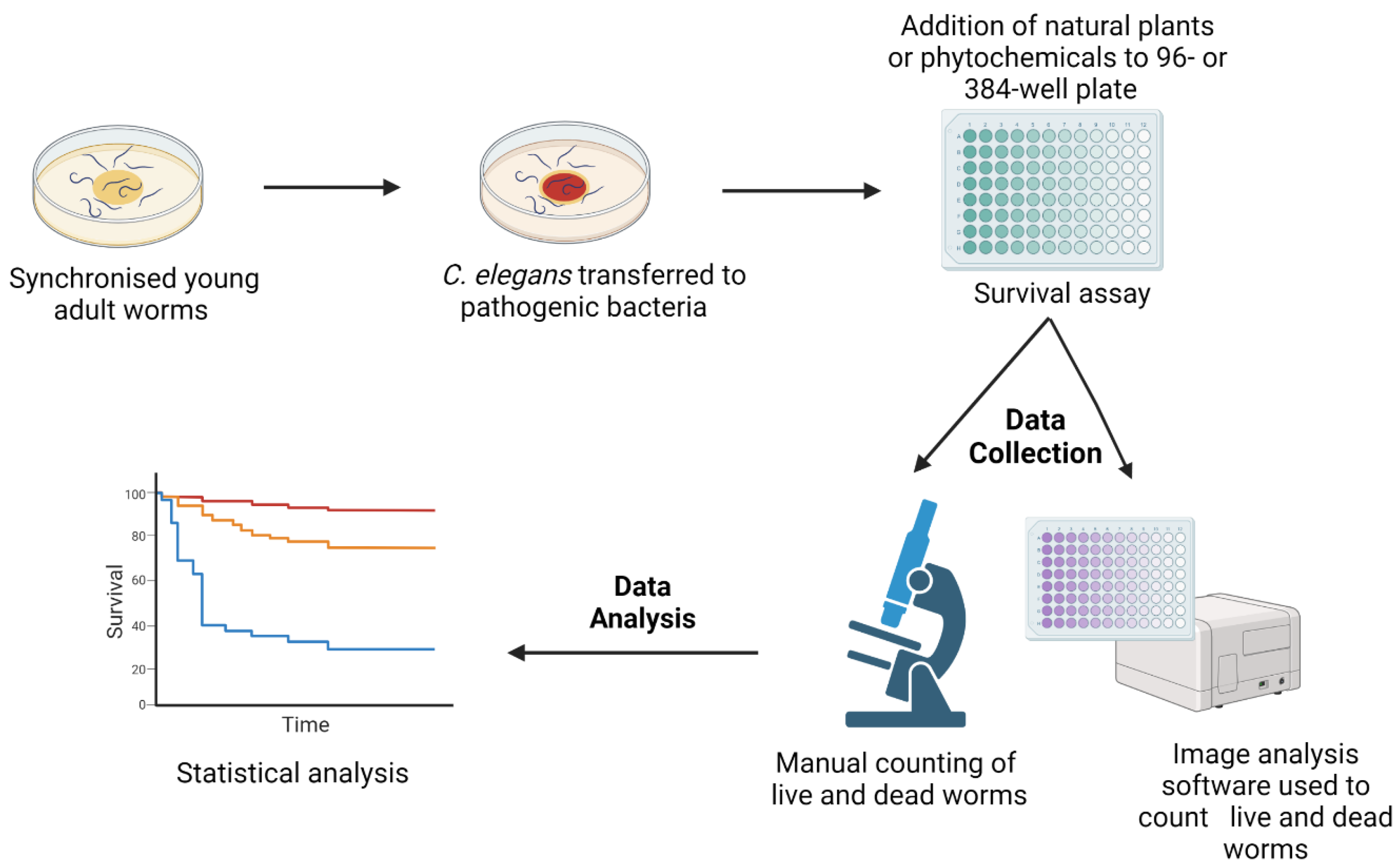
Figure 1. A schematic diagram of the survival assays used for the C. elegans-based screening of novel compounds with antimicrobial activity. Synchronised C. elegans are generated either by egg laying or the bleaching method. Worms are grown in E. coli OP50 plates for 2–3 days at 20 °C until their adulthood. Subsequently, they are transferred to plates seeded with pathogenic microbes. Following an incubation period, worms are transferred to 12-, 24-, 96-, or 384-well assay plates containing different concentrations of antimicrobial compounds. Numbers of live and dead worms can be scored manually under the microscope or by imaging software using a cell permeable dye identifying dead nematodes. The image was created with BioRender.com.
The main advantage of the live animal model is that it not only allows for the assessment of direct antimicrobial effects of defined agents with novel mechanisms of action or molecular targets, but it also addresses their potential role as immune modulators [30][9], indirectly contributing to fighting microbial virulence and increasing the likelihood of host survival. Moreover, the use of a whole live animal model provides valuable information on systemic toxicity of specific compounds in the early stages of drug discovery.
Importantly, the small size and short lifespan of C. elegans convincingly support its suitability as a model organism. Indeed, C. elegans is a 1 mm long, free-living, transparent nematode that develops from a hatched egg to adulthood in 3 days at 20 °C. Once they reach adulthood, wild-type worms live for three additional weeks. The short lifespan and quick generation time accelerate the antimicrobial discovery process and critically favour its automation.
C. elegans was the first multicellular organism whose genome was fully sequenced (C. elegans Sequencing Consortium 1998). Most notably, C. elegans is a self-fertilised organism, and all offspring are genetically identical, thus providing a distinct advantage compared to fruit flies and murine models [31][10]. Indeed, inbreeding of mutant strains to clarify mechanisms of action of novel compounds can be easily performed starting from a highly homogeneous genetic background.
Thus, C. elegans qualifies as an excellent model organism, bridging the gap between in vitro assays and the complexities of vertebrate models.
2. Natural Products Active against Bacterial Infection
Staphylococcus aureus (S. aureus), a Gram-positive bacterium, frequently implicated in various community- and hospital-acquired infections, has developed resistance against penicillin and semisynthetic methicillin. The ability to produce potent toxins and enzymes and form biofilms makes these bacteria highly feared pathogens. Methicillin-resistant S. aureus (MRSA) has become widespread in all settings, and new strains continue to emerge and develop higher resistance to the few available antibiotics [75][11]. A liquid-based assay used to screen novel compounds has shown that natural extracts from Nypa fruticans (mangrove palm), Swietenia macrophylla (big leaf mahogany), Curcuma longa (turmeric), Eurycoma longifolia (longjack), Orthosiphon stamineus (cat’s whiskers), and Silybum eburneum (milk thistle) increase the lifespan of S. aureus-infected C. elegans by 2.8-fold [46][12]. N. fruticans husks and C. longa inhibit S. aureus growth at concentrations <200 μg/mL. Furthermore, extracts from N. fruticans roots, S. macrophylla seeds, and O. stamineus leaves protect worms from MRSA, and reduced colonisation of the nematode intestines by S. aureus was observed following treatment with N. fruticans root and O. stamineus leaf extracts. These results suggest that products from these plants, historically used for their analgesic, anti-inflammatory, and antioxidative effects in Southeast Asia, could promote host immunity and/or decrease bacterial virulence [46][12]. In another study, a herbal formulation used to treat wound infections and derived mainly from leaves and bark of plants such as Azadirachta indica (Neem), Acacia nilotica (Babul), Ocimum sanctum (Tulsi), and Annona squamosa (sugar apple) showed promising activity upon repeated exposure of C. elegans to S. aureus. This formulation, containing the Curcuma longa (turmeric) rhizome and seed oil of Ricinus communis (castor oil), inhibits bacterial growth and production of staphyloxanthin, a quorum-sensing (QS)-regulated pigment, at concentrations ≥0.025% v/v. In addition, it prevents biofilm formation and significantly modifies the bacterial transcriptome by targeting genes associated with haemolysis, virulence, enzyme activity, basic cellular processes, quorum-sensing, and transcriptional regulation [51][13]. P. aeruginosa is another opportunistically resistant bacterium posing major healthcare challenges, due to a range of virulence factors [76][14], with intrinsic and acquired mechanisms including low outer membrane permeability, multidrug efflux pumps, and enzymes. Moreover, P. aeruginosa demonstrates adaptive resistance through persistent biofilm formation in infected tissues [77][15]. Secondary metabolites from endophytic fungi of Alangium, Angelica sinensis (female ginseng), Bupleurum chinense, Herba plantaginis (plantain), Schisandra chinensis (magnolia vine fruit), Herba menthae (mint), and Stephania japonica (snake vine), used in traditional Chinese medicine (TCM), were tested in a liquid-based, slow-killing assay with C. elegans infected with MDR P. aeruginosa. This restudy earch identified 36 extracts prolonging C. elegans survival, and 4 with antimicrobial activity against P. aeruginosa. The 18s rRNA amplicon identified the fungal strains producing these four extracts with in vitro and in vivo activity as Alternaria sp. from Angelica sinensis; Herba plantaginis and Phoma exigua from Alangium sp.; and Aspergillus sydowii from Plantago depressa [54][16]. In another study, polysaccharide extracts from Sophora moorcroftiana seeds, a medicinal shrub endemic to China, were shown to improve the lifespan of C. elegans as well as its reproductive capacity upon P. aeruginosa infection, and they demonstrated specific antibacterial activity [55][17]. Both studies suggest that these natural compounds may enhance host immunity and/or reduce bacterial virulence. Swietenia macrophylla (S. macrophylla), a source of limonoids (modified terpenoids), has shown antimicrobial activity against MRSA [46][12]. Moreover, S. macrophylla seed extracts significantly improve the survival of P. aeruginosa-infected C. elegans. Notably, seed ethyl acetate and methanol extracts do not inhibit P. aeruginosa growth. However, S. macrophylla enhances the expression of the immune modulator gene lys-7, which is known to be suppressed by the QS mechanisms of P. aeruginosa [56,57][18][19]. Modulation of the QS mechanisms by natural products is further supported by studies highlighting the antibacterial effects of CBO. In vitro studies have shown that CBO inhibits elastases A and B and regulates signalling molecules of P. aeruginosa. Importantly, it also decreases the expression and production of other virulence factors, such as chitinase, pyocyanin, exopolysaccharide, and biofilm formation, at subinhibitory concentrations (<3.2%). Treatment of P. aeruginosa-infected C. elegans with 1.6% v/v clove oil improves worm survival in vivo [58,59][20][21]. Similar effects were observed by using Murraya koenigii (curry tree), a medicinal plant with anti-infective and antioxidant properties [60][22]. Marine plants, such as the macroalgae Chondrus crispus (C. crispus), a red seaweed, are rich sources of compounds with immunomodulatory abilities [78][23]. A water extract of C. crispus has been shown to improve the survival of P. aeruginosa-infected C. elegans at a 500 µg/mL concentration. These effects result from both host immune-system activation through the pmk-1, daf-2/daf-16, and skn-1 pathways, and QS and virulence gene suppression [61][24]. Furthermore, an extract of Ascophyllum nodosum, a brown seaweed, promotes the survival of C. elegans through similar mechanisms and by significantly reducing biofilm formation by P. aeruginosa [62][25]. Ayurveda, an ancient holistic approach used for disease prevention and treatment, has also been extensively studied in the context of innovative drug discovery. For instance, TF, a formulation of the three plants Phyllanthus emblica (Indian gooseberry), Terminalia bellerica (Bibhitaka), and Terminalia chebula (Chebulic Myrobalan), is commonly used in Ayurvedic medicine to improve general health and to treat a variety of diseases [79][26]. C. elegans has been used as an in vivo model to test TF antibacterial activity against S. aureus, P. aeruginosa, S. pyogenes, S. marcescens, and C. violaceum [63][27]. All pathogens except S. pyogenes showed reduced virulence (18–45%) towards the nematode host following pretreatment with TF. Moreover, C. elegans pretreated with TF also showed an increased survival rate (14–41%) following infection with these five pathogens. In addition, TF is effective in worms infected with S. marcescens, C. violaceum, or S. aureus. However, TF is ineffective in worms exposed to a mixed population of S. aureus and P. aeruginosa. TF antibacterial activity can be attributed to a combination of reduced QS pigment production in Gram-negative bacteria, virulent enzyme inhibition in S. aureus, and upregulation of the host immune defence accompanied by decreased biofilm formation by S. marcescens and S. aureus. In a similar study, bacteria pretreated with TF leaf extracts showed decreased virulence associated with QS modifications [79][26]. Psoralea corylifolia (Babchi), a herb used in traditional Chinese medicine and Ayurveda to treat skin diseases [80][28], has also been studied for its QS-dampening constituents. Its seed extracts contain bakuchiol, a terpenophenol that inhibits in vitro biofilm formation by P. aeruginosa, A. hydrophila, C. violaceum, S. marcescens, and L. monocytogenes. To study their effects in vivo, C. elegans infected with P. aeruginosa were treated with 1000 µg/mL P. corylifolia seed extract. Worms survived longer than untreated nematodes [64][29]. While increased survival was also observed in a similar study testing Mangifera indica (mango) leaf extracts [65][30], underlying in vivo mechanisms remain to be investigated. Streptococcus pyogenes (S. pyogenes), a Gram-positive, β-haemolytic streptococcus, can cause highly contagious infections with serious complications, such as rheumatic fever. Although first-line treatment with penicillin is still effective, some resistant strains are emerging [81][31]. The bioflavonoid fukugiside from Garcinia travancorica (mangosteen) has shown promising concentration-dependent antibacterial properties against S. pyogenes in vitro by downregulating various virulence genes and reducing its ability to evade phagocytosis. Fukugiside significantly improves the survival rate of C. elegans infected with S. pyogenes without displaying any toxicity [66][32]. Betulin, a triterpenoid present in the bark of birch trees, has also shown similar effects in vivo [82][33]. Interestingly, in an innovative antimicrobial approach, extracts of Tripterygium wilfordii (Thunder God Vine) were able to photosensitise S. pyogenes and S. aureus for the treatment of skin infections. Accordingly, the combination of T. wilfordii extract and antimicrobial photodynamic therapy improved the lifespan of C. elegans infected with S. pyogenes [67][34]. Among pathogens that conventional antimicrobials cannot eradicate [83][35], shigatoxin-producing enterohemorrhagic E coli 0157:H7 causes bloody diarrhoea, which can only be managed supportively. Within this context, the C. elegans model was used to test the antibacterial properties of clove oil, known to downregulate various virulence factors, including QS molecules, and biofilm formation [58,59][20][21]. Indeed, eugenol, the major constituent of clove oil, displays high efficacy against E coli 0157:H7 and prolongs nematode survival [68][36]. Another gut pathogen, Salmonella typhimurium, is susceptible to B. oleracea var. Botrytis (cauliflower) byproduct extracts. Studies in cauliflower-treated C. elegans have shown a dose-dependent reduction in Salmonella typhimurium virulence factors, significantly reduced colonisation, and prolonged nematode survival [69,70][37][38]. In addition to flavonoids, phytoestrogens such as honokiol and magnolol, derived from magnolia species, have demonstrated antibacterial activity [71][39]. The survival of Vibrio cholerae-infected C. elegans can be significantly prolonged by treatment with these polyphenols. In vitro studies suggest that these compounds decrease host inflammatory responses to this potent diarrhoeal pathogen [74][40]. Burkholderia pseudomallei, a tropical pathogen that causes serious melioidosis, shows reduced pathogenicity in vivo when infected C. elegans are treated with curcumin (turmeric). Further investigations have shown that this effect relies on alterations of the bacterial transcriptome, resulting in the attenuation of virulence factors [72][41].3. Natural Products Active against Fungal Infection
Candida albicans (C. albicans), causing opportunistic infections, has represented a major target for the discovery of natural antifungal products. The limited array of antifungal drugs and the intrinsic ability of C. albicans to quickly develop tolerance highlight the significance of the C. elegans infection model in this regard [89][42]. A total of 2560 natural products were screened to identify compounds favouring the survival of C. albicans-infected nematodes, and 12 triterpenoid-based saponins significantly prolonged it. Six of these compounds also inhibited in vitro growth of the yeast, including its resistant strains, and two prevented biofilm formation [87][43]. Saponins are plant glycosides known in traditional medicine for their antitumorigenic, antimicrobial, and anti-inflammatory properties. Although they can be toxic at high concentrations due to their haemolytic properties [90][44], they have shown promising potential as enhancers of photosensitiser uptake in combination with photodynamic treatments [87][43]. Gallic acid and gallates are secondary metabolites present in various plants and are known for their antioxidant and anti-inflammatory properties [91][45]. In vitro studies support their antifungal effects against Candida spp., Cryptococcus spp., Paracoccidioides spp., and Histoplasma capsulatum [92][46]. The in vivo model of C. albicans–C. elegans indicates that they significantly increase the nematode lifespan at >1 µg/mL concentrations, although their toxicity is also increased at high concentrations [84][47]. Another hydrophobic polyphenol, thymol, also prolongs the survival of C. albicans-infected worms and shows antibiofilm activity in vitro. This compound, extracted from thyme, also prevents dysregulation of the host p38 MAPK signalling pathway, playing a key role in the immune response [93][48]. An investigative study identified 15 out of 1266 tested compounds that promoted the longevity of infected worms and prevented the fungal yeast–hyphal transition. They included plant-based compounds, caffeic acid phenethyl ester (CAPE), and lapachol [85][49]. CAPE is a polyphenolic ester naturally occurring in plants, and its bioactive form can be extracted from honeybee hive propolis. Its mechanism of action has been proposed to rely on yeast gene suppression and immunomodulation [94][50]. On the other hand, lapachol, a naphthoquinone derived from the bark of Tabebuia avellaneda (lapacho tree), is used in traditional Amazonian medicine as an antibacterial and antiparasitic agent [45][51]. Both compounds were shown to inhibit C. albicans biofilm formation in vitro, although only CAPE demonstrated antifungal effects in a murine model [85][49]. Fungal filamentation and biofilm formation are key virulence factors accounting for C. albicans pathogenicity by allowing microbes to colonise mucosal surfaces and medical devices, and to invade human tissues. Moreover, the biofilm matrix renders fungi resistant to immune defences and drugs, leading to intractable infections [95][52]. The non-toxic naturally occurring antifungals magnolol and honokiol significantly suppress in vitro yeast cell adhesion, hyphal growth, and biofilm formation at 4–32 μg/mL concentrations by downregulating virulence genes, including those of the Ras1-cAMP-Efg1 pathway. Importantly, both honokiol and magnolol significantly prolong the lifespan of C. albicans-infected worms by preventing colonisation [86][53]. In a similar study, tetrandrine (TET), an alkaloid obtained from the roots of Stephania tetrandra, a medicinal herb known as Fang Ji used in Chinese herbal medicine for joint diseases, inhibited the yeast-to-hyphae transition and biofilm formation in vitro by downregulating the expression of genes from the Ras/cAMP pathway, responsible for QS and biofilm formation [96][54]. Accordingly, TET enhanced the lifespan of infected nematodes at the same concentration range as magnolol and honokiol [88][55]. Major issues related to the use of antifungal agents are associated with their adverse effects and drug-resistance development, since treatment requires long-term administration due to frequent relapses [97][56]. Therefore, natural plants may represent a safe and effective alternative and C. elegans may provide an early detection tool to identify compounds with selective toxicity against fungi and/or unravel alternative mechanisms of action inhibiting fungal infection.References
- Moy, T.I.; Ball, A.R.; Anklesaria, Z.; Casadei, G.; Lewis, K.; Ausubel, F.M. Identification of novel antimicrobials using a live-animal infection model. Proc. Natl. Acad. Sci. USA 2006, 103, 10414–10419.
- Moy, T.I.; Conery, A.L.; Larkins-Ford, J.; Wu, G.; Mazitschek, R.; Casadei, G.; Lewis, K.; Carpenter, A.E.; Ausubel, F.M. High-throughput screen for novel antimicrobials using a whole animal infection model. ACS Chem. Biol. 2009, 4, 527–533.
- Silver, L.L. Challenges of antibacterial discovery. Clin. Microbiol. Rev. 2011, 24, 71–109.
- Adonizio, A.; Leal, S.M.; Ausubel, F.M.; Mathee, K. Attenuation of Pseudomonas aeruginosa virulence by medicinal plants in a Caenorhabditis elegans model system. J. Med. Microbiol. 2008, 57, 809–813.
- Burns, A.R.; Kwok, T.C.; Howard, A.; Houston, E.; Johanson, K.; Chan, A.; Cutler, S.R.; McCourt, P.; Roy, P.J. High-throughput screening of small molecules for bioactivity and target identification in Caenorhabditis elegans. Nat. Protoc. 2006, 1, 1906–1914.
- Tampakakis, E.; Okoli, I.; Mylonakis, E. A C. elegans-based, whole animal, in vivo screen for the identification of antifungal compounds. Nat. Protoc. 2008, 3, 1925–1931.
- Gosai, S.J.; Kwak, J.H.; Luke, C.J.; Long, O.S.; King, D.E.; Kovatch, K.J.; Johnston, P.A.; Shun, T.Y.; Lazo, J.S.; Perlmutter, D.H.; et al. Automated high-content live animal drug screening using C. elegans expressing the aggregation prone serpin alpha1-antitrypsin Z. PLoS ONE 2010, 5, e15460.
- Giunti, S.; Andersen, N.; Rayes, D.; De Rosa, M.J. Drug discovery: Insights from the invertebrate Caenorhabditis elegans. Pharmacol. Res. Perspect. 2021, 9, e00721.
- Peterson, N.D.; Pukkila-Worley, R. Caenorhabditis elegans in high-throughput screens for anti-infective compounds. Curr. Opin. Immunol. 2018, 54, 59–65.
- Phillips, P.C. Self-fertilization sweeps up variation in the worm genome. Nat. Genet. 2012, 44, 237–238.
- Lakhundi, S.; Zhang, K. Methicillin-Resistant Staphylococcus aureus: Molecular Characterization, Evolution, and Epidemiology. Clin. Microbiol. Rev. 2018, 31, 10–1128.
- Kong, C.; Yehye, W.A.; Abd Rahman, N.; Tan, M.W.; Nathan, S. Discovery of potential anti-infectives against Staphylococcus aureus using a Caenorhabditis elegans infection model. BMC Complement Altern. Med. 2014, 14, 4.
- Patel, P.; Joshi, C.; Kothari, V. Anti-Pathogenic Efficacy and Molecular Targets of a Polyherbal Wound- Care Formulation (Herboheal) against Staphylococcus aureus. Infect. Disord. Drug Targets 2019, 19, 193–206.
- Silby, M.W.; Winstanley, C.; Godfrey, S.A.C.; Levy, S.B.; Jackson, R.W. Pseudomonas genomes: Diverse and adaptable. FEMS Microbiol. Rev. 2011, 35, 652–680.
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192.
- Zhou, Y.M.; Shao, L.; Li, J.A.; Han, L.Z.; Cai, W.J.; Zhu, C.B.; Chen, D.J. An efficient and novel screening model for assessing the bioactivity of extracts against multidrug-resistant Pseudomonas aeruginosa using Caenorhabditis elegans. Biosci. Biotechnol. Biochem. 2011, 75, 1746–1751.
- Zhang, Y.; Mi, D.Y.; Wang, J.; Luo, Y.P.; Yang, X.; Dong, S.; Ma, X.M.; Dong, K.Z. Constituent and effects of polysaccharides isolated from Sophora moorcroftiana seeds on lifespan, reproduction, stress resistance, and antimicrobial capacity in Caenorhabditis elegans. Chin. J. Nat. Med. 2018, 16, 252–260.
- Dharmalingam, K.; Tan, B.K.; Mahmud, M.Z.; Sedek, S.A.; Majid, M.I.; Kuah, M.K.; Sulaiman, S.F.; Ooi, K.L.; Khan, N.A.; Muhammad, T.S.; et al. Swietenia macrophylla extract promotes the ability of Caenorhabditis elegans to survive Pseudomonas aeruginosa infection. J. Ethnopharmacol. 2012, 139, 657–663.
- Evans, E.A.; Kawli, T.; Tan, M.W. Pseudomonas aeruginosa suppresses host immunity by activating the DAF-2 insulin-like signaling pathway in Caenorhabditis elegans. PLoS Pathog. 2008, 4, e1000175.
- Haripriyan, J.; Omanakuttan, A.; Menon, N.D.; Vanuopadath, M.; Nair, S.S.; Corriden, R.; Nair, B.G.; Nizet, V.; Kumar, G.B. Clove Bud Oil Modulates Pathogenicity Phenotypes of the Opportunistic Human Pathogen Pseudomonas aeruginosa. Sci. Rep. 2018, 8, 3437.
- Husain, F.M.; Ahmad, I.; Asif, M.; Tahseen, Q. Influence of clove oil on certain quorum-sensing-regulated functions and biofilm of Pseudomonas aeruginosa and Aeromonas hydrophila. J. Biosci. 2013, 38, 835–844.
- Ganesh, P.S.; Rai, R.V. Inhibition of quorum-sensing-controlled virulence factors of Pseudomonas aeruginosa by Murraya koenigii essential oil: A study in a Caenorhabditis elegans infectious model. J. Med. Microbiol. 2016, 65, 1528–1535.
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597.
- Liu, J.; Hafting, J.; Critchley, A.T.; Banskota, A.H.; Prithiviraj, B. Components of the cultivated red seaweed Chondrus crispus enhance the immune response of Caenorhabditis elegans to Pseudomonas aeruginosa through the pmk-1, daf-2/daf-16, and skn-1 pathways. Appl. Environ. Microbiol. 2013, 79, 7343–7350.
- Kandasamy, S.; Khan, W.; Evans, F.; Critchley, A.T.; Prithiviraj, B. Tasco(R): A product of Ascophyllum nodosum enhances immune response of Caenorhabditis elegans against Pseudomonas aeruginosa infection. Mar. Drugs 2012, 10, 84–105.
- Bag, A.; Bhattacharyya, S.K.; Pal, N.K. Antibacterial potential of hydroalcoholic extracts of triphala components against multidrug-resistant uropathogenic bacteria—A preliminary report. Indian J. Exp. Biol. 2013, 51, 709–714.
- Patel, H.; Patel, F.; Jani, V.; Jha, N.; Ansari, A.; Paliwal, B.; Rathod, B.; Patel, D.; Patel, P.; Kothari, V. Anti-pathogenic potential of a classical ayurvedic Triphala formulation. F1000Res 2019, 8, 1126.
- Khushboo, P.S.; Jadhav, V.M.; Kadam, V.J.; Sathe, N.S. Psoralea corylifolia Linn.-“Kushtanashini”. Pharmacogn. Rev. 2010, 4, 69–76.
- Husain, F.M.; Ahmad, I.; Khan, F.I.; Al-Shabib, N.A.; Baig, M.H.; Hussain, A.; Rehman, M.T.; Alajmi, M.F.; Lobb, K.A. Seed Extract of Psoralea corylifolia and Its Constituent Bakuchiol Impairs AHL-Based Quorum Sensing and Biofilm Formation in Food- and Human-Related Pathogens. Front. Cell. Infect. Microbiol. 2018, 8, 351.
- Husain, F.M.; Ahmad, I.; Al-Thubiani, A.S.; Abulreesh, H.H.; AlHazza, I.M.; Aqil, F. Leaf Extracts of Mangifera indica L. Inhibit Quorum Sensing—Regulated Production of Virulence Factors and Biofilm in Test Bacteria. Front. Microbiol. 2017, 8, 727.
- Kanwal, S.; Vaitla, P. Streptococcus Pyogenes; StatPearls: Treasure Island, FL, USA, 2023.
- Nandu, T.G.; Subramenium, G.A.; Shiburaj, S.; Viszwapriya, D.; Iyer, P.M.; Balamurugan, K.; Rameshkumar, K.B.; Karutha Pandian, S. Fukugiside, a biflavonoid from Garcinia travancorica inhibits biofilm formation of Streptococcus pyogenes and its associated virulence factors. J. Med. Microbiol. 2018, 67, 1391–1401.
- Viszwapriya, D.; Subramenium, G.A.; Prithika, U.; Balamurugan, K.; Pandian, S.K. Betulin inhibits virulence and biofilm of Streptococcus pyogenes by suppressing ropB core regulon, sagA and dltA. Pathog. Dis. 2016, 74, ftw088.
- Alam, S.T.; Hwang, H.; Son, J.D.; Nguyen, U.T.T.; Park, J.S.; Kwon, H.C.; Kwon, J.; Kang, K. Natural photosensitizers from Tripterygium wilfordii and their antimicrobial photodynamic therapeutic effects in a Caenorhabditis elegans model. J. Photochem. Photobiol. B 2021, 218, 112184.
- Vaou, N.; Stavropoulou, E.; Voidarou, C.; Tsigalou, C.; Bezirtzoglou, E. Towards Advances in Medicinal Plant Antimicrobial Activity: A Review Study on Challenges and Future Perspectives. Microorganisms 2021, 9, 2041.
- Kim, Y.G.; Lee, J.H.; Gwon, G.; Kim, S.I.; Park, J.G.; Lee, J. Essential Oils and Eugenols Inhibit Biofilm Formation and the Virulence of Escherichia coli O157:H7. Sci. Rep. 2016, 6, 36377.
- Ibáñez-Peinado, D.; Pina-Pérez, C.; García-Carrión, G.; Martínez, A.; Rodrigo, D. In vivo Antimicrobial Activity Assessment of a Cauliflower By-Product Extract Against Salmonella Typhimurium. Front. Sustain. Food Syst. 2020, 4, 8.
- Sanz-Puig, M.; Lazaro, E.; Armero, C.; Alvares, D.; Martinez, A.; Rodrigo, D.S. Typhimurium virulence changes caused by exposure to different non-thermal preservation treatments using C. elegans. Int. J. Food Microbiol. 2017, 262, 49–54.
- Jacobo-Salcedo Mdel, R.; Gonzalez-Espindola, L.A.; Alonso-Castro, A.J.; Gonzalez-Martinez Mdel, R.; Dominguez, F.; Garcia-Carranca, A. Antimicrobial activity and cytotoxic effects of Magnolia dealbata and its active compounds. Nat. Prod. Commun. 2011, 6, 1121–1124.
- Kim, H.-I.; Kim, J.-A.; Choi, E.-J.; Harris, J.B.; Jeong, S.-Y.; Son, S.-J.; Kim, Y.; Shin, O.S. In vitro and in vivo antimicrobial efficacy of natural plant-derived compounds against Vibrio cholerae of O1 El Tor Inaba serotype. Biosci. Biotechnol. Biochem. 2015, 79, 475–483.
- Eng, S.A.; Nathan, S. Curcumin rescues Caenorhabditis elegans from a Burkholderia pseudomallei infection. Front. Microbiol. 2015, 6, 290.
- Costa-de-Oliveira, S.; Rodrigues, A.G. Candida albicans Antifungal Resistance and Tolerance in Bloodstream Infections: The Triad Yeast-Host-Antifungal. Microorganisms 2020, 8, 154.
- Coleman, J.J.; Okoli, I.; Tegos, G.P.; Holson, E.B.; Wagner, F.F.; Hamblin, M.R.; Mylonakis, E. Characterization of plant-derived saponin natural products against Candida albicans. ACS Chem. Biol. 2010, 5, 321–332.
- Podolak, I.; Galanty, A.; Sobolewska, D. Saponins as cytotoxic agents: A review. Phytochem. Rev. 2010, 9, 425–474.
- Rosas, E.C.; Correa, L.B.; das Graças Henriques, M. Chapter 28—Antiinflammatory Properties of Schinus terebinthifolius and Its Use in Arthritic Conditions. In Bioactive Food as Dietary Interventions for Arthritis and Related Inflammatory Diseases, 2nd ed.; Watson, R.R., Preedy, V.R., Watson, R.R., Preedy, V.R., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 489–505.
- De Paula, E.S.A.C.; Costa-Orlandi, C.B.; Gullo, F.P.; Sangalli-Leite, F.; de Oliveira, H.C.; da Silva Jde, F.; Scorzoni, L.; Pitangui Nde, S.; Rossi, S.A.; Benaducci, T.; et al. Antifungal Activity of Decyl Gallate against Several Species of Pathogenic Fungi. Evid. Based Complement. Altern. Med. 2014, 2014, 506273.
- Singulani, J.L.; Scorzoni, L.; Gomes, P.C.; Nazare, A.C.; Polaquini, C.R.; Regasini, L.O.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J.S. Activity of gallic acid and its ester derivatives in Caenorhabditis elegans and zebrafish (Danio rerio) models. Future Med. Chem. 2017, 9, 1863–1872.
- Shu, C.; Sun, L.; Zhang, W. Thymol has antifungal activity against Candida albicans during infection and maintains the innate immune response required for function of the p38 MAPK signaling pathway in Caenorhabditis elegans. Immunol. Res. 2016, 64, 1013–1024.
- Breger, J.; Fuchs, B.B.; Aperis, G.; Moy, T.I.; Ausubel, F.M.; Mylonakis, E. Antifungal chemical compounds identified using a C. elegans pathogenicity assay. PLoS Pathog. 2007, 3, e18.
- Murtaza, G.; Karim, S.; Akram, M.R.; Khan, S.A.; Azhar, S.; Mumtaz, A.; Bin Asad, M.H. Caffeic acid phenethyl ester and therapeutic potentials. Biomed. Res. Int. 2014, 2014, 145342.
- Cragg, G.; Newman, D. Natural products and drug discovery and development: A history of success and continuing promise for the future. Planta Med. 2014, 80, IL1.
- Gulati, M.; Nobile, C.J. Candida albicans biofilms: Development, regulation, and molecular mechanisms. Microbes Infect. 2016, 18, 310–321.
- Sun, L.; Liao, K.; Wang, D. Effects of Magnolol and Honokiol on Adhesion, Yeast-Hyphal Transition, and Formation of Biofilm by Candida albicans. PLoS ONE 2015, 10, e0117695.
- Ahmad Khan, M.S.; Altaf, M.M.; Sajid, M. Chapter 14—Insights of Phyto-Compounds as Antipathogenic Agents: Controlling Strategies for Inhibiting Biofilms and Quorum Sensing in Candida albicans. In New Look to Phytomedicine; Ahmad Khan, M.S., Ahmad, I., Chattopadhyay, D., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 367–389.
- Zhao, L.X.; Li, D.D.; Hu, D.D.; Hu, G.H.; Yan, L.; Wang, Y.; Jiang, Y.Y. Effect of Tetrandrine against Candida albicans Biofilms. PLoS ONE. 2013, 8, e79671.
- Benson, J.M.; Nahata, M.C. Clinical use of systemic antifungal agents. Clin. Pharm. 1988, 7, 424–438.
More