The intake of linoleic acid (LA) has increased dramatically in the standard American diet. LA is generally promoted as supporting human health, but there exists controversy regarding whether the amount of LA currently consumed in the standard American diet supports human health. While LA is considered to be an essential fatty acid and support health when consumed in modest amounts, an excessive intake of LA leads to the formation of oxidized linoleic acid metabolites (OXLAMs), impairments in mitochondrial function through suboptimal cardiolipin composition, and likely contributes to many chronic diseases that became an epidemic in the 20th century, and whose prevalence continues to increase. As LA consumption increases, the potential for OXLAM formation also increases. OXLAMs have been associated with various illnesses, including cardiovascular disease, cancer, and Alzheimer’s disease, among others. Lowering dietary LA intake can help reduce the production and accumulation of OXLAMs implicated in chronic diseases.
- linoleic acid (LA)
- seed oils
- cardiolipin
- oxidized linoleic acid metabolites (OXLAMs)
- 4-hydroxynonenal (HNE)
- omega-3
- omega-6
1. Introduction
2. How Much LA Is Required in the Human Diet?
The first demonstration of the essential requirement for LA in animal diets was obtained by Burr and Burr in 1929–1930 [13]. They showed that rats receiving 0.6% of their total dietary calories as LA were 30% higher in body weight compared to total-fat-deficient rats and did not develop skin desquamation and tail necrosis [17,18,19][17][18][19]. This led to the establishment of the requirement of 1% of the total daily calories being omega-6 fat, which was later extended to 2% of the total daily calorie intake in humans to ensure sufficiency. This was confirmed by two studies where the physiological symptoms of omega-6 deficiency in human infants, as established by scaling of the skin, were abolished [20]. However, a careful review of the data used to establish LA being essential to the diet found that this conclusion was established using control diets that were not only deficient in omega-6 fatty acids, but also omega-3 fatty acids. This dual deficiency in the control diets seemed to invalidate the establishment of an omega-6 fatty acid requirement from these data [21]. Subsequent studies in rat models similar to those used to originally classify LA as an essential fatty acid have demonstrated that the dietary omega-3 fatty acid, alpha-linolenic acid (ALA), is able to diminish the symptoms of LA deficiency [22]. This strongly suggests that the absence of ALA in the original studies probably heightened the significance of the physiological symptoms caused by LA deficiency. It seems that, at least for the rat model, the nutritional requirement for LA has probably been seriously overestimated. A more precise estimation of the LA requirement is likely closer to a 75% reduction, or 0.5%, of the dietary energy rather than 2% [23].3. The Omega 3:6 Ratio
Among the many types of omega-3 fatty acids, the three most important are eicosapentaenoic acid (EPA), docosahexaenoic acid (DHA), which are “long-chain” omega-3 fatty acids, and alpha linoleic acid (ALA)—a “short-chain” omega-3 fatty acid. The human body is unable to produce essential fatty acids endogenously. Therefore, omega-3 fatty acids must be consumed regularly through the diet. Animal foods such as cold-water fish are ideal sources of EPA and DHA, while ALA is mainly derived from plants. EPA and DHA have anti-inflammatory effects and ALA must be converted to EPH and DHA. This conversion is an inefficient process, particularly in men, with less than 20% being converted [45][24]. Therefore, special attention should be paid to obtaining EPA and DHA through diet or supplementation. The benefits of maintaining the proper omega 3:6 ratio are well-established. Bodily tissues consist primarily of saturated and monounsaturated fats, which are a readily utilized source of nutrients that support the development and maintenance of cells [46][25]. The primary dietary PUFAs are omega-3 and omega-6 fats, which the body needs in relatively small quantities. It is important to consume sufficient amounts of omega-3 fatty acids to sustain optimal health, with the recommended daily serving being between 500 and 1000 milligrams of omega-3 fatty acids [47,48][26][27]. Contrary to previous perceptions, however, consuming larger servings of omega-3 fatty acids does not support an ideal ratio. Instead, excessive quantities of omega-3s may cause additional metabolic damage—similar to that which occurs due to the conversion of elevated LA levels.4. Pathophysiological Mechanism of Elevated LA Levels
As revealed in Figure 1, the primary reason why excess LA in the body leads to cellular tissue damage is because this highly fragile PUFA is easily altered through oxidation. Like other types of PUFAs, LA consists of fragile double bonds that are susceptible to oxidative damage [54][28].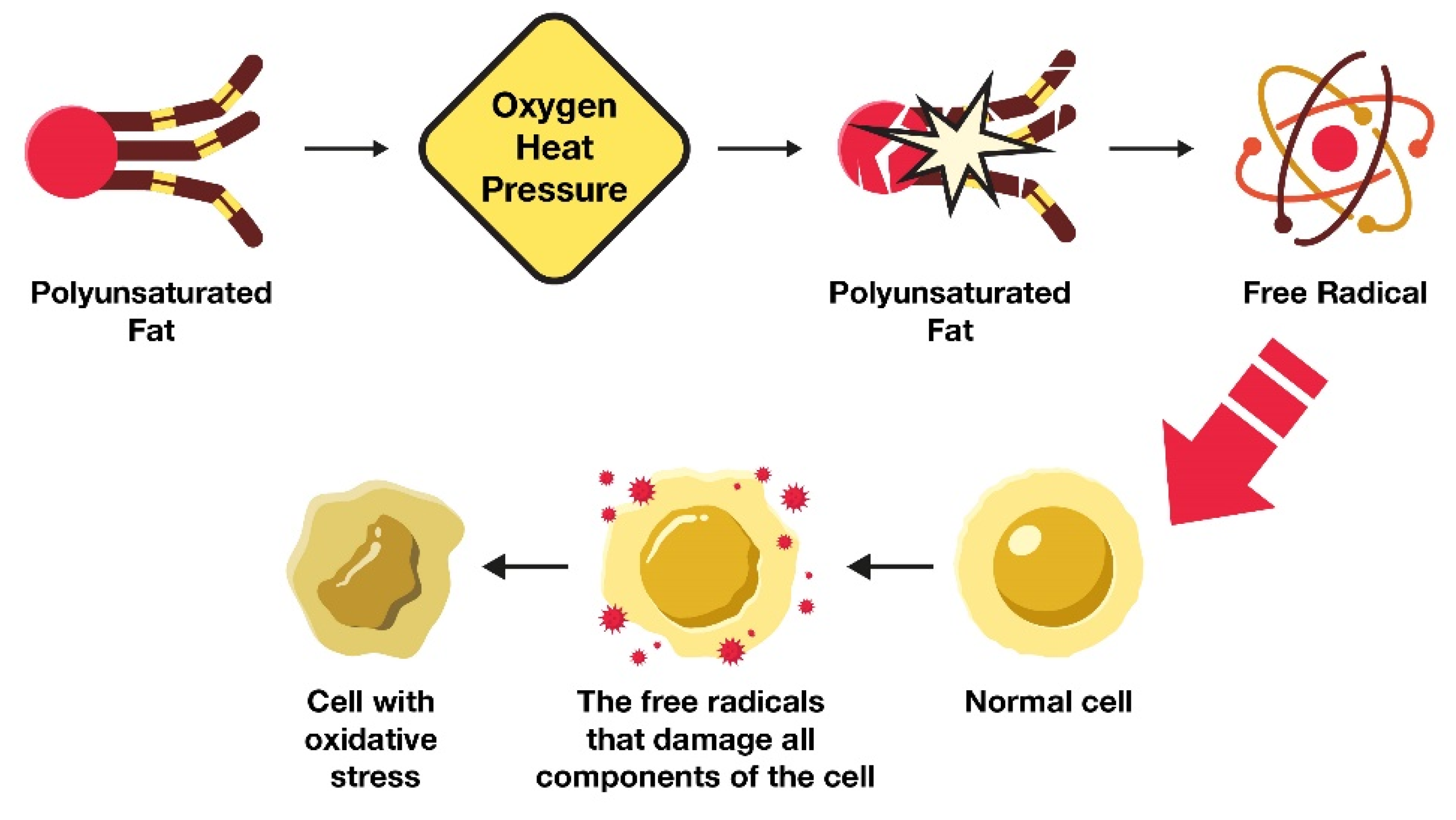
4.1. How Excess LA Consumption Alters Health
4.2. LA Remains in Tissues for Extended Time Periods
Another major reason why seed oils are pernicious to overall health is that they remain in the body for extended periods. The half-life of LA is approximately 680 days, or approximately two years [79][36]. This means that it takes approximately six years to replace 95% of the LA in the body with healthy fats—making this a primary reason to maintain low LA intake. Omega-3 fatty acids, such as DHA and EPA, have half-lives of 2.5 years and a few months, respectively [80,81][37][38]. The body also converts certain amounts of DHA to EPA.4.3. Cardiolipin: Stealth Fat in Mitochondria
Mitochondria are subcellular organelles that are responsible for producing most of the body’s cellular energy in the form of adenosine triphosphate (ATP) [83][39]. It is the presence of mitochondria that distinguishes mammals (e.g., humans) from bacteria and allow life to be multicellular. These organelles produce approximately 85% of the energy for the body through the generation of ATP during oxidative phosphorylation. If mitochondrial dysfunction develops, physical symptoms (e.g., chronic fatigue) may arise, along with an increased susceptibility to disease. It is vital to take preventive measures to improve and maintain mitochondrial health as it profoundly impacts longevity. Cardiolipin optimization reinforces enhanced mitochondria activity and energy production. To emphasize the importance of this phospholipid, 20% of the fat in the mitochondria is found in the form of cardiolipin [85][40]. The human body has over 100,000 trillion mitochondria, and mitochondrial health is largely dependent upon the type of dietary fats that are available for cardiolipin synthesis within these specialized organelles. Cardiolipin is composed of four fatty acids [86][41], unlike triglycerides, which have three fatty acids, but the individual fats that comprise cardiolipin vary widely. Examples include LA, palmitic acid, and the fatty acids found in fish oil, such as DHA and EPA. Cardiolipin is synthesized from fatty acids that are consumed through the diet; thus, the overconsumption of LA in the form of seed oils can alter the formation of the inner mitochondrial membrane, cristae, and complex IV. Cardiolipin molecules containing one or more LA fatty acids are highly susceptible to free-radical-induced lipid peroxidation. The oxidation of cardiolipin is involved in regulating apoptosis, mitophagy, and other cellular functions. Interestingly, LA-containing cardiolipin is preferentially oxidized over other phospholipids in the inner mitochondrial membrane, even in the presence of more-oxidizable fatty acids such as arachidonic acid. This is important, because mitochondrial lipids are necessary for maintaining the structural integrity and proper functioning of mitochondria. Cardiolipin is prone to free-radical-induced lipid peroxidation due to the presence of up to four chains of LA. The oxidation of cardiolipin plays an important role in the regulation of apoptosis, mitophagy, and other cellular functions. The oxidation of cardiolipin plays an important role in the regulation of apoptosis, mitophagy, and other cellular functions. LA-containing cardiolipin is preferentially oxidized over other phospholipids that are found in the inner mitochondrial membrane, including phosphatidylinositol, phosphatidylserine, phosphatidylinositol, and phosphatidylethanolamine. This occurs even in the presence of more oxidizable fatty acids, such as arachidonic acid. Emerging evidence suggests that mitochondrial lipid peroxidation not only affects the structural integrity of mitochondria, but also mitochondrial functions, such as protein transportation, respiratory metabolism for ATP generation, mitochondrial dynamics and quality control through the fission and fusion of mitochondria, and mitophagy. Mitochondrial proteins, which constitute a significant portion of proteins within the inner mitochondrial membrane, are highly susceptible to modification by 4-HNE. Approximately thirty percent of all proteins modified by 4-HNE are mitochondrial proteins. Consequently, endogenously produced 4-HNE resulting from oxidative stress has been shown to cause mitochondrial dysfunction in various cell types and organs during both physiological and pathological conditions. This vulnerability of mitochondrial proteins to 4-HNE modification contributes to their critical role in the development of mitochondrial dysfunction.5. Associations between LA Intake and Chronic Disease
Oxidative stress, tissue damage, and mitochondrial dysfunction from excess LA is not only responsible for cardiovascular disease and Alzheimer’s onset, but additional chronic diseases, including cancer, dementia, obesity, and diabetes are also associated with oxidized metabolites. There is conflicting evidence on the associations between LA intake and many of these chronic diseases, which are summarized below in Table 1.| Proposed Neutral or Health Benefits | Health Risks | ||||
|---|---|---|---|---|---|
| Reduces cardiovascular disease risk by decreasing total cholesterol levels [88] | Reduces cardiovascular disease risk by decreasing total cholesterol levels [42] | Increases risk of cardiovascular disease by increasing oxidized LDL [2,89] | Increases risk of cardiovascular disease by increasing oxidized LDL [2][43] | ||
| Unrecognized as having an impact on cancer [90] | Unrecognized as having an impact on cancer [44] | Increases risk of cancer by impairing mitochondrial function and increasing systemic oxidative stress [91] that adversely impacts cardiolipin in the inner mitochondrial membrane [92,93, | Increases risk of cancer by impairing mitochondrial function and increasing systemic oxidative stress [ | 94] | 45] that adversely impacts cardiolipin in the inner mitochondrial membrane [46][47][48] |
| Reduces the risk of type 2 diabetes [95] | Reduces the risk of type 2 diabetes [49] | Increases risk of diabetes [91] | Increases risk of diabetes [45] | ||
| Role in obesity is contentious [96] | Role in obesity is contentious [50] | Increases risk of obesity [97] | Increases risk of obesity [51] | ||
| Unrecognized as having an impact on dementia | Increases risk of dementia [98] | Increases risk of dementia [52] |
6. Dietary Sources of LA and Mitigation Strategies
In general, the lowest LA-containing source of fats would be the preferred fats of choice for lowering the LA burden in the diet. Olive oil is a popular cooking oil that is prominently featured in Mediterranean diets, which generally contain far fewer seed oils considering the abundant use of olive oil.
However, olive oil demonstrated a nearly 10-fold wide variability in the percentage of LA and the vast majority of commercial olive oil, and avocado oils, are adulterated with seed oils. A recent study evaluated 89 olive cultivars and found a range of 3% to 27% levels of LA [142][58]. Tests have also revealed that anywhere from 60 to 90% of the olive oils sold in American grocery stores and restaurants are adulterated with cheap, oxidized, omega-6 vegetable oils, such as sunflower oil or peanut oil, or nonhuman-grade olive oils, which are harmful to health in a number of ways [143][59].Although this problem is concerning, instead of avoiding all cooking oils (e.g., vegetable and seed oil), healthier choices include those that have been used for centuries, such as butter and beef tallow. In addition to containing the lowest LA content, these sources of fats also provide the fat-soluble vitamins A, D, and K2. Despite these sources being readily available, most Americans simply fail to obtain enough preformed vitamin A in their diet. This can contribute to many chronic diseases, including cardiovascular disease [144][60] and cancers [145][61]. An elevated seed oil intake also adversely impacts health, survival, and vision [146][62]. Coconut oil is also very low in LA but does not have the essential fat-soluble vitamins that tallow and butter contain.
6.1. Sources of Animal Protein and Varying LA Contents
Animals are typically fed corn, soy, or other seeds and grains, which is radically different from their native, traditional, or ancestral diet. This presents a problem for nonruminant animals due to the concentration of LA in the seeds and grains they are fed. Ruminants are animals with multiple parts to their stomachs. This includes cows, buffalo, sheep, lamb, goats, deer, elk, and many other game animals. Ruminants have low LA in both their meat and milk, no matter what they eat [149][63]. This is because their stomach has a ‘biohydrogenation chamber’ that contains bacteria that can convert the high LA fat they eat from grains and seeds into saturated and monounsaturated fats. This is in contrast to animals with one stomach, such as chickens and pigs, that when fed a diet high in LA, including corn and soy, experience an increase in high levels of LA in their tissues, similar to the process that has been observed in humans [150][64]. The ability of the biohydrogenation chamber to efficiently convert high LA fat to saturated and monounsaturated fats is well known, as the difference in LA in ruminants that are 100% grass-fed and those that are fed corn and soy is only approximately 0.5%. This is why, from an LA intake perspective, there is not much difference between concentrated animal feeding operation (CAFO) beef and grass-fed-only beef. However, grass-fed beef is preferred, as it contains less glyphosate, other toxins, and hormones.6.2. Carnosine Helps Lower Oxidative Damage from LA
Carnosine is an endogenously produced dipeptide, and it consists of only two amino acids, beta-alanine and histidine [151][65]. It is a potent antioxidant that helps limit the damage from excess LA by binding to ALEs. It serves as a sacrificial sink for reactive oxygen species (ROS) and ALEs [152][66] by letting these damaging molecules destroy it rather than mitochondria, DNA, or proteins, as depicted in Figure 52.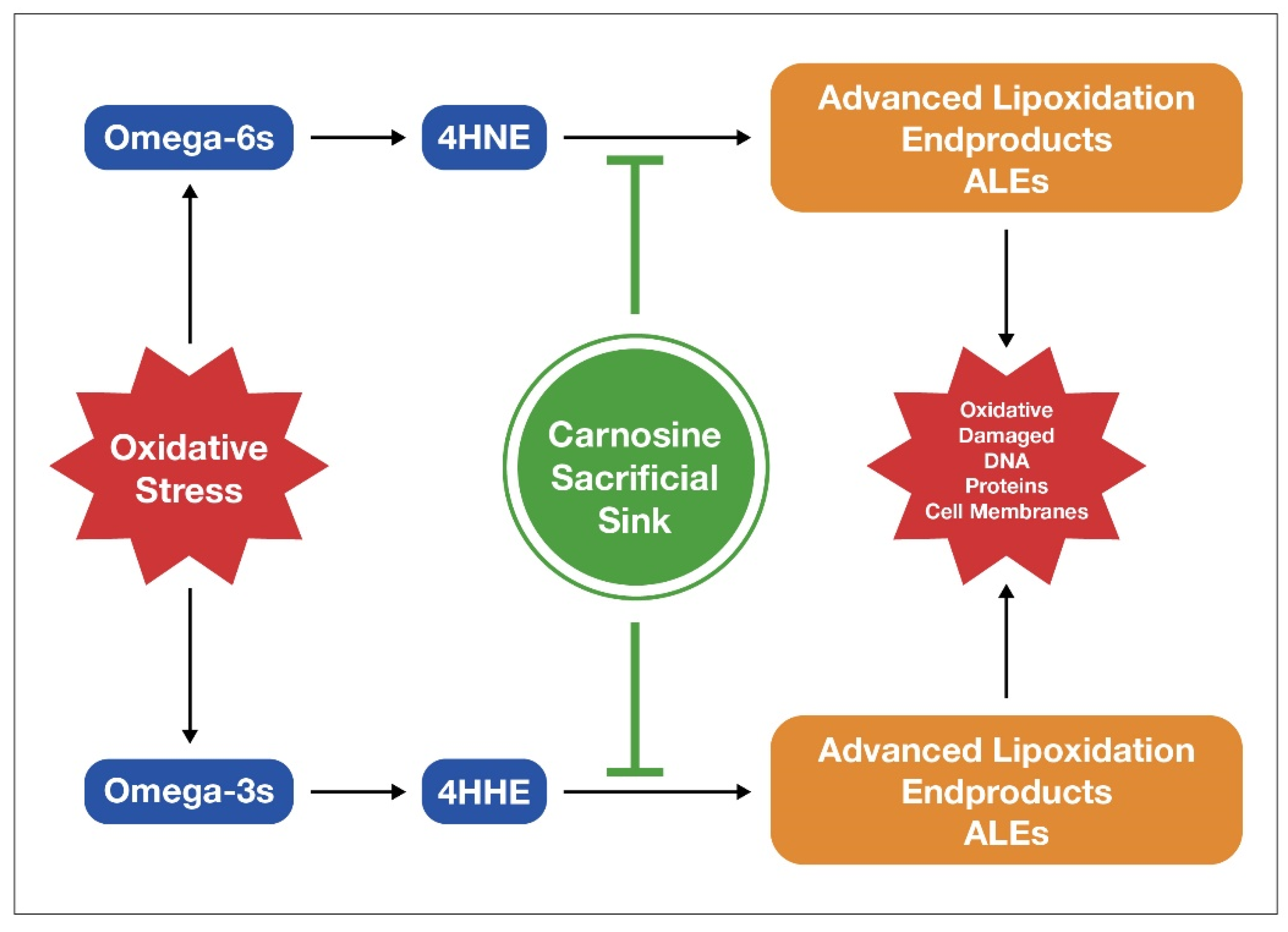
References
- Chapkin, R.S.; McMurray, D.N.; Davidson, L.A.; Patil, B.S.; Fan, Y.Y.; Lupton, J.R. Bioactive dietary long-chain fatty acids: Emerging mechanisms of action. Br. J. Nutr. 2008, 100, 1152–1157.
- Ramsden, C.E.; Ringel, A.; Feldstein, A.E.; Taha, A.Y.; MacIntosh, B.A.; Hibbeln, J.R.; Majchrzak-Hong, S.F.; Faurot, K.R.; Rapoport, S.I.; Cheon, Y.; et al. Lowering dietary linoleic acid reduces bioactive oxidized linoleic acid metabolites in humans. Prostaglandins Leukot. Essent. Fat. Acids 2012, 87, 135–141.
- Holman, R.T.; Johnson, S.B.; Kokmen, E. Deficiency of essential fatty acids and membrane fluidity during pregnancy and lactation. Proc. Natl. Acad. Sci. USA 1968, 61, 1053–1059.
- Behrman, R.E.; Reller, L.B.; Carey, H.V. Essential fatty acid deficiency in a patient receiving prolonged intra-venous alimentation. N. Engl. J. Med. 1971, 285, 19–21.
- Klein, P.D.; Johnson, R.M. Phosphorous metabolism in unsaturated fatty acid-deficient rats. J. Biol. Chem. 1954, 211, 103–110.
- Hayashida, T.; Portman, O.W. Swelling of liver mitochondria from rats fed diets deficient in essential fatty acids. Proc. Soc. Exp. Biol. Med. 1960, 103, 656–659.
- Cornwell, D.G.; Panganamala, R.V. Atherosclerosis an intracellular deficiency in essential fatty acids. Prog. Lipid Res. 1981, 20, 365–376.
- Smith, E.B. The effects of age and of early atheromata on the intimal lipids in men. Biochem. J. 1962, 84, 49.
- Smith, E.B. Lipids carried by S1 0–12 lipoprotein in normal and hypercholesterolaemic serum. Lancet 1962, 2, 530–534.
- Das, U.N. Bioactive lipids and vascular disease. Eur. J. Clin. Nutr. 2021, 75, 1528–1531.
- Salem, N.; Wegher, B.; Mena, P.; Uauy, R. Arachidonic and docosahexaenoic acids are biosynthesized from their 18-carbon precursors in human infants. Proc. Natl. Acad. Sci. USA 1996, 93, 49–54.
- Ramsden, C.E.; Zamora, D.; Faurot, K.R.; MacIntosh, B.; Horowitz, M.; Keyes, G.S.; Yuan, Z.X.; Miller, V.; Lynch, C.; Honvoh, G.; et al. Dietary alteration of n-3 and n-6 fatty acids for headache reduction in adults with migraine: Randomized controlled trial. BMJ 2021, 374, n1448.
- Burr, G.O.; Burr, M.M. A new deficiency disease produced by the rigid exclusion of fat from the diet. J. Biol. Chem. 1929, 82, 345–367.
- Suzuki, N.; Sawada, K.; Takahashi, I.; Matsuda, M.; Fukui, D.; Tokuyasu, H.; Shimizu, H.; Yokoyama, J.; Akaike, A. Association between polyunsaturated fatty acid and reactive oxygen species production of neutrophils in the general population. Nutrients 2020, 12, 3222.
- Santoro, N.; Caprio, S.; Giannini, C.; Kim, G.; Kursawe, R.; Pierpont, B.; Shaw, M.M.; Feldstein, A.E. Oxidized fatty acids: A potential pathogenic link between fatty liver and type-2 diabetes in obese adolescents. Antioxid. Redox Signal. 2014, 20, 383–389.
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative stress: Harms and benefits for human health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763.
- Osborne, T.; Mendel, L.B. Growth on diets poor in true fats. JBC 1920, 45, 145–152.
- Spector, A.A.; Kim, H.Y. Discovery of fatty acids. J. Lipid. Res. 2015, 56, 11–21.
- Hansen, A.E. Essential fatty acids and infant nutrition; Borden award address. Pediatrics 1958, 21, 494–501.
- Hansen, A.E.; Wiese, H.F.; Boelsche, A.N. Role of linoleic acid in infant nutrition study Clinical and Chemical Study of 428 Infants Fed on Milk Mixtures Varying in Kind and Amount of Fat. Pediatrics 1963, 31, 171–192.
- Cunnane, S.C.; Guesnet, P. Linoleic acid recommendations—A house of cards. Prostaglandins Leukot. Essent Fat. Acids 2011, 85, 399–402.
- Igarashi, M.; Gao, F.; Kim, H.W.; Ma, K.; Bell, J.M.; Rapoport., S.I. Dietary n-6 PUFA deprivation for 15 weeks reduces arachidonic acid concentrations while increasing n-3 PUFA concentrations in organs of post-weaning male rats. Biochim. Biophys. Acta 2009, 1791, 132–139.
- Guesnet, P.; Lallemand, S.M.; Alessandri, J.M.; Jouin, M.; Cunnane, S.C. α-Linolenate reduces the dietary requirement for linoleate in the growing rat. Prostaglandins Leukot. Essent Fat. Acids 2011, 85, 353–360.
- Stark, A.H.; Crawford, M.A.; Reifen, R. Update on alpha-linolenic acid. Nutr. Rev. 2008, 66, 326–333.
- DiNicolantonio, J.J.; O’Keefe, J.H. Monosaturated fat vs. saturated fat: Effects on cardio-metabolic health and obesity. Mo. Med. 2022, 119, 69–73.
- Vannice, G.; Rasmussen, H. Position of the Academy of Nutrition and Dietetics: Dietary fatty acids for healthy adults. J. Acad. Nutr. Diet. 2014, 114, 136–153.
- Kris-Etherton, P.M. Fish consumption, fish oil, omega-3 Fatty Acids, and cardiovascular disease. Circulation 2002, 106, 2747–2757.
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438.
- Hoxha, M.; Spahiu, E.; Prendi, E.; Zappacosta, B. A Systematic Review on the Role of Arachidonic Acid Pathway in Multiple Sclerosis. CNS Neurol. Disord. Drug Targets 2022, 21, 160–187.
- Birkic, N.; Azar, T.; Maddipati, K.R.; Minic, Z.; Reynolds, C.A. Excessive dietary linoleic acid promotes plasma accumulation of pronociceptive fatty acyl lipid mediators. Sci. Rep. 2022, 12, 17832.
- Singh, P.N.; Arthur, K.N.; Orlich, M.J.; James, W.; Purty, A.; Job, J.S.; Rajaram, S.; Sabaté, J. Global epidemiology of obesity, vegetarian dietary patterns, and noncommunicable disease in Asian Indians. Am. J. Clin. Nutr. 2014, 100, 359S–364S.
- Rett, B.S.; Whelan, J. Increasing dietary linoleic acid does not increase tissue arachidonic acid content in adults consuming Western-type diets: A systematic review. Nutr. Metab. 2011, 8, 36.
- Yoshida, Y.; Yoshikawa, A.; Kinumi, T.; Ogawa, Y.; Saito, Y.; Ohara, K.; Yamamoto, H.; Imai, Y.; Niki, E. Hydroxyoctadecadienoic acid and oxidatively modified peroxiredoxins in the blood of Alzheimer’s disease patients and their potential as biomarkers. Neurobiol. Aging 2009, 30, 174–185.
- Poirier, B.; Michel, O.; Bazin, R.; Bariéty, J.; Chevalier, J.; Myara, I. Conjugated dienes: A critical trait of lipoprotein oxidizability in renal fibrosis. Nephrol. Dial. Transplant. 2001, 16, 1598–1606.
- Esterbauer, H.; Gebicki, J.; Puhl, H.; Jürgens, G. The role of lipid peroxidation and antioxidants in oxidative modification of LDL. Free Radical Biol. Med. 1992, 13, 341–390.
- Dayton, S.; Hashimoto, S.; Dixon, W.; Pearce, M.L. Composition of lipids in human serum and adipose tissue during prolonged feeding of a diet high in unsaturated fat. J. Lipid. Res. 1966, 7, 103–111.
- Umhau, J.C.; Zhou, W.; Carson, R.E.; Rapoport, S.I.; Polozova, A.; Demar, J.; Hussein, N.; Bhattacharjee, A.K.; Ma, K.; Esposito, G.; et al. Imaging incorporation of circulating docosahexaenoic acid into the human brain using positron emission tomography. J. Lipid. Res. 2009, 50, 1259–1268.
- Braeckman, R.A.; Stirtan, W.; Soni, P.N. Pharmacokinetics of eicosapentaenoic acid in plasma and red blood cells after multiple oral dosing with icosapent ethyl in healthy subjects. Clin. Pharmacol. Drug Dev. 2014, 3, 101–108.
- Bertram, R.; Pedersen, M.G.; Luciani, D.S.; Sherman, A. A simplified model for mitochondrial ATP production. J. Theor. Biol. 2006, 243, 575–586.
- Ahmadpour, S.T.; Maheo, K.; Servais, S.; Brisson, L.; Dumas, J.F. Cardiolipin, the mitochondrial signature lipid: Implication in cancer. Int. J. Mol. Sci. 2020, 21, 8031.
- Dudek, J. Role of cardiolipin in mitochondrial signaling pathways. Front. Cell. Dev. Biol. 2017, 5, 90.
- Marangoni, F.; Agostoni, C.; Borghi, C.; Catapano, A.L. Dietary linoleic acid and human health: Focus on cardiovascular and cardiometabolic effects. Atherosclerosis 2020, 292, 90–98.
- Bu, T.; Tang, D.; Liu, Y.; Chen, D. Trends in Dietary Patterns and Diet-related Behaviors in China. Am. J. Health Behav. 2021, 45, 371–383.
- Zock, P.L.; Katan, M.B. Linoleic acid intake and cancer risk: A review and meta-analysis. Am. J. Clin. Nutr. 1998, 68, 142–153.
- Jaganjac, M.; Zarkovic, N. Lipid Peroxidation Linking Diabetes and Cancer: The Importance of 4-Hydroxynonenal. Antioxid. Redox Signal. 2022, 37, 1222–1233.
- Panov, A.V.; Dikalov, S.I. Cardiolipin, Perhydroxyl Radicals, and Lipid Peroxidation in Mitochondrial Dysfunctions and Aging. Oxidative Med. Cell. Longev. 2020, 2020, 1323028.
- Kim, J.; Minkler, P.E.; Salomon, R.G.; Anderson, V.E.; Hoppel, C.L. Cardiolipin: Characterization of distinct oxidized molecular species. J. Lipid Res. 2011, 52, 125–135.
- Kuschner, C.E.; Choi, J.; Yin, T.; Shinozaki, K.; Becker, L.B.; Lampe, J.W.; Kim, J. Comparing phospholipid profiles of mitochondria and whole tissue: Higher PUFA content in mitochondria is driven by increased phosphatidylcholine unsaturation. J. Chromatogr. B 2018, 1093–1094, 147–157.
- Belury, M.A.; Cole, R.M.; Snoke, D.B.; Banh, T.; Angelotti, A. Linoleic acid, glycemic control and Type 2 diabetes. Prostaglandins Leukot. Essent. Fat. Acids 2018, 132, 30–33.
- Naughton, S.S.; Mathai, M.L.; Hryciw, D.H.; McAinch, A.J. Linoleic acid and the pathogenesis of obesity. Prostaglandins Other Lipid Mediat. 2016, 125, 90–99.
- DiNicolantonio, J.J.; O’Keefe, J.H. Good Fats versus Bad Fats: A Comparison of Fatty Acids in the Promotion of Insulin Resistance, Inflammation, and Obesity. Mo. Med. 2017, 114, 303–307.
- Currenti, W.; Godos, J.; Alanazi, A.M.; Lanza, G.; Ferri, R.; Caraci, F.; Grosso, G.; Galvano, F.; Castellano, S. Dietary Fats and Cognitive Status in Italian Middle-Old Adults. Nutrients 2023, 15, 1429.
- Kim, H.J.; Zhao, J.; Walewski, J.L.; Sparrow, J.R. A High Fat Diet Fosters Elevated Bisretinoids. J. Biol. Chem. 2023, 299, 104784.
- Deol, P.; Kozlova, E.; Valdez, M.; Ho, C.; Yang, E.W.; Richardson, H.; Gonzalez, G.; Truong, E.; Reid, J.; Valdez, J.; et al. Dysregulation of Hypothalamic Gene Expression and the Oxytocinergic System by Soybean Oil Diets in Male Mice. Endocrinology 2020, 161, bqz044.
- Deol, P.; Evans, J.R.; Dhahbi, J.; Chellappa, K.; Han, D.S.; Spindler, S.; Sladek, F.M. Soybean oil is more obesogenic and diabetogenic than coconut oil and fructose in mouse: Potential role for the liver. PLoS ONE 2015, 10, e0132672.
- Deol, P.; Fahrmann, J.; Yang, J.; Evans, J.R.; Rizo, A.; Grapov, D.; Salemi, M.; Wanichthanarak, K.; Fiehn, O.; Phinney, B.; et al. Omega-6 and omega-3 oxylipins are implicated in soybean oil-induced obesity in mice. Sci. Rep. 2017, 7, 12488.
- Piñeros, A.R.; Kulkarni, A.; Gao, H.; Orr, K.S.; Glenn, L.; Huang, F.; Liu, Y.; Gannon, M.; Syed, F.; Wu, W.; et al. Proinflammatory signaling in islet β cells propagates invasion of pathogenic immune cells in autoimmune diabetes. Cell Rep. 2022, 39, 111011.
- Hernandez, M.L.; Sicardo, M.D.; Belaj, A.; Martínez-Rivas, J.M. The oleic/linoleic acid ratio in olive (Olea europaea Lfruit mesocarp is mainly controlled by OeFAD2-2 and OeFAD2-5 genes together with the different specificity of extraplastidial acyltransferase enzymes. Front. Plant Sci. 2021, 12, 653997.
- CBS News. 60 Minutes Overtime: Don’t Fall Victim to Olive Oil Fraud. Available online: https://www.cbsnews.com/news/60-minutes-overtime-how-to-buy-olive-oil/ (accessed on 13 April 2023).
- Shah, A.K.; Dhalla, N.S. Effectiveness of some vitamins in the prevention of cardiovascular disease: A narrative review. Front. Physiol. 2021, 12, 729255.
- Ong, D.E.; Chytil, F. Vitamin A and cancer. Vitam. Horm. 1983, 40, 105–144.
- Sommer, A. Vitamin A deficiency and clinical disease: An historical overview. J. Nutr. 2008, 128, 1835–1839.
- Cabiddu, A.; Delgadilla-Puga, C.; Decandia, M.; Molle, G. Extensive ruminant production systems and milk quality with emphasis on unsaturated fatty acids, volatile compounds, antioxidant protection degree and phenol content. Animals 2019, 9, 771.
- Nürnberg, K.; Wegner, J.; Ender, K. Factors influencing fat composition in muscle and adipose tissue of farm animals. Livest. Prod. Sci. 1998, 56, 145–156.
- Klebanov, G.I.; YuO, T.; Babenkova, I.V.; Lyubitsky, O.B.; OYu, R.; Boldyrev, A.A.; YuA, V. Effect of carnosine and its components on free-radical reactions. Membr. Cell Biol. 1998, 12, 89–99.
- Boldyrev, A.; Bulygina, E.; Leinsoo, T.; Carpenter, D.O. Protection of neuronal cells against reactive oxygen species by carnosine and related compounds. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2004, 137, 81–88.
- Chan, K.M.; Decker, E.A. Endogenous skeletal muscle antioxidants. Crit. Rev. Food Sci. Nutr. 1994, 34, 403–426.