Drug development for Alzheimer’s disease, the leading cause of dementia, has been a long-standing challenge. Saponins, which are steroid or triterpenoid glycosides with various pharmacological activities, have displayed therapeutic potential in treating Alzheimer’s disease. 阿尔茨海默病(痴呆的主要原因)的药物开发一直是一个长期的挑战。皂苷是具有各种药理活性的类固醇或三萜类糖苷,在治疗阿尔茨海默病方面显示出治疗潜力。
- Alzheimer’s disease
- saponin
- amyloid beta
- inflammation
- oxidative stress
- apoptosis
1. Introduction简介
Alzheimer’s disease (AD), the leading cause of dementia, is a progressive neurodegenerative disease [1] that is clinically characterized by memory loss, cognitive impairment, and behavioral disturbances [2]. Since its discovery in 1906, AD has emerged as one of the most costly, fatal, and burdensome diseases of this century [3]. The pathogenesis of AD involves various biological processes [4] involving the abnormal deposition of amyloid beta peptide (Aβ) [5], the accumulation of neurofibrillary tangles (NFTs) [6], neuroinflammation [7], neuronal apoptosis [8], neurotransmitter abnormities [9], and oxidative stress [10]. Despite considerable efforts, drug discovery for the treatment of AD has been slow, with only acetylcholinesterase (AChE)/butyrylcholinesterase (BChE) inhibitors [11] such as galantamine, donepezil, tacrine, and rivastigmine currently available as therapies [12]. However, these treatments only delay the onset of symptoms and cannot halt disease progression and are often associated with significant side effects [13]. Therefore, the development of new therapeutic drugs is urgently needed. Saponins, a type of natural compound, have been extensively studied for their various pharmacological properties [14]. Of particular interest is their potential to enhance learning and memory in individuals with AD [15].
Saponins are naturally occurring compounds that are widely distributed in various plants [16], and they can be divided into two major groups based on their chemical structure: triterpenoid saponins and steroidal saponins [17]. Triterpenoid saponins are further subdivided into tetracyclic triterpenes and pentacyclic triterpenes and are mainly found in plants such as Pentaaceae, Leguminosae, Poria, and Platycodonaceae [18]. The main saponin skeletons of the triterpenoid saponins include dammarane, oleanane, ursane, and lupane (Figure
皂苷是天然存在的化合物,广泛分布于各种植物中[16],根据其化学结构可分为两大类:三萜皂苷和甾体皂苷[17]。三萜皂苷进一步细分为四环三萜和五环三萜,主要存在于五萜科、豆科、茯苓科和鸭嘴科等植物中[18]。三萜皂苷的主要皂苷骨架包括达玛烷、苹果酸、熊烷和鲁帕烷(图 1A–D) [19]. Steroidal saponins, on the other hand, are mainly found in plants such as Dioscoreaceae, Liliaceae, and Scrophulariaceae [20]. The main saponin metaskeletons of steroidal saponins include spirostane, furostane, cholestane, and cardenolide (Figure
A-D)[19]。另一方面,甾体皂苷主要存在于薯蓣科、百合科和蝙蝠科等植物中[20]。甾体皂苷的主要皂苷代谢物包括螺糠烷、呋喃烷、胆甾烷和卡氏内酯(图 1E–H) [21]. Saponins possess multiple bioactivities, such as reduction of amyloid beta (Aβ) deposition [22], inhibition of tau protein phosphorylation [23], antioxidation [24], antiapoptosis [25], and anti-inflammation [26]. These properties make saponins promising therapeutic candidates for AD and other neurological disorders [27]. Meanwhile, the diversity of saponins found in different plants [28] makes them a valuable source of potential drugs for the treatment of AD.
E-H)[21]。皂苷具有多种生物活性,如减少β淀粉样蛋白沉积[22]、抑制tau蛋白磷酸化[23]、抗氧化[24]、抗细胞凋亡[25]和抗炎[26]。这些特性使皂苷成为AD和其他神经系统疾病的候选药物[27]。同时,在不同植物中发现的皂苷的多样性[28]使它们成为治疗AD的潜在药物的宝贵来源。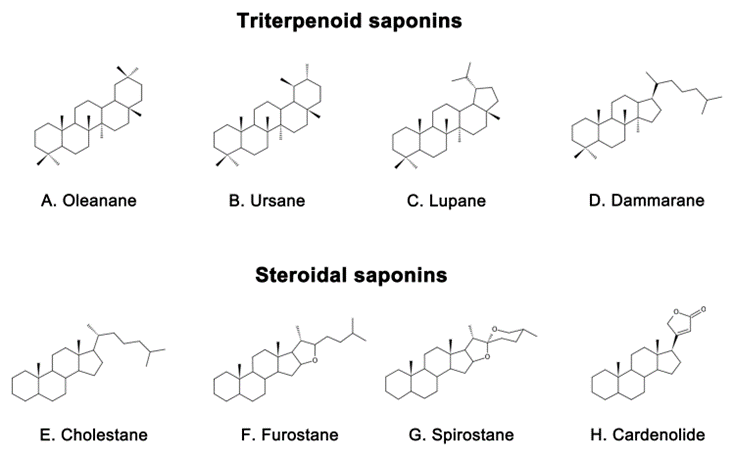
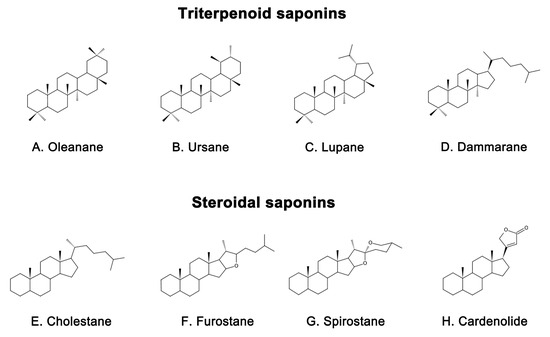
Figure 图1. Representative triterpenoid saponins include (
A) Oleanane, (
B) Ursane, (
C) Lupane, (
D) Dammarane and steroidal saponins include (
E) Cholestane, (
F) Furostane, (
G) Spirostane, (
H) Cardenolide.
2. Mechanism of Saponins in Treating Alzheimer’s Disease皂苷治疗阿尔茨海默病的机制
2.1. Inhibition of Aβ Deposition and Neurotoxicity
2.1. 抑制Aβ沉积和神经毒性
2.2. Inhibiting Aberrant Tau Protein Phosphorylation
2.2. 抑制异常tau蛋白磷酸化
NFTs, a well-known pathological hallmark of 是 AD [78],的一个众所周知的病理标志 are[104],由称为微管蛋白相关单元 composed of excessively phosphorylated proteins linked by helical filaments known as tubulin-associated unit (tau) protein [79]. (tau) 蛋白的螺旋丝连接的过度磷酸化蛋白质组成 [105]。Tau pro蛋白主要存在于神经元轴突中,是一种微管相关蛋白,参与多种功能,如轴突微管维持、细胞质转运和神经元信号传导调节[106,107]。其最佳功能取决于正常磷酸化的低水平[108]。然而,tein, mainly found in neuronal axons, is a microtubule-associated protein involved in various functions such as axonal microtubule maintenance, cytoplasmic transport, and regulation of neuronal signaling [80][81]. Its optimal function depends on the low levels of normal phosphorylation [82]. However, overphosphorylation of tau protein leads to its detachment from microtubules, resulting in microtubule depolymerization, aggregation into u蛋白的过度磷酸化导致其与微管分离,导致微管解聚,聚集成NFTs, and ultimately neurodegeneration and cell death [83]. The correlation between ,最终导致神经变性和细胞死亡[109]。NFT densi密度与认知能力下降之间的相关性表明ty and cognitive decline suggests that tau protein plays a central role in AD [84]. Furau蛋白在AD中起核心作用[110]。此外,thermore, the prevalence of tau protein pathology positively correlates with cognitive impairment in AD, where a person with AD has increased levels of tau protein, hyperphosphorylated tau protein, and aggregated tau in u蛋白病理学的患病率与AD患者的认知障碍呈正相关,AD患者的大脑中tau蛋白、过度磷酸化tau蛋白和聚集tau蛋白水平升高[111]。异常their brain [85]. Abnormal tau protein phosphorylation plays a greater role in 蛋白磷酸化在AD than the total tau中的作用大于总tau蛋白[112]。因此,它被认为是 protein [86]. As such, it is considered a critical pathogenic step in the development of NFTs and, subsequently, 以及随后 AD [87].发展的关键致病步骤[113]。 Several saponins have been found 已经发现几种皂苷可以抑制to inhibit the hyperphosphorylation of tau proteins, protect synaptic structures, and reduce neuronal damage. An octillol-type saponin, PF11, has been found to increase the activity of phosphatase-2A (PP2A) by enhancing leucine carboxyl methyltransferase-1 (LCMT-1) and improving cognitive impairment in SAMP/8 mice by inhibiting the hyperphosphorylation of tau protein at serine 396 and tyrosine 205 in the brain [58]. au蛋白的过度磷酸化,保护突触结构,并减少神经元损伤。研究发现,辛替洛尔型皂苷PF11通过增强亮氨酸羧基甲基转移酶-2(LCMT-2)来增加磷酸酶-1A(PP1A)的活性,并通过抑制大脑中丝氨酸8和酪氨酸396处tau蛋白的过度磷酸化来改善SAMP/205小鼠的认知障碍[63]。链脲佐菌素(Streptozotocin (STZ) is a glucosamine–nitrosourea compound commonly used in AD studies to induce tau proteins’ hyperphosphorylation and cognitive impairment [88]. )是一种氨基葡萄糖-亚硝基脲化合物,常用于AD研究,以诱导tau蛋白的过度磷酸化和认知障碍[114]。Tau protein phosphorylation occurs at multiple sites, usually associated with glycogen synthase kinase (蛋白磷酸化发生在多个位点,通常与糖原合酶激酶(GSK) 3 and cyclin-dependent kinase 5 (CDK5). PF11 was also shown to attenuate STZ-induced tau hyperphosphorylation in the brain of male Wistar rats by correcting the dysregulation of the insulin receptor substrate )3和细胞周期蛋白依赖性激酶5(CDK5)相关。PF11还通过纠正胰岛素受体底物1/PI3K/Akt/GSK-3β and calpain 和钙蛋白酶I/CDK5 signaling pathways [65]. Addi信号通路的失调,减弱STZ诱导的雄性Wistar大鼠大脑中的tionally, u过度磷酸化[70]。此外,PF11 ameliorated 改善了STZ-induced learning and memory deficits, synaptic damage, and neuronal death [65]. Another active compound, Ginsenoside 诱导的学习和记忆缺陷、突触损伤和神经元死亡[70]。在人参中发现的另一种活性化合物人参皂苷Rb1[11, found in ginseng [89], exhibits neuroprotective properties against different neurotoxins [90]. 5]对不同的神经毒素具有神经保护特性[116]。研究表明,人参皂苷Research has shown that ginsenoside Rb1 counters cognitive impairment induced by aluminum oxide in ICR mice while reducing Ser396 tau phosphorylation by increasing PP2A levels and decreasing p-GSK levels [85][91]. Ginsenoside Rd, anothe可对抗ICR小鼠氧化铝诱导的认知障碍,同时通过增加PP396A水平和降低p-GSK水平来减少Ser compound found in ginseng [92], has been shown 2to have potential therapeutic applications in neurou磷酸化[111,117]。人参中发现的另一种化合物人参皂苷Rdegenerative diseases [93]. It has been demonstrated to reduce tau proteins’ hyperphosphorylation and neurotoxicity induced by okadaic acid (O[118]已被证明在神经退行性疾病中具有潜在的治疗应用[119]。在A) by increasing D的大鼠和皮质神经元模型中,已经证明可以通过增加PP2A activity in both rat and cortical neuronal models of AD [94].活性来减少冈田酸(OA)诱导的tau蛋白过度磷酸化和神经毒性[120]。 Theasaponin 茶皂苷E1, a vital compound found in green tea seeds, shows remarkable potential in reducing p-tau protein levels and inhibiting AD-promoting genes while activating AD-remitting genes [95]. This potential has been observed in AD models constructed from 是一种在绿茶种子中发现的重要化合物,在降低p-tau蛋白水平和抑制AD促进基因方面显示出显着的潜力,同时激活AD缓解基因[121]。在由SHY5Y and glioblastoma (和胶质母细胞瘤(HTB2) cells. Notably, theasaponin E1 successfully reduced p-tau protein by inhibiting several enzymes, including )细胞构建的AD模型中已经观察到这种潜力。值得注意的是,茶皂苷E1通过抑制几种酶(包括GSK-3β, CDK5, 、CDK5、c-Jun NH2-terminal kinase (JNK), MAPK, 末端激酶(JNK)、MAPK、ERK1/MARK, and calmodulin-dependent protein kinase II alpha [95]. Similarly, Xan和钙调蛋白依赖性蛋白激酶IIΑ)成功还原p-thoceraside, a triterpene saponin extracted from the shell of Xanthoceras sorbifolia Bunge, has been shown u蛋白[121]。同样,黄花芍是一种从山梨黄芪壳中提取的三萜皂苷,已被证明通过抑制to exert neuroprotective effects by inhibiting the phosphorylation of tau protein [96]. In a study conducted on mice with 蛋白的磷酸化来发挥神经保护作用[122]。在一项针对AD, the administration of Xanthoceraside significantly decreased the ex小鼠的研究中,给予黄尾花苷显著降低了pression of pGSK-3β and tau protein at 和tau蛋白在Ser396 and 和Ser404, thereby effectively inhibiting the phosphorylation of tau protein [54].处的表达,从而有效抑制了tau蛋白的磷酸化[59]。2.3. 抗炎作用
2.3. Anti-Inflammatory Effect
In recent years, neuroinflammation has become recognized as a third neuropathological hallmark of 近年来,神经炎症已成为除AD, in addition to Aβ and NFTs [97]. Neuroinflammation is an inflammatory response to pathological insults that occur within the central nervous system (CNS), such as infection, trauma, ischemia, and toxin accumulation [98]. This response is characterized by the proliferation and activation of microglia and astrocytes in the brain [99]. Such activation is usually accompanied by the secretion of inflammatory cytokines such as interleukin-1β (和NFTs之外AD的第三个神经病理学标志[123]。神经炎症是对中枢神经系统内发生的病理性损伤(如感染、创伤、缺血和毒素积累)的炎症反应[124]。这种反应的特征是大脑中小胶质细胞和星形胶质细胞的增殖和活化[125]。这种激活通常伴有炎症细胞因子的分泌,例如IL-1β), interleukin 6 (、白细胞介素1(IL-6), and tumor necrosis factor α (TNF-α) [100]. Microglia are innate immune cells that reside within the )和肿瘤坏死因子α(TNF-α)[6]。小胶质细胞是存在于CNS and are thought to be important regulators in the inflammatory response within the brain [101]. In the early stages of 内的先天免疫细胞,被认为是大脑内炎症反应的重要调节因子[126]。在AD, activated microglia can exert protective effects by phagocytosing and degrading 的早期阶段,活化的小胶质细胞可以通过吞噬和降解Aβ [102][103]. However, when 发挥保护作用[127,128]。然而,当Aβ accumulates excessively, it binds various receptors on the surface of microglia such as cluster of differentiation (过度积累时,它会结合小胶质细胞表面的各种受体,如分化簇(CD) 14, CD36 [104], )129、CD14[36]、Toll-like receptor (样受体(TLR) 4, and TLR2, to promote the release of inflammatory factors [105]. This response leads to neuroinflammation and further impairment of cognitive function. Similarly, astrocyte activation is also associated with )130和TLR4,促进炎症因子的释放[2]。这种反应导致神经炎症和认知功能的进一步损害。同样,星形胶质细胞活化也与Aβ [106]. 有关[131]。活化的星形胶质细胞产生各种炎症介质[132]并分泌少量Activated astrocytes produce various inflammatory mediators [107] and secrete small amounts of Aβ [108], resulting in a chronic vicious cycle [107]. These reactions can exacerbate neuroinflammation and damage neurons.[133],导致慢性恶性循环[134]。这些反应会加剧神经炎症并损害神经元。 Multiple pathways are involved in microglia小胶质细胞和星形胶质细胞介导的神经炎症涉及多种途径[135],其中一条是核因子κ- and astrocyte-mediated neuroinflammation [109], one of which is the nuclear factor kappa-B ((NF-κB) pathway [110]. )途径[136]。该途径激活This pathway activates the transcription of TNF-α, IL-1β, and IL-6, which exacerbates inflammation [111]. Microglial activation also depends on other signaling pathways such as the 、IL-1β和IL-6的转录,从而加剧炎症[137]。小胶质细胞激活还取决于其他信号通路,例如PI3K/Akt/mTOR pathway [112] which can promote the expression of the inflammatory mediators 通路[138],其可促进炎症介质NO synthase (合酶(NOS) and cyclooxygenase-2 (Cox-2) [113][114]. Furthermore, )和环氧合酶-2(Cox-2)的表达[139,140]。此外,MAPK/p38 [115], [141]、Janus kinase (JAK)/signal transducer, and activator of transcription (STAT) and other pathways also contribute to neuroinflammation [116]. 激酶/信号转导器、转录激活剂(STAT)和其他通路也有助于神经炎症[142]。因此,调节炎症信号通路和抑制促炎因子的产生可能为治疗AD提供一种有希望的方法。 一般来说,天然皂苷具有抗炎生理活性,从而通过调节炎症相关的信号通路来抑制炎症因子的表达或释放来保护神经细胞。2.4. 改善线粒体功能和抗氧化应激
线粒体作为有氧代谢的必需细胞器,可显著促进氧化应激的产生[180]。同时,现在已经表明,线粒体氧化应激可导致轴突和神经元损伤,并最终导致神经功能障碍[181]。当线粒体内发生氧化应激时,可导致线粒体肿胀、外膜破裂和钙离子丢失,导致ROS的进一步产生和释放[182]。在生理条件下,ROS可以刺激细胞生长[183],并被抗氧化酶和其他抗氧化防御系统[184]消除,如超氧化物歧化酶(SOD)、谷胱甘肽过氧化物酶(GSH-Px)和过氧化氢酶(CATherefore, modula)[185]。然而,当发生氧化还原失衡状态时,自由基大量积聚,导致脂质过氧化,导致丙二醛(MDA)的产生[186]、细胞膜损伤、线粒体膜损伤,甚至神经元死亡[187]。此外,氧化应激可能与大脑中的Aβ聚集[188]、ting inflammatory signaling pathways and inhibiting the production of proinflammatory factors may offer a promising approach to the treatment of u蛋白磷酸化[189]和金属离子微环境损伤相互作用,放大神经元损伤,最终导致AD.的发病机制[190,191]。2.4. Improvement in Mitochondrial Function and Antioxidative Stress
Oxidative stress refers to an imbalance between oxidation and antioxidation in the body [117], which is mainly characterized by the increased production of reactive oxygen species and reduced ability of antioxidants to combat them [118]. The brain is susceptible to oxidative damage due to its high lipid content and lacks effective antioxidant defense mechanisms, despite high oxygen consumption [119][120]. It is now well established that the level of oxidation in the brain also increases with age [121]. Interestingly, extensive oxidative damage can be observed in the stage of mild cognitive impairment that precedes the typical clinical manifestations of AD, suggesting that oxidative stress may be a central mechanism in driving the disease [122].
Several natural saponins may act as exogenous antioxidants to combat AD by managing oxidative stress [123][124][192,193]. Ginsenoside Rh2, a rare ginsenoside with few reports on its neuroprotective effects compared to other similar molecules [125][194], has shown significant neuroprotective effects in a scopolamine-induced model of memory impairment in ICR mice. Ginsenoside Rh2 effectively managed MDA levels while increasing glutathione (GSH) levels and SOD activity in the brain, effectively inhibiting oxidative stress [126][195]. These findings suggest that ginsenoside Rh2 has great potential for the treatment of AD by inhibiting oxidative stress in the brain. Crude saponins from BF showed potent antioxidant effects in addition to anti-inflammatory effects. In addition, saponins from BF alleviated oxidative stress, reduced ROS, and MDA levels by restoring GSH-Px, GSH, and CAT levels in the Aβ ICV-induced BALB/c AD mouse model [127][161]. Similarly, ginsenoside Rd was found to reduce protein carbonyls and 4-hydroxy-2-nonenal (4-HNE) levels by decreasing the glutathione disulfide (GSSG)/GSH ratio, thereby counteracting oxidative stress in the brain of Aβ-induced AD mouse models [128][170]. Ginsenoside Rg3, a tetracyclic triterpenoid saponin, is the major bioactive component of ginseng and is widely used for the treatment of tumors [129][196]. Results from in vivo studies have shown that ginsenoside Rg3 can improve mitochondrial dysfunction and enhance ROS scavenging ability by increasing SOD, CAT, and GSH-Px levels, thus ameliorating oxidative stress in the brain of D-gal-induced AD rats [130][197]. PF11 was found to inhibit the production of both APP and Aβ in the cortex and hippocampus, while restoring the activity of SOD and GSH-Px in the brains of AD mice and reducing MDA levels in the cortex [131][198]. Ginsenoside Rg1 has also been found to protect primary cortical neurons against Aβ-induced increases in ROS and H2O2 levels by inhibiting oxidative stress [132][199]. This results in a decrease in mitochondrial membrane potential and an increase in cytochrome C (Cyt-c) oxidase activity, ultimately leading to a decrease in Cyt-c release and indicating improved mitochondrial function [132][199]. Another newly discovered saponin compound, huangjingsterol B from Polygonatum cyrtonema Hua, showed significant antioxidant activity [133][200] by reducing oxidative stress and protecting cells from Aβ-induced damage through modulating the activity of GSH-Px and SOD [133][200]. Similarly, researchers have found that a saponin from the fruit of Solanum anguivi can protect the P2 region of rat brain synaptosomes from oxidative damage induced by ferrous iron and sodium nitroprusside, reduce ROS generation, restore total thiol levels, and protect mitochondrial function [123][192]. This suggests that saponins from Solanum anguivi fruits may have antioxidant properties that benefit mitochondrial function [134][201]. PGS has a similar effect with its ability to upregulate intracellular heme oxygenase-1 (HO-1), SOD, CAT, and GSH-Px to inhibit Aβ-induced ROS production and reduce oxidative damage in AD brains [135][153]. Despite the demonstrated neuroprotective effects of Panax ginseng, the mechanisms underlying its antioxidant properties remain poorly understood. However, a recent study has shown that the primary bioactive compound in Panax ginseng, NTR1, is able to prevent the loss of mitochondrial membrane potential and reduce the accumulation of ROS induced by Aβ [136][202]. This suggests that NTR1 may exert neuroprotective effects by inhibiting oxidative stress. Furthermore, most previous experiments have focused on saponins extracted from Panax ginseng roots, and the pharmacological effects of other parts, such as saponins in P. notoginseng leaves (LPNS), are largely unknown [137][203]. LPNS, which mainly include the ginsenosides Rb1, Rb2, Rb3, and Rc, have been shown to have potent neuroprotective properties [137][203]. In H2O2- or oxygen and glucose deprivation (OGD)-induced astrocytes from the cortex of SD rats and SH-SY5Y cells, LPNS increased the levels of nuclear factor erythroid2-related factor 2 (Nrf2) and its downstream HO-1 and glutathione S-transferase pi 1 (GSTP1) in response to increased ROS, as well as significantly reducing lactate dehydrogenase (LDH) expression and increasing the cellular activity of astrocytes [137][203].