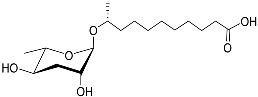
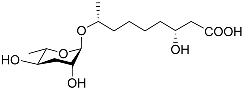
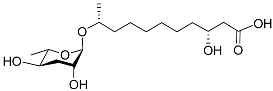
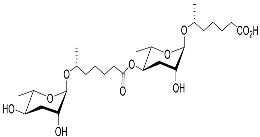
1. Development-Related Pheromones Secreted by Nematodes
The ability of nematodes to be so widely distributed in nature is closely related to their special developmental patterns.
C. elegans lives freely in soil; it has a small, transparent body and serves as a model nematode species
[1][39]. When the environment is suitable, an individual
C. elegans starts to develop from a fertilized egg and progresses to adulthood through four stages of development. However, it stops feeding and developing if it encounters extreme conditions, such as a lack of food, elevated temperatures, or an increase in population density, and then the larva may enter a highly stress-resistant state called dauer diapause. This stage can last for several months. The nematodes eventually resume development and molt into the reproductive cycle under suitable conditions
[2][3][4][5][6][40,41,42,43,44]. Much research shows that chemical pheromones can control dauer entry and exit
[7][8][45,46].
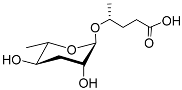
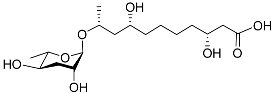
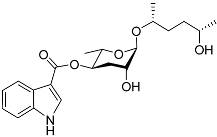
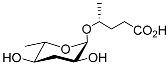
The first dauer-inducing pheromone (daumone) was identified by means of the ethyl acetate extraction of a
C. elegans liquid medium. The molecular structure of daumone was thereby determined to be (2)-(6
R)-(3,5-dihydroxy-6-methyltetrahydropyran-2-yloxy) heptanoic acid, abbreviated to ascr#1 (also called daumone-1/ascaroside C7/asc-C7) (
Table 1)
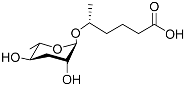
[9][47]. Subsequently, ascr#2 (also called daumone-2/ascaroside C6) (
Table 1) and ascr#3 (also called daumone-3/asc-∆C9/ascaroside C9) (
Table 1) were also isolated and identified. Ascr#2 and ascr#3 induce dauer formation about 100 times more potently than ascr#1 does
[10][48]. The pheromones that induce dauer formation may be single ascarosides or mixtures of different ascarosides, and these pheromones often act synergistically when mixed together. The dauer pheromone of
C. elegans is mainly composed of ascr#2, ascr#3, and several other components, while in
Caenorhabditis briggsae, the main component of the dauer pheromone is ascr#2
[11][49]. A derivative of ascr#2 has a β-glucosyl substituent linked to C2 of the ascarylose in ascr#2 and is named ascr#4 (also called daumone-4) (
Table 1). The activity of ascr#4 is low
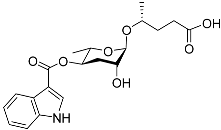
[12][50]. Ascr#5 (also called daumone-5/ascaroside C3/asc-ωC3) (
Table 1) is a potent dauer pheromone. The main function of ascr#5 is the activation of the axon regeneration pathway via SRG-36/SRG-37 GPCRs and EGL-30, indicating ascaroside signaling promotes axon regeneration by activating the GPCR-Gqα pathway
[13][25]. In addition, the
crh-1 gene plays an autonomous role in ascr#5, sensing ASI neurons in order to inhibit the dauer formation of
C. elegans L2d
[14][26]. Ascr#5 also produces synergistic effects with ascr#2 and ascr#3
[15][51]. In ASI neurons, ascaroside pheromones (compounds composed of ascr#2, ascr#3, and ascr#5) reversibly inhibit the expression of the
str-3 chemical receptor gene, and when ascarosides are removed, its expression resumes. This process mainly occurs through the GPCR receptors SRBC-64/66 and SRG-36/37, which are required for
str-3 repression
[16][35]. Ascr#8 (
Table 1) uniquely possesses a p-aminobenzoate group in its terminus; this group is a folate precursor that is derived from bacteria and is not synthesized by
C. elegans [17][52].
P. pacificus is a model species that has been extensively studied in biology
[18][55]. This nematode can enter the dauer stage or other stages if food is enough for growth
[19][20][56,57]. Typically, the mouth of an adult that preys on other nematodes is more complex than that of a bacterivorous nematode. Pheromones can regulate the mouth dimorphism of
P. pacificus [21][58]. Neelanjan et al.
[22][59] analyzed fractions of the
P. pacificus exo-metabolome and found that it has rich signaling molecules controlling adult phenotypic plasticity, including ascarosides ascr#1, 9, and 12. Pasc#9 (
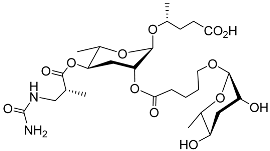
Table 1) was the most abundant derivative after pasc#1 and pasc#12. Pasc#9 comprises an N-succinyl 1-phenylethanolamide connected to ascarylose with a 4-hydroxypentanoic acid chain. Dasc#1 (
Table 1) consists of two ascr#1 units; one ascr#1 unit is connected to carbon 4 of the other ascr#1 unit. A 3-ureido isobutyrate moiety is also present on carbon 4 of ubas#1 (
Table 1), and ubas#1 also contains ascr#9 with the (ω)-oxygenated ascaroside oscr#9 connected at position 2
[22][59]. Recent studies revealed that the formation of ubas#1 and related metabolites specifically requires the putative carboxylesterase
Ppa-uar-1 [23][60]. Additionally, dimeric ascarosides and ureido isobutyrate-substituted metabolites were first reported in
P. pacificus. L-paratose forms the basis of part#9 (
Table 1). Part#9 only differs from ascr#9 in terms of the stereochemistry of one hydroxyl group. Part#9 is also one of the components of npar#1. Npar#1 (
Table 1) contains a derivative of the nucleoside adenosine. Although part#9, npar#1, ubas#1, and pasc#9 can induce dauer formation, npar#1 has a more intense effect than the others. Pasc#9, ascr#1, dasc#1, and npar#1 can induce eurystomatous mouth formation, a predatory morphology in the final larval and juvenile stages, in which
P. pacificus-specific dasc#1 plays an important role
[24][25][61,62].
Heterorhabditis bacteriophora is parasitic toward insects and has a developmental process similar to that of
C. elegans. In the soil, the infective juveniles (IJs) survive as the only state of entomopathogenic nematodes. After IJs infect the host insects, they recover and lay eggs in their adults, which develop through four larval stages (J1–J4) to form the next generation
[26][27][28][63,64,65]. In this process,
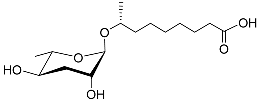
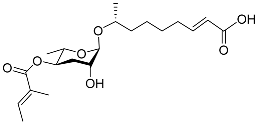
H. bacteriophora secretes the ascaroside C11 ethanolamine (asc C11 EA) (
Table 1), which prevents IJs from recovering to the J4 stage. Asc C11 EA comprises an ascarylose sugar, an ethanolamine fragment, and a carbon side chain containing ω-1 alcohol; the fatty-acid-derived portion of the side chain is 11 carbons long. Asc C11 EA and the dauer pheromone of
C. elegans show structural similarity
[29][66].
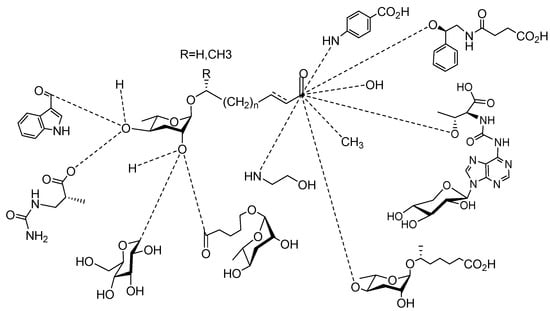
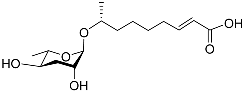
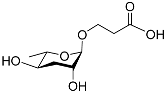
Figure 1 illustrates the schematic structure of the pheromones secreted by nematodes during their development. The figure presents a schematic diagram showing ascarylose sugars, variable-length fatty acids, and other moieties modifying them, which form different species of development-related ascaroside species.
Figure 1. Overview of the chemical structure of development-related pheromones secreted by nematodes. Nacq#1 does not have an ascaroside structure, so it is not shown here, and its detailed structure is shown in Table 1.
2. Sex Pheromones Secreted by Nematodes
Mate selection is universal in sexually reproducing organisms, and pheromones provide individuals with advantageous mating information that helps them to select high-quality mates. In the twentieth century, the first sex pheromone was named bombykol, which is released from female silk moths (
Bombyx mori)
[30][67]. Sex pheromones have since been researched in more depth; they are defined as chemical substances produced by individuals that cause innate and rigid sexual behavior
[31][68]. These pheromones have both sex- and species-specific effects. Nematode mating behavior is also regulated by pheromones
[32][69]. Generally, the nematode mating response can be induced when the pheromone concentration is much lower than the concentration required for dauer formation.
C. elegans mainly reproduces as a hermaphrodite. However, most
Caenorhabditis worm species achieve this by means of cross-fertilization. These hermaphrodites are essentially females with the ability to self-fertilize, and they can also mate with males, but their numbers are typically relatively low. Hermaphrodites do not appear to be attracted to male
C. elegans, but males are attracted to them
[33][70]. The ascarosides ascr#2, 3, 4, and 8 (
Table 1) not only play roles in regulating nematode development but also function as sex pheromones that are known to attract males
[34][35][71,72]. They show synergistic effects, whereby a mixture of ascr#2, 3, and 4 is an effective male attractant at low concentrations. Ascr#3 attracts
C. elegans males but repels hermaphrodites and can increase the lifespan of
C. elegans. Ascr#8 is a strong male-specific attractant and shows synergy with ascr#2 and ascr#3. A mixture composed of ascr#3 and ascr#8 strongly attracts males at ultra-low concentrations, but at higher concentrations, it is repulsive to hermaphrodites
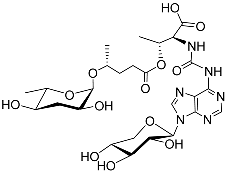
[12][34][36][36,50,71]. The other two ascarosides with sex pheromone functions, ascr#6.1 (
Table 1) and ascr#6.2 (
Table 1), were identified by Paul as diastereomeric side-chain-hydroxylated ascarosides
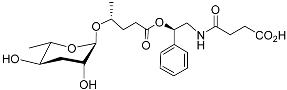
[34][71]. Ascr#10 (also called asc-C9) (
Table 1) makes up the majority of the sex-specific milieu of ascarosides produced by male
C. elegans. Ascr#3 has an α, β-unsaturated fatty acid moiety, whereas ascr#10 has the corresponding dihydro-derivative; such minor structural modifications deeply influence their signaling properties. The male pheromone ascr#10 strongly attracts hermaphrodite nematodes and shortens their lifespan
[37][38][39][73,74,75]. It also can increase germline proliferation and physiological cell death
[40][76] and change the reproductive physiology of hermaphroditism, such as by improving sperm orientation and increasing the number of reproductive precursor cells in adults
[41][42][43][77,78,79]. Furthermore, Dong et al. conducted a comparative analysis of indole ascaroside signaling for 14
Caenorhabditis species. Icas#2 and icas#6.2 (
Table 1) were isolated from hermaphrodites of
C. briggsae and were found to synergistically attract conspecific males
[44][80].
Panagrellus redivivus has an ecological niche similar to that of
C. elegans; it has a free-living lifestyle but belongs to a different clade. In contrast with
C. elegans, the virgin females of
P. redivivus attract and are attracted by the males, but they do not attract the same sex
[45][81]. The ascaroside biosynthesis in
P. redivivus is highly sex-specific. The females of
P. redivivus can excrete ascr#1, ascr#10, and bhas#10 (
Table 1)
[38][74]. The males of
P. redivivus can excrete dhas#18 (
Table 1)
[38][74]. Ascr#1 can strongly attract males, but high concentrations of ascr#1 repel the females of
P. redivivus. At high concentrations, bhas#10 and ascr#10 attract males rather than females. Dhas#18, which is a known dihydroxy derivative of ascr#18 secreted by males as well as an ascaroside with extensive functionality as a characteristic of its lipid-derived side chain, can strongly attract the females of
P. redivivus. Bhas#18 (
Table 1) is a precursor for dhas#18 synthesis, but its exact function is unclear
[38][74].
Rhabditis sp. SB347 is a unique free-living dioecious species that is often used in the laboratory
[46][82]. The females of SB347 produce ascr#1 and ascr#9 (
Table 1), which function as sex pheromones. At femtomolar levels, ascr#1 and ascr#9 are strongly attracted to males, but not to hermaphrodites and female nematodes
[47][83].
The dimorphism of the adults is an important feature of the life history of
Globodera rostochiensis. Hermaphrodites attract males for mating by producing pheromones. Four fractions of the homospecific sex pheromone produced by virgin females, which were isolated using chromatography technology, were tested for their ability to attract male
G. rostochiensis; only two of the fractions showed sex pheromone activity. Several weakly basic polar compounds constitute the sex pheromone of
G. rostochiensis. The exact structure of its components is unclear
[48][88].
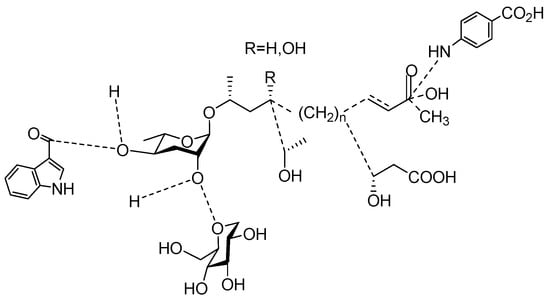
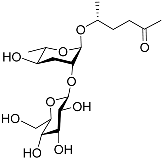
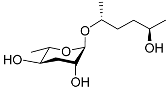
The chemical structure diagram for sex pheromones secreted by nematodes is shown in Figure 2. This diagram shows ascaroside building blocks associated with the mating of different species of nematodes, including ascarylose sugars, variable-length fatty acids, and other modification groups.
Figure 2. Overview of the chemical structures of sex pheromones secreted by nematodes. Vanillic acid does not have an ascaroside structure, so it is not shown here, and its detailed structure is presented in Table 1.
3. Aggregation of Pheromones Secreted by Nematodes
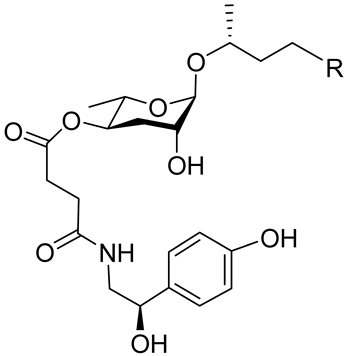
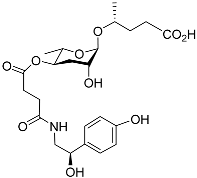
C. elegans uses specifically modified forms of the ascarosides that contain indole units as highly effective aggregation pheromones. The indole ascarosides (ICASs) incorporate an L-tryptophan-derived indole-3-carboxylic acid group, which is linked to the four-position of the ascarylose moiety. An indole carboxy unit forms one indole derivative, and it is connected to an ascarylose bearing a nine-carbon unsaturated side chain identical to that found in the known ascr#3; this indole carboxy ascaroside is called “icas#3” (
Table 1). The icas#3 occurs primarily by means of an expression protein in
C. elegans, CEST-3, adding an IC group to the corresponding unmodified ascr#3
[49][50][15,89]. Icas#3 and icas#9 are relatively good attractants
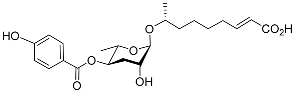
[49][15]. The 4-hydroxybenzoyl derivative of ascr#3 is called hbas#3 (
Table 1). Hbas#3 was the first ascaroside with a 4-hydroxybenzoyl structure to be discovered. Hbas#3 strongly attracts
C. elegans at low concentrations (10 fM), more effectively so than icas#3 and icas#9
[51][19]. Ascr#5 in combination with ascr#2 or ascr#3 may influence the aggregation of
C. elegans adults; however, more in-depth research is needed on this topic
[52][90].
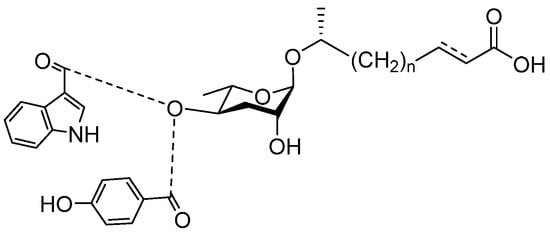
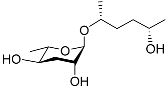
See Figure 3 for a schematic overview of nematode pheromones related to their aggregation. It illustrates ascarylose sugars, fatty acids with variable lengths, and other modifications that form aggregation-related ascaroside species.
Figure 3.
Overview of the chemical structure of the aggregation of pheromones secreted by nematodes.
4. Pheromones with Other Functions Secreted by Nematodes
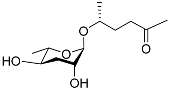
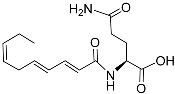
The L1 larvae of
C. elegans can specifically produce certain octopamine ascarosides, in which the ascarylose four-position is linked to a side chain derived from the succinylation of the neurotransmitter octopamine. The octopamine ascarosides osas#2 (
Table 1), osas#9 (
Table 1), and osas#10 (
Table 1) play roles in dispersal
[35][72]. Osas#9 is a pheromone that acts as a dispersal signal, especially in the case of a lack of food. Avoidance reactions to osas#9 require the G-protein-coupled receptor TYRA-2
[53][34]. With a continuous decrease in food, ascr#10 and osas#10 are converted to ascr#9, osas#9, and icas#9
[35][72].
Ascr#3 was found to regulate metabolism and avoidance behavior, it was defined as a population density pheromone. When food is scarce, ascr#3 causes hermaphrodites to have an avoidance effect
[54][55][56][91,92,93]. When the ADF of a single sensory neuron is removed, both sexes are weakly rejected by the ascaroside ascr#3. Although ADF has functions in both sexes, ascr#3 is only detected in males, which is the result of the main sex regulator
tra-1 [57][94]. A derivative of ascr#3 called mbas#3 (
Table 1) was the first ascaroside discovered to have an (E)-2-methyl-2-butenoyl structure
[51][19], and it acts as a dispersal signal in
C. elegans, as well as having an antagonistic effect on the attractant characteristics of indole ascarides such as icas#3 and icas#9
[58][95].
C. elegans has shown a tendency to be attracted to a series of odorous substances, and with the passage of time, this tendency changes from attraction to dispersal
[59][60][98,99]. This varied pheromone-mediated behavior is called olfactory plasticity, which depends on the population density
[61][110]. However, the pheromone component that plays a major role in this process has not been identified. In addition, the pheromones released by injured conspecific nematodes are repellent to nematodes, and they may contain alarm pheromones. These alarm pheromones may not belong to the ascaroside class of pheromones
[62][111]. However, their exact structure has not been identified.
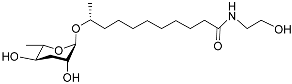
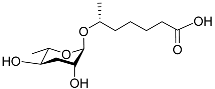
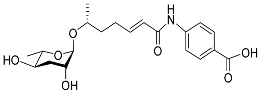
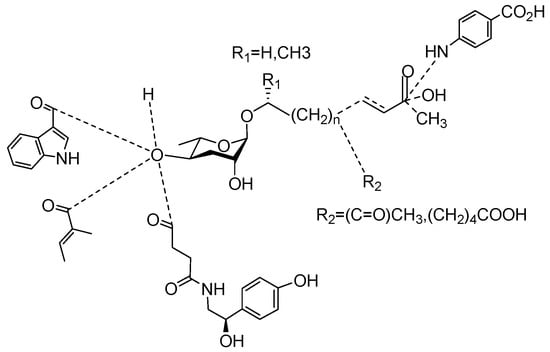
The following Figure 4 displays the chemical structure diagram of function pheromones secreted by nematodes. This diagram shows the ascaroside building blocks associated with the functions of different species of nematodes, including ascarylose sugars, variable-length fatty acids, and other modification groups.
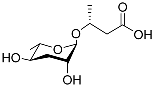
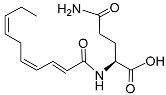
Table 1.
Pheromones are secreted by nematodes.
Figure 4.
Overview of the chemical structures of pheromones with other functions secreted by nematodes.