Autophagy is a “self-eating” process that engulfs cellular contents for their subsequent digestion in lysosomes to engage the metabolic need in response to starvation or environmental insults. According to the contents of degradation, autophagy can be divided into bulk autophagy (non-selective autophagy) and selective autophagy. Bulk autophagy degrades non-specific cytoplasmic materials in response to nutrient starvation while selective autophagy targets specific cargoes, such as damaged organelles, protein aggregates, and intracellular pathogens. Selective autophagy has been documented to relate to the reproductive processes, especially for the spermatogenesis, fertilization, and biosynthesis of testosterone.
- autophagy
- selective autophagy
- spermatogenesis
- male fertility
Note:Dear author, the following contents are excerpts from your papers. They are editable. And the entry will be online only after authors edit and submit it.
1. Introduction
Spermatogenesis is a complex biological process of germ cell proliferation and differentiation that produces a large number of spermatozoa in the seminiferous tubules. It contains three processes: mitosis of spermatogonia, meiosis of spermatocytes, and spermiogenesis, by which round spermatids transform to become the elongated spermatids [1].[1] Successful spermatogenesis, which progresses through precisely timed and highly organized cycles, is crucial to produce spermatozoa continuously and maintain adult male fertility. In contrast, abnormal spermatogenesis usually results in male subfertility or infertility [2]. The fine communication between germ cells and somatic cells within seminiferous tubules is fundamental to normal spermatogenesis. Among them, the support and nutritional function of Sertoli cells and endocrine function of Leydig cells are particularly important [3,4][3][4].
Autophagy is a highly conserved catabolic process for cell degradation, which is lysosomal dependent and essential for maintaining cellular homeostasis. The degraded components need to be sequestrated by double-membrane vesicles called autophagosomes, degraded by lysosomal enzymes after fusing with lysosomes, and finally complete autophagy [4]. In the beginning, autophagy was thought to be non-selective, which is called bulk autophagy. However, an increasing amount of evidence shows that autophagy can selectively degrade misfolded proteins and damaged organelles. The main factors involved in selective autophagy include autophagy receptors and adaptor proteins, which connect substrates to autophagy devices. According to the different substrates, selective autophagy can be divided into various subcategories, such as mitochondria (mitophagy), liposome (lipophagy), endoplasmic reticulum (reticulophagy), pathogens (xenophagy), peroxisomes (pexophagy), ribosomes (ribophagy), and aggregated proteins (aggrephagy) [5].
2. Mitophagy
Mitochondrial homeostasis maintained by mitochondrial dynamics and mitophagy is important for the generation of energy, cellular homeostasis, steroidogenesis, and regulation of apoptosis [13][6]. Of note, mitophagy is a process that the cell selectively wraps and degrades damaged or superfluous mitochondria through autophagy, thereby maintaining mitochondrial homeostasis [14][7]. In addition, mitophagy has been implicated in the pathogenesis of cardiovascular disease and neurodegenerative diseases [15,16][8][9]. When mitochondria are damaged, mitochondria will split, and the damaged mitochondrion will be cleared by mitophagy to maintain the normal function of the mitochondria [13][6]. Thus, mitophagy could help to produce a new smaller healthy mitochondrion that is essential to the recycling of mitochondria function.
Although it has been reported that mitochondria can be degraded by bulk autophagy, the mechanism of mitophagy and bulk autophagy is different. Usually, the mechanisms by which the LC3 adaptor recognizes mitochondrial proteins and induces mitophagy can be divided into ubiquitin-dependent and ubiquitin-independent mechanisms. The induction of mitophagy via ubiquitin-dependent mechanisms can be further mediated by the PINK1 (a mitochondrial serine/threonine-protein kinase)/Parkin (a cytosolic E3 ligase)-dependent pathway or PINK1-dependent, Parkin-independent pathway [17][10]. PINK and Parkin were initially found to be associated with Parkinson’s disease [18][11]. Concretely, under normal circumstances, PINK1 can be imported into mitochondria through the translocase of the outer membrane (TOM) and the translocase of the inner membrane (TIM), so that the mitochondrial targeted sequence of PINK1 is cleaved by mitochondrial processing peptidase in the matrix, and PINK1 protein is degraded by protease presenilin-associated rhomboid-like protein (PARL) on the inner mitochondrial membrane (IMM) [19][12]. In contrast, when mitochondria are damaged, since the potential of the mitochondrial membrane decreases and the mitochondrial membrane depolarizes, PINK1 can only pass through the outer mitochondrial membrane (OMM), and PINK1 cannot enter the mitochondria to be degraded. At this time, PINK1 accumulates on OMM and is activated by phosphorylation [20][13]. The activated PINK1 phosphorylates ubiquitin at Ser65 to recruit and activate PARKIN ubiquitin ligase activity. Then, PARKIN produces polyubiquitin chains, which are recognized by autophagy receptors, including P62, OPTN, NDP52, TAX1BP1, and NBR1 [21][14]. Thus, receptor proteins will recruit mitochondria to the forming autophagosomes for degradation. Above is the PINK1/Parkin-dependent mitophagy pathway. In addition to the PINK1/Parkin-dependent mitophagy pathway, PINK1 can recruit OPTN and NDP52 to mitochondria in the absence of Parkin, and further recruit ULK1, DFCP1, and WIPI1 to induce the Parkin-independent mitophagy pathway [22][15]. Moreover, there are some proteins on the mitochondrial membrane that can directly recognize LC3 and induce mitochondrial autophagy directly, including NIX, BNIP3, FUNDC1, BCL2L13, FKBP8, and NLRX1 [22][15]. Among them, NIX and BNIP3 interact with LC3 through their BH3 domain, further inducing mitophagy [23[16][17],24], while FUNDC1 directly binds to LC3 to induce mitochondrial autophagy under hypoxic conditions [25][18]. BCL2L13, FKBP8, and NLRX1 directly bind to LC3 through their LIR motifs and induce mitophagy [26,27,28][19][20][21]. The ubiquitin-dependent and ubiquitin-independent pathways of mitophagy are illustrated in Figure 1. Importantly, it has been shown that defective mitophagy impairs spermatogenesis as discussed below.
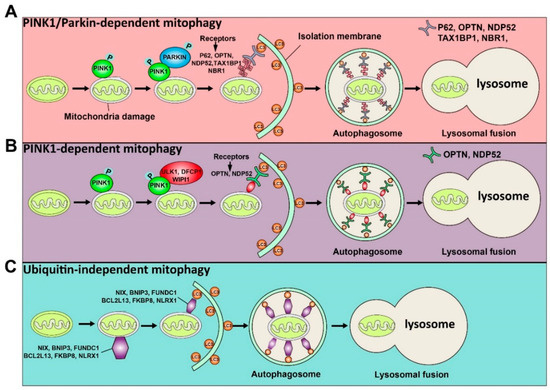
Figure 1. Schematic illustration of the ubiquitin-dependent and ubiquitin-independent pathways of mitophagy is shown. The ubiquitin-dependent mitophagy can be divided into PINK1/Parkin-dependent mitophagy (A) and PINK1-dependent mitophagy (B). (A) Once mitochondria are damaged, PINK1 will accumulate on the OMM (outer mitochondrial membrane) and activate it by phosphorylation. The activated PINK1 recruits and activates PARKIN by phosphorylation. Then, the activated PARKIN produces the polyubiquitin chains recognized by receptors (P62, OPTN, NDP52, TAX1BP1, and NBR1). These receptors will bind with the LC3 adaptor to engulf mitochondria to complete autophagy. (B) The activated PINK1 recruits ULK1, DFCP1, and WIPI1, which can be recognized by receptors (OPTN and NDP52) to induce PINK1-dependent mitophagy. (C) Some proteins, including NIX, BNIP3, FUNDC1, BCL2L13, FKBP8, and NLRX1, can directly recognize the LC3 adaptor and initiate the mitophagy process.
3. Lipophagy
The lipid droplets (LDs), which are mainly made up of cholesteryl ester and triglycerides, are the main lipid storage form in living organisms. Its degradation can regulate the process of lipid metabolism to provide energy for cells. There are two main catabolic pathways to degrade LDs in response to nutrient limitation: lipolysis and lipophagy [53][22]. Lipolysis needs a large number of LD-related lipases to release lipids from LDs, the initiation of which needs adipose triglyceride lipase (ATGL) [54][23]. However, lipophagy, a process that releases fatty acids from LDs by autophagy, has more significant lipophagic activity during starvation compared to lipolysis [54,55][23][24]. In addition, lipophagy includes macrolipophagy and microlipophagy. It has been reported that SQSTM1/p62 (sequestosome-1/p62) protein is the key receptor for the specific recognition of LDs during macrolipophagy [56][25]. Interestingly, LDs can be recognized as a selective substrate and sequestered by the autophagosome, and degraded by hydrolase after combining with the lysosome, while in microlipophagy in yeast, LDs contact the vacuole (lysosome) directly with docking sites instead of being engulfed by the autophagosome [57][26]. Lipophagy is associated with fatty liver disease, obesity, renal cell carcinoma, and liver cancer cells [58,59][27][28]. Furthermore, lipophagy plays an essential role in energy metabolism and lipid homeostasis, and is not only closely related to hepatic diseases but also participates in the regulation of spermatogenesis [60][29]. We next focus on the understanding of lipophagy and summarize how lipophagy is involved in the regulation of spermatogenesis.
Lipophagy is regulated by many factors, such as Rab GTPase, transcription factors, hormones, and small molecules. For example, small Rab GTPase is involved in the regulation of fat-soluble proteins, and Rab GTPase can be used as a molecular switch cycling between active GTP and inactive GDP [61][30]. After nutrient depletion, the small GTPase on the LD surface will activate and switch to an active GTP state [62][31]. It is worth mentioning that this activated state will recruit the degradation devices (multivesicular bodies and lysosomes) to the vicinity of LD and degraded LD by lipophagy [63][32]. RAB protein is the most critical member of the Rab GTPase superfamily, and studies have found that Rab protein can affect lipid autophagy and metabolism [64,65][33][34]. Among them, RAB7 mainly participates in the process of autophagosomal maturation and intracellular transport [64][33] and functions in the fusion of autophagosome membranes and late endocytic membranes with the help of SNARE proteins and HOPS tethering complex [66][35]. Besides, it has been well established that RAB10 on the LD surface will be activated during starvation [65][34]. Additionally, Rab10 can promote the degradation of LD through lipophagy by interacting with EH-domain-binding protein 1 and membrane-deforming ATPase EHD2 [65][34]. Moreover, Rab10 knockdown results in increased LD accumulation in hepatocytes [65][34]. Additionally, RAB32 has been shown to co-localize with autophagy markers in Drosophila fat body [67][36]. In addition to RAB proteins, transcription factors can be involved in the regulation of lipid metabolism. For instance, transcriptional factor EB (TFEB) can induce lipophagy during lipid metabolism via the PPARα (Peroxisome proliferator-activated receptor alpha) and PGC1α (Peroxisome proliferator-activated receptor gamma coactivator 1alpha) signaling pathways [68][37]. Another transcriptional factor FOXO1 (Forkhead box protein O1) can trigger lipophagy by upregulating lysosomal acid lipase (LAL) and the autophagy gene Atg14 in adipocytes [69][38]. In addition, it is well known that the mTOR signaling pathway, which is involved in regulation of autophagy, can inhibit the initiation of autophagy and participate in the regulation of lipophagy as a modulator in response to the change of nutrients and hormones, such as glucose, amino acids, and insulin [15][8]. The activity of lipid metabolism and lipophagy will increase in rapamycin (mTOR signaling pathway inhibitor)-treated hepatocytes [70][39]. Interestingly, some small molecules can modulate lipophagy, such as caffeine, tetrandrine, the dietary polyphenol bergamot, and the red wine bioactive resveratrol [59][28].
4. Therapeutic Modulation of Autophagy to Treat Male Infertility
Infertility is defined as failing to achieve a clinical pregnancy after 12 months or more of regular intercourse without contraception [101][40]. In recent years, infertility has become a global public health issue, affecting 15% of all reproductive-age couples. Among these, male factors account for ~25% of cases of infertility cases, especially abnormal semen quality [102,103][41][42]. More and more studies believe that the components of diet and nutrients may be an important factor affecting sperm quality and fertility [103,104][42][43]. For instance, diets rich in calories, trans-fatty acids (TFAs), saturated fats, or cholesterol have a harmful role in spermatogenesis and male fertility [105,106,107][44][45][46]. These bad diet habits often lead to obesity, related to impaired fertility [108,109][47][48]. Previous studies have demonstrated that autophagy was increased in obese individuals by feeding a high-fat diet (HFD) [110,111][49][50]. Jing Yang’s group investigated the role of autophagy in HFD-induced spermatogenesis deficiency [112][51]. They found that the level of autophagy increased in HFD mice, and spermatogenesis and male fertility were also disrupted in HFD mice. To know the correlation between autophagy and spermatogenesis deficiency in HFD mice, they inhibited and induced autophagy by injecting CQ and RAP intraperitoneally. They found that HFD mice subjected to CQ, an inhibitor of autophagy, showed improved spermatogenesis and decreased infertility. Simultaneously, autophagy was also overactivated in sperm samples from obese subfertile male patients [112][51]. Inhibition of excessive autophagy could protect against HFD-induced spermatogenesis deficiency and male infertility. This can provide a new clinical therapeutic method for increasing semen quality and male infertility.
References
- Staub, C.; Johnson, L. Review: Spermatogenesis in the bull. Animal 2018, 12, s27–s35.
- Satouh, Y.; Ikawa, M. New Insights into the Molecular Events of Mammalian Fertilization. Trends Biochem. Sci. 2018, 43, 818–828.
- Zhou, R.; Wu, J.; Liu, B.; Jiang, Y.; Chen, W.; Li, J.; He, Q.; He, Z. The roles and mechanisms of Leydig cells and myoid cells in regulating spermatogenesis. Cell. Mol. Life Sci. 2019, 76, 2681–2695.
- Mizushima, N.; Komatsu, M. Autophagy: Renovation of cells and tissues. Cell 2011, 147, 728–741.
- Stolz, A.; Ernst, A.; Dikic, I. Cargo recognition and trafficking in selective autophagy. Nat. Cell Biol. 2014, 16, 495–501.
- Wang, R.; Wang, G. Autophagy in Mitochondrial Quality Control. Adv. Exp. Med. Biol. 2019, 1206, 421–434.
- Vives-Bauza, C.; Zhou, C.; Huang, Y.; Cui, M.; de Vries, R.L.; Kim, J.; May, J.; Tocilescu, M.A.; Liu, W.; Ko, H.S.; et al. PINK1-dependent recruitment of Parkin to mitochondria in mitophagy. Proc. Natl. Acad. Sci. USA 2010, 107, 378–383.
- Khawar, M.B.; Gao, H.; Li, W. Autophagy and Lipid Metabolism. Adv. Exp. Med. Biol. 2019, 1206, 359–374.
- Liu, J.; Liu, W.; Li, R.; Yang, H. Mitophagy in Parkinson’s Disease: From Pathogenesis to Treatment. Cells 2019, 8, 712.
- Twig, G.; Shirihai, O.S. The interplay between mitochondrial dynamics and mitophagy. Antioxid. Redox Signal. 2011, 14, 1939–1951.
- Ding, W.X.; Yin, X.M. Mitophagy: Mechanisms, pathophysiological roles, and analysis. Biol. Chem. 2012, 393, 547–564.
- Matsuda, N.; Sato, S.; Shiba, K.; Okatsu, K.; Saisho, K.; Gautier, C.A.; Sou, Y.S.; Saiki, S.; Kawajiri, S.; Sato, F.; et al. PINK1 stabilized by mitochondrial depolarization recruits Parkin to damaged mitochondria and activates latent Parkin for mitophagy. J. Cell Biol. 2010, 189, 211–221.
- Park, S.; Choi, S.G.; Yoo, S.M.; Nah, J.; Jeong, E.; Kim, H.; Jung, Y.K. Pyruvate stimulates mitophagy via PINK1 stabilization. Cell. Signal. 2015, 27, 1824–1830.
- Nguyen, T.N.; Padman, B.S.; Lazarou, M. Deciphering the Molecular Signals of PINK1/Parkin Mitophagy. Trends Cell Biol. 2016, 26, 733–744.
- Lazarou, M.; Sliter, D.A.; Kane, L.A.; Sarraf, S.A.; Wang, C.; Burman, J.L.; Sideris, D.P.; Fogel, A.I.; Youle, R.J. The ubiquitin kinase PINK1 recruits autophagy receptors to induce mitophagy. Nature 2015, 524, 309–314.
- Hanna, R.A.; Quinsay, M.N.; Orogo, A.M.; Giang, K.; Rikka, S.; Gustafsson, Å.B. Microtubule-associated protein 1 light chain 3 (LC3) interacts with Bnip3 protein to selectively remove endoplasmic reticulum and mitochondria via autophagy. J. Biol. Chem. 2012, 287, 19094–19104.
- Novak, I.; Kirkin, V.; McEwan, D.G.; Zhang, J.; Wild, P.; Rozenknop, A.; Rogov, V.; Löhr, F.; Popovic, D.; Occhipinti, A.; et al. Nix is a selective autophagy receptor for mitochondrial clearance. EMBO Rep. 2010, 11, 45–51.
- Liu, L.; Feng, D.; Chen, G.; Chen, M.; Zheng, Q.; Song, P.; Ma, Q.; Zhu, C.; Wang, R.; Qi, W.; et al. Mitochondrial outer-membrane protein FUNDC1 mediates hypoxia-induced mitophagy in mammalian cells. Nat. Cell Biol. 2012, 14, 177–185.
- Murakawa, T.; Yamaguchi, O.; Hashimoto, A.; Hikoso, S.; Takeda, T.; Oka, T.; Yasui, H.; Ueda, H.; Akazawa, Y.; Nakayama, H.; et al. Bcl-2-like protein 13 is a mammalian Atg32 homologue that mediates mitophagy and mitochondrial fragmentation. Nat. Commun. 2015, 6, 7527.
- Lim, G.G.; Lim, K.L. Parkin-independent mitophagy-FKBP8 takes the stage. EMBO Rep. 2017, 18, 864–865.
- Zhang, Y.; Yao, Y.; Qiu, X.; Wang, G.; Hu, Z.; Chen, S.; Wu, Z.; Yuan, N.; Gao, H.; Wang, J.; et al. Listeria hijacks host mitophagy through a novel mitophagy receptor to evade killing. Nat. Immunol. 2019, 20, 433–446.
- Dong, H.; Czaja, M.J. Regulation of lipid droplets by autophagy. Trends Endocrinol. Metab. 2011, 22, 234–240.
- Garcia, E.J.; Vevea, J.D.; Pon, L.A. Lipid droplet autophagy during energy mobilization, lipid homeostasis and protein quality control. Front. Biosci. 2018, 23, 1552–1563.
- Singh, R.; Kaushik, S.; Wang, Y.; Xiang, Y.; Novak, I.; Komatsu, M.; Tanaka, K.; Cuervo, A.M.; Czaja, M.J. Autophagy regulates lipid metabolism. Nature 2009, 458, 1131–1135.
- Münz, C. The Macroautophagy Machinery in Endo- and Exocytosis. J. Mol. Biol. 2017, 429, 473–485.
- Vevea, J.D.; Garcia, E.J.; Chan, R.B.; Zhou, B.; Schultz, M.; Di Paolo, G.; McCaffery, J.M.; Pon, L.A. Role for Lipid Droplet Biogenesis and Microlipophagy in Adaptation to Lipid Imbalance in Yeast. Dev. Cell 2015, 35, 584–599.
- Baerga, R.; Zhang, Y.; Chen, P.H.; Goldman, S.; Jin, S. Targeted deletion of autophagy-related 5 (atg5) impairs adipogenesis in a cellular model and in mice. Autophagy 2009, 5, 1118–1130.
- Schulze, R.J.; Sathyanarayan, A.; Mashek, D.G. Breaking fat: The regulation and mechanisms of lipophagy. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 1178–1187.
- Tarique, I.; Vistro, W.A.; Bai, X.; Yang, P.; Hong, C.; Huang, Y.; Haseeb, A.; Liu, E.; Gandahi, N.S.; Xu, M.; et al. LIPOPHAGY: A novel form of steroidogenic activity within the LEYDIG cell during the reproductive cycle of turtle. Reprod. Biol. Endocrinol. 2019, 17, 19.
- Li, G.; Marlin, M.C. Rab family of GTPases. Methods Mol. Biol. 2015, 1298, 1–15.
- Stenmark, H. Rab GTPases as coordinators of vesicle traffic. Nature reviews. Mol. Cell Biol. 2009, 10, 513–525.
- Carmona-Gutierrez, D.; Zimmermann, A.; Madeo, F. A molecular mechanism for lipophagy regulation in the liver. Hepatology 2015, 61, 1781–1783.
- Feng, Y.; Press, B.; Wandinger-Ness, A. Rab 7: An important regulator of late endocytic membrane traffic. J. Cell Biol. 1995, 131, 1435–1452.
- Li, Z.; Schulze, R.J.; Weller, S.G.; Krueger, E.W.; Schott, M.B.; Zhang, X.; Casey, C.A.; Liu, J.; Stöckli, J.; James, D.E.; et al. A novel Rab10-EHBP1-EHD2 complex essential for the autophagic engulfment of lipid droplets. Sci. Adv. 2016, 2, e1601470.
- Balderhaar, H.J.; Ungermann, C. CORVET and HOPS tethering complexes coordinators of endosome and lysosome fusion. J. Cell Sci. 2013, 126, 1307–1316.
- Wang, C.; Liu, Z.; Huang, X. Rab32 is important for autophagy and lipid storage in Drosophila. PLoS ONE 2012, 7, e32086.
- Settembre, C.; De Cegli, R.; Mansueto, G.; Saha, P.K.; Vetrini, F.; Visvikis, O.; Huynh, T.; Carissimo, A.; Palmer, D.; Klisch, T.J.; et al. TFEB controls cellular lipid metabolism through a starvation-induced autoregulatory loop. Nat. Cell Biol. 2013, 15, 647–658.
- Lettieri Barbato, D.; Tatulli, G.; Aquilano, K.; Ciriolo, M.R. FoxO1 controls lysosomal acid lipase in adipocytes: Implication of lipophagy during nutrient restriction and metformin treatment. Cell Death Dis. 2013, 4, e861.
- Liu, K.; Czaja, M.J. Regulation of lipid stores and metabolism by lipophagy. Cell Death Differ. 2013, 20, 3–11.
- Chandra, A.; Martinez, G.M.; Mosher, W.D.; Abma, J.C.; Jones, J. Fertility, family planning, and reproductive health of U.S. women: Data from the 2002 National Survey of Family Growth. Vital Health Stat. Ser. 23 Data Natl. Surv. Fam. Growth 2005, 25, 1–160.
- Boivin, J.; Bunting, L.; Collins, J.A.; Nygren, K.G. International estimates of infertility prevalence and treatment-seeking: Potential need and demand for infertility medical care. Hum. Reprod. 2007, 22, 1506–1512.
- Salas-Huetos, A.; Bulló, M.; Salas-Salvadó, J. Dietary patterns, foods and nutrients in male fertility parameters and fecundability: A systematic review of observational studies. Hum. Reprod. Update 2017, 23, 371–389.
- Abbasi, A.A.; Prasad, A.S.; Rabbani, P.R. Experimental zinc deficiency in man: Effect on spermatogenesis. Trans. Assoc. Am. Physicians 1979, 92, 292–302.
- Rato, L.; Alves, M.G.; Cavaco, J.E.; Oliveira, P.F. High-energy diets: A threat for male fertility? Obes. Rev. Off. J. Int. Assoc. Study Obes. 2014, 15, 996–1007.
- Ng, S.F.; Lin, R.C.; Laybutt, D.R.; Barres, R.; Owens, J.A.; Morris, M.J. Chronic high-fat diet in fathers programs β-cell dysfunction in female rat offspring. Nature 2010, 467, 963–966.
- Morgan, D.H.; Ghribi, O.; Hui, L.; Geiger, J.D.; Chen, X. Cholesterol-enriched diet disrupts the blood-testis barrier in rabbits. American journal of physiology. Endocrinol. Metab. 2014, 307, E1125–E1130.
- Mitchell, M.; Bakos, H.W.; Lane, M. Paternal diet-induced obesity impairs embryo development and implantation in the mouse. Fertil. Steril. 2011, 95, 1349–1353.
- Palmer, N.O.; Bakos, H.W.; Fullston, T.; Lane, M. Impact of obesity on male fertility, sperm function and molecular composition. Spermatogenesis 2012, 2, 253–263.
- Liu, Y.; Palanivel, R.; Rai, E.; Park, M.; Gabor, T.V.; Scheid, M.P.; Xu, A.; Sweeney, G. Adiponectin stimulates autophagy and reduces oxidative stress to enhance insulin sensitivity during high-fat diet feeding in mice. Diabetes 2015, 64, 36–48.
- Sun, Q.; Nie, S.; Wang, L.; Yang, F.; Meng, Z.; Xiao, H.; Xiang, B.; Li, X.; Fu, X.; Wang, S. Factors that Affect Pancreatic Islet Cell Autophagy in Adult Rats: Evaluation of a Calorie-Restricted Diet and a High-Fat Diet. PLoS ONE 2016, 11, e0151104.
- Mu, Y.; Yan, W.J.; Yin, T.L.; Zhang, Y.; Li, J.; Yang, J. Diet-induced obesity impairs spermatogenesis: A potential role for autophagy. Sci. Rep. 2017, 7, 43475.