1. Lymphatic-Induced Tumor Cell Dissemination by Exosome Secretion
The lymphatic system plays a critical role in immune cell tolerance. Given that secreted factors drain into the lymph before dilution into the blood, lymph fluid is thought to be enriched in cancer biomarkers. It is also postulated that cancer-derived exosomes can promote immune cell tolerance by reprogramming T cells in the lymph fluid
[1][61].
Several groups, including Broggi et al., identified and characterized exosomes in the lymphatic system of melanoma patients. They demonstrated that melanoma cells could reprogram resident immune cells in the lymph to protect against degradation or an immune response. Lymph collected from melanoma patients undergoing lymph node dissection was enriched in exosomes containing melanoma proteins and miRNA compared to plasma. Distinct proteomic signatures were identified in patients with extra-nodal spread compared to patients with sentinel lymph node proteome. Notably, the exosomes from the extra-nodal spread were enriched with S100 proteins, a well-characterized melanoma antigen previously shown to promote pre-metastatic niche formation
[2][62].
A study by Ekstrom et al. isolated exosomes from the lymphatic drainage of breast cancer patients during axillary lymph node dissection and identified a subset of markers that contained CSC markers and markers associated with platelets
[3][63]. Overall,
the studies from both groups provide evidence that the lymphatic system is an additional source for exosome transport that seems to be involved in promoting metastasis (
Figure 1). Their findings can be extrapolated as biomarkers of tumor progression.
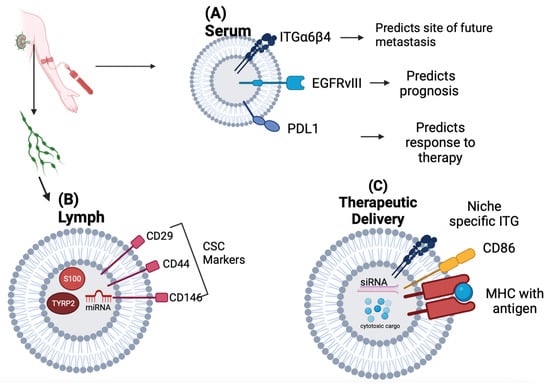
Figure 1. Summary of potential exosome diagnostic and therapeutic applications schematic. (A) Exosome surface markers can be detected in patient serum, providing insights on the prognosis and response to treatment. The identification and quantification of the expression of organotrophic integrins can predict sites of current or future metastases (lung metastasis is associated with increased serum ITGβ4 and liver metastasis with increased ITGαv). The exosome expression of pathogenic receptor EGFRvIII may stratify a subset of glioblastoma patients with a more aggressive disease. The serum levels of exosome PD-L1 may predict the response to chemotherapy and overall survival in lung cancer patients. (B) Lymph collected during axillary lymph node dissection can be an abundant source of tumor-derived exosomes and may be helpful in stratifying patients with increased metastasis and a more aggressive disease. Melanoma tumor-derived exosomes in patients with extra-nodal spread beyond the sentinel lymph node have been shown to be enriched in melanoma-specific miRNAs, S100, and TYRP2, among other melanoma-specific proteins. Exosomes from the lymph of breast cancer patients have been shown to express cancer stem cell markers CD29, CD44, and CD146. (C) Designing synthetic exosomes embedded with organotrophic integrins may be an intriguing strategy in delivering drug cargos to pre-metastatic niches. Therapeutic exosomes can be loaded with antisense oligonucleotides targeting oncoproteins or oncogenic miRNAs, as well as small molecule cytotoxic agents, to counter p-glycoprotein efflux pumps on tumor cells. Dendritic cell-derived exosomes expressing MHC and co-stimulatory domains can induce antigen-specific immune responses against tumors. Figure 1 was created with BioRender.com.
1.1. Challenges of Managing Metastatic Disease
A fSome
w challenges associated with diagnosing and treating metastatic disease
include: metastasisare that it can occur
s before a diagnosis
where disseminated tumor cells undergoand undergo cellular dormancy before the primary tumor is detected, a
nd dormant cells ars well as the difficult
to treay treating dormant
[4]cells [64]. Skeletal metastasis of unknown primary (SMUP) is a condition in which cancer cells have spread from an unknown primary tumor to the bones and presents a new challenge in diagnosing and treating this type of metastasis. This is a rare condition, accounting for 2% of all cancers. The most common sites of metastases are the spine, ribs, pelvis, and skull, which are associated with increased skeletal risk
[5][65]. In a retrospective study of 286 SMUP patients after their initial visit, about 89% of the patients’ tumor origins were detected. Using either blood, a bone biopsy, or CT scans, tumors in the bone were diagnosed with primary tumors of origin as lung cancers (25%), multiple myeloma as the second-leading cancer identified, and about 4% were from breast cancer via blood draws and CT scans
[6][7][66,67]. SMUP presents a clinical challenge due to the lack of standardized diagnostic tools, complete understanding of the underlying mechanism that causes SMUP, and incomplete clinical data; therefore, there is a need for diagnostic and therapeutic strategies to treat patients. The role of exosomes and the presence of tumor secretome such as CTCs are understudied in SMUP. In both cases, they may provide insight into tumor origins. Currently, chemotherapy and radiation therapy are the most common treatments, with surgery as a palliative treatment option, perhaps implementing bone nanoplatforms loaded with bisphosphonates and chemotherapy as a treatment option for patients.
1.2. Exosome-Induced Cellular Dormancy
Metastatic-promoting traits are obtained at the invasive front of the primary tumor and appear to be selective for what is most advantageous for tumor progression at both local and distant sites
[8][68]. It was suggested by Bainer et al. that primary breast tumor cells reprogram local stromal cells by using transcriptional regulation to phenocopy the metastatic state of the tumor, while their nonmetastatic tumor model had an enhanced proliferative state but was not invasive, indicating that transcriptional changes differ based on the metastatic status of the tumor
[9][69]. Disseminated CSCs can exist in a quiescent state in the BM for decades, evading chemotherapy and endogenous immune responses. Mesenchymal stem cells (MSCs) that surround the central sinus have been shown to control CSC entry into the BM and traffic them towards the endosteum, where they undergo cellular dormancy by forming fap junctions with the bone marrow stroma
[10][70]. An exchange of exosome cargo is involved with the recruitment and reprogramming of MSCs into an immunosuppressive phenotype to induce CSC dormancy. Upon priming, MSCs release exosomes containing miR-222/223, which cause cellular quiescence and P glycoprotein expression when taken up by B-CSCs
[11][15]. Developing neoadjuvant therapies that promote resurgence could subject dormant cells to treatment when undergoing chemotherapy and should be further studied.
Subsequent studies have also supported this phenomenon of MSC priming by using metastatic human breast cell lines that secrete exosomes containing miR-23b to induce cellular dormancy chemoresistance by the downregulation of MARCKS, a driver of cell proliferation and motility
[12][71]. Both studies demonstrated how metastatic cancer cells can prime MSCs to secrete exosomes that promote breast cancer dormancy and chemoresistance. Macrophages are critical in modulating breast cancer dormancy within the bone marrow. For instance, immunosuppressive M2 macrophages promote B-CSC quiescence through gap junctional intercellular communication. The lipopolysaccharide stimulation of TLR 4 on MSCs leads to the recruitment and reprogramming of M2 macrophages. M2 macrophages are then converted from an anti-inflammatory to a proinflammatory M1 phenotype. Once transformed into M1 macrophages, they have been shown to secrete exosomes, driving B-CSC migration and enhanced cell cycling via NF-kB activation. Utilizing macrophage polarization to reverse breast cancer dormancy shows potential as a therapeutic adjuvant to chemotherapy. The delivery of M1 macrophage as an adjuvant to carboplatin in vivo leads to increased B-CSC sensitivity to carboplatin and improved mouse survival
[13][72].
Breast cancer dormancy interactions between disseminated metastatic cells and the resident stroma have also been observed in liver metastasis. In a 2D transwell assay, hepatocyte preincubation with MDA-MB-231 primary tumor exosomes increased cancer cell migration and survival. However, in a subsequent 3D human liver micro-physiological system, hepatic milieu preincubation with tumor exosomes increased cancer cell seeding, and hepatic cell proliferation was inhibited. Exosomal miRNA and protein analysis revealed that tumor exosome incubation altered the hepatic niche’s exosomal profile. In addition, the isolation of hepatic niche exosomes and subsequent incubation with aggressive metastatic lines led to the increased expression of E-cadherin, an indicator of a mesenchymal-to-epithelial transition in these cells
[14][73]. This altered secretion promotes metastatic breast cancer cell seeding within the hepatic niche while inhibiting their proliferation and driving a mesenchymal-to-epithelial transition toward cellular dormancy.
2. A Clinical Perspective on Exosomes
For determining whether tumors are present, noninvasive sampling methods such as liquid biopsies are used to isolate tumor-derived exosomes from bodily fluids such as blood, lymphatics, and urine. Circulating exosomes may detect the regression, progression, and recurrence of disease processes. For instance, in bladder cancer, exosomes isolated from urine sub as a liquid easily accessible and consistent with the clinical manifestation of this tumor. Carbonic anhydrase (CA-9) is overexpressed in bladder cancer and is considered a molecular biomarker of this disease. Wen et al. detected CA-9 mRNA in exosomes from 168 bladder cancer patients compared to 90 control patients, with a sensitivity of 85% and specificity of 83%
[15][74], thereby highlighting the versatility of exosome isolation and the reflection of the underlying pathology.
2.1. Exosomes as Biomarkers of Tumor Progression
The optimal biomarkers have high specificity for disease and allow for early detection to promote prompt intervention. However, biomarker development is limited by the dichotomy that, while protein-rich bodily fluids may contain disease-specific markers, they are minimal and difficult to detect. In contrast, biomarkers found in more advanced diseases are abundant but are no longer necessary for early-stage detection and interventions. Exosomes offer a more organized and tissue-specific approach to biomarker development based on the intricate packaging and secretion process
[16][75].
Multiple studies have shown the value of exosomes in monitoring resistance to therapy. The mechanisms by which this occurs may vary. Previous works from Corcoran and others provide evidence that docetaxel resistance is directly transferable by exosomes using the multidrug resistance-1 protein, known to efflux taxanes out of the cell
[17][18][76,77]. Conversely, other studies have reported that the exosome transport of microRNAs promotes post-transcriptional modifications that support tumor progression. One study showed that exposure to 5-FU increased microRNA secretion in colon cancer cells, resulting in altered intracellular physiology and resistance to treatment
[19][78], while another study also uncovered that microRNA transport through exosomes decreased the expression of CyclinD1 in glioblastoma cells, resulting in cellular quiescence and chemoresistance
[20][79].
Serum exosomes may show promise as biomarkers of cancer prognosis compared to bone marrow aspirates commonly used in clinics to determine bone marrow metastasis. Lung cancer patients who showed a response to chemotherapy and overall improved survival were noted to have lower levels of
PD-L1-derived exosomes
[21][80]. Studies have also shown that pathogenic receptor
EGFRvIII may be “shared” among glioblastoma cells through exosomes. In this context, Skog et al. confirmed that exosomes might allow for the secretion of pathogenic receptor
EGFRvIII and are readily detected in the serum of patients with glioblastoma
[22][81]. Thus, exosome proteins such as
PD-L1 and
EGFRvIII may serve as plasma markers for cancer prognosis. In addition to serving as direct biomarkers of diseases, it is now evident that exosomes isolated from body fluids can be analyzed for their donor and recipient cell types based on their cargos and surface markers (
Figure 14A). Finally, exosomal integrins can also serve as biomarkers to predict metastasis
[23][82]. Regardless of the tumor type, patients with lung metastasis were found to have elevated levels of exosome integrin β4 compared to those with liver or no metastasis. Patients with liver metastasis had higher levels of exosomal integrin αv than those with lung or no metastasis. Pancreatic cancer patients that went on to develop liver metastasis had the highest levels of integrin αv at the time of diagnosis.
In summary, exosomes from various bodily fluids may serve as functional biomarkers for cancer detection, resistance to therapy, and disease progression. In addition, exosome isolation from bodily fluids is a minimally invasive method to assess pathology and malignancy mechanisms. Further development for GLP grade isolation and characterization markers will enable reusearchers to adopt isolation techniques for clinical application.
2.2. Exosomes as Delivery Systems
In the study by Kamerkar et al., transfected exosomes with a siRNA targeting an oncogenic
KRAS G12D mutant in pancreatic ductal adenocarcinoma
[24][83]. Upon repeated intraperitoneal injections in mice with orthotopic PDAC tumors, the exosomes displayed staunch retention in the bloodstream, with effective tumor uptake, reduced
KRAS G12D mRNA levels, impaired tumor growth, and improved survival over 30 days. This technology is currently being utilized in a phase I clinical trial on patients with advanced stage PDAC at MD Anderson (NCT03608631) (
Table 1).
In addition to delivering antisense oligonucleotides, exosomes can be loaded with small molecule chemotherapeutics. For example, macrophage-derived exosomes loaded with paclitaxel through sonication were incubated with a malignant canine kidney cell line expressing P-glycoprotein. The loaded exosomes displayed enhanced cytotoxicity compared to free paclitaxel in both drug resistance and sensitivity
[25][84]. Furthermore, in a P-glycoprotein-positive murine model of pulmonary metastasis, exosome-loaded paclitaxel therapy conferred reduced tumor growth compared to free paclitaxel (
Figure 1C).
Cancer vaccination strategies utilizing exosomes loaded with antigenic peptides have been evaluated in animal and clinical trials
[26][85]. Dendritic cell-derived exosomes have received attention because they express MHC I and II and CD86 and can activate CD4+ and CD8+ T-cell responses. It has been demonstrated that dendritic exosomes can induce antigen-specific immune responses several ways, including endocytosis and the internalization of antigenic peptides on dendritic cells. These peptides are then displayed by surface MHC molecules, the direct transfer of MHC/antigen complexes to the dendritic cell surface, and the direct transfer of MHC/antigen complexes to the tumor cell surface, the latter triggering direct T cell-mediated tumor targeting
[27][86]. In a phase I study, a treatment with autologous dendritic cell exosomes loaded with genes from melanoma tumor antigens was well tolerated in non-small cell lung cancer patients. In a subset of these patients, antigen-specific T-cell activation was also observed
[28][87]. Building from this
wor
esearchk, it has been found that stimulating dendritic cells with IFN-γ prior to exosome collection triggers increased T- and NK-cell responses in vitro and may confer a more immunogenic vaccine
[29][88]. In a phase II clinical trial (NCT01159288), allogeneic IFN-γ mature Dex loaded with MHC-restricted cancer antigens was administered as maintenance immunotherapy in 22 patients with unresectable NSCLC. Dex therapy enhanced the NK-cell responses, and seven patients had tumor progression stabilization for >4 months. Additionally, in a phase I/II study in Japan, SART1-pulsed Dex enhanced cytotoxic T-cell responses in patients with advanced squamous cell carcinoma of the esophagus
[30][89].
Identifying the genetic material and its use in promoting cellular reprogramming provides potential therapeutic strategies to reduce metastasis
[32][33][90,91]. To achieve successful treatments, it is essential to target the desired cells or tissues efficiently, avoid off-target effects, and ensure stability and reproducibility while overcoming barriers such as the immune system and the blood–brain barrier.
Great strides have been made to overcome some of the challenges associated with immune cell activation, including engineering exosomes with immune-suppressing agents such as “self” antigens or coating them with molecules that prevent immunogenicity to increase their delivery to recipient cells
[34][35][92,93]. Additionally, viral nanoparticles are nanocarriers with modified viral capsids that can deliver genetic material such as siRNA or mRNA to cells without invoking an immune response due to their high permeability specificity
[36][94]. Comprehensive studies are necessary to assess the safety and immunogenicity of exosome-based therapies or viral nanoparticles. This includes investigating potential immune responses, evaluating long-term effects, and identifying strategies to mitigate immunogenic reactions.
Targeting glioblastoma has been challenging due to the blood–brain barrier. Munoz et al. used MSC-mediated delivery to inhibit TMZ resistance
[32][90], demonstrating that MSCs could cross the BBB and circumvent the bioaccumulation of nascent exosomes, which normally accumulate in the liver or spleen after injection. Others have engineered exosomes with modified surface proteins that are similar to proteins found in brain cells. Other strategies include fusing exosomes to liposomes as a delivery system to the brain
[37][95]. Whether using cells or synthetic exosomes to cross the blood–brain barrier, the size and composition must be small enough to traverse this area. In addition, the use of nanotechnology-based approaches such as liposomes, nanoparticles, and viral vectors can enhance the delivery of exosomes across the blood–brain barrier or other biological barriers.
2.3. Nanotechnology-Based Approaches
Nanocarriers are a type of nanotechnology that is used to deliver drugs or therapeutic agents in a targeted manner and can be engineered to have specific properties, such as size, shape, surface charge, and functional groups, that can enhance their stability, solubility, and targeting ability. Nanocarriers are typically engineered to have specific properties that allow them to encapsulate or attach therapeutic agents and transport them to the desired site in the body. Furthermore, nanocarriers may provide solutions to rectify off-target effects and overcome immune system and blood–brain barriers that thwart clinical exosome applications. For example, core–shell nanocarriers containing miRNA coupled with docetaxel were used as codelivery systems in treating metastatic breast cancer, and miR34a encapsulated in the core of the nanocarrier was shielded from RNase degradation, while the shell interacted with the caveolae-mediated pathway to prevent lysosome degradation
[38][96].
Types of nanocarriers include liposomes, polymeric nanoparticles, dendrimers, and micelles
[39][97]. Strategies based on inhibiting or replacing miRs have emerged as promising approaches in cancer therapy, given that miRs can act as either oncogenes or tumor suppressors. Exosomes have nanocarrier-like properties and are being used in research to deliver anti-miR, chemotherapy, or ATP molecules for cancer treatment and chemoresistance
[25][40][41][42][84,98,99,100]. For example,
MRX34 is a liposome-encapsulated miRNA that targets tumor suppressors miRNA, miR21, and miR-21 and is overexpressed in many types of cancer, and it plays a role in promoting tumor growth and metastasis.
MRX34 works by downregulating miR-21 expression, thereby inhibiting tumor growth and metastasis.
MRX34 is relatively safe and is well tolerated in patients. There was a phase I/II trial on pancreatic cancer where
MRX34 was shown to reduce tumor sizes in some patients
[43][101]. Further,
MRX34 coupled with dexamethasone pretreatment reduced tumor sizes and improved cancer patient outcomes in seven cancer types, including solid tumors and hematology malignancy
[44][102].
Desantis et al. specifically provided an overview of three different miR-based nanocarriers, liposomes, polymers, and exosomes, and highlighted their potential as codelivery molecules that were used to increase stability and decrease degradation. Further, they described various nanoparticle systems that incorporated fungoides, non-living bacteria, or locked nucleic acid oligonucleotides complexes with anti-miR or mimic miR responses that have been implemented to treat various types of cancer, including multiple myeloma
[45][103]. Although nanocarriers offer precise targeted therapies with increased efficacy, such as miR stability for treating cancer patients, a small percentage of nanocarriers are approved for clinical application
[46][104]. In addition to liposomes and other biomaterials such as dendrimers, PEGs were shown to overcome some clinical barriers such as interpatient and tumor heterogeneity and immune cells and renal clearance, as well as bioaccumulation
[47][105]; however, the microenvironment and bioavailability must be considered when designing personalized treatments.
Exosomes have been modified and engineered in several ways to deliver targeted drugs, including altering exosome surfaces with specific ligands, vaccines, or antibodies to enhance tumor cell targeting
[48][49][106,107]. Additionally, loading techniques such as electroporation, sonication, and lipid-based approaches have been explored to improve the loading capacity of exosomes with therapeutic cargos.
Engineered exosomes coupled to liposomes, PEGs, or other molecules have also been reported to increase exosome drug delivery in a controllable way and could circumvent pharmacokinetic drawbacks
[50][108]. Poly(amidoamine) PAMAM dendrimers can be utilized as a loading technique for exosomes to achieve the encapsulation or attachment of specific cargo molecules. The study from Nair et al. used the PAMAM dendrimer–exosome hybrid approach to increase the delivery of siRNA and
PD-L1 cargo to cells with greater efficacy than dendrimers alone, which seemed to be exosome-driven, thereby evading the concerns associated with exosome transport or premature degradation before reaching its target
[51][109].
Nanocarriers can address some challenges associated with exosome therapeutic potential. For example, encapsulating exosomes in nanocarriers would provide a universal size distribution, and their cargo composition would also be uniform. Additionally, adding surface modifications to exosome-loaded nanocarriers could increase the exosome circulation time and biodistribution. Finally, given that nanocarriers can be synthesized and manufactured in large batches, identifying exosome cargo and loading them into nanocarriers could address the concerns regarding the variations in the isolation methods or exosome heterogeneity.