1. Introduction
Atherosclerosis is a chronic disease that affects medium and large arteries of the body through a major increase in the lipoproteins of their intimal layer
[1]. Risk factors associated with atherosclerosis include diabetes mellitus, hypertension, dyslipidemia, obesity, and smoking
[2]. Diabetes mellitus is a major public health problem that has been a leading cause for mortalities worldwide. Diabetes is characterized by elevated levels of blood glucose, which leads over time to serious damage to the heart, blood vessels, eyes, kidneys, and nerves
[3]. As a result, diabetes mellitus is associated with accelerated atherosclerosis, leading to vascular lesions that include cardiovascular disease (CVD), coronary artery disease, cerebrovascular disease, and peripheral arterial disease (PAD), with CVD being the major cause of premature death in diabetes. Another consequence of diabetes mellitus is microangiopathy that occurs in the colon, and which has been reported to be more common in diabetics than non-diabetics
[4]. Microangiopathy can also occur in the retina, skin (specifically foot skin/diabetic foot ulcer), nerve, kidney, muscle, and heart of diabetic patients and is associated with the thickening of the capillary basement membrane
[5]. Other mediators of diabetes that cause vascular complications include dyslipidemia, chronic hyperglycemia, and insulin resistance. Dyslipidemia and chronic inflammation are among the major causes of the development of atherosclerosis, which causes chronic accumulation of lipid-rich plaque in the arteries in diabetic patients
[6][7][6,7]. Therefore, the regulation of these chronic diseases is crucial.
The human gut microbiome is a huge microbial community that plays a vital role in human health. With the development in research, the influence of intestinal flora on human diseases has been gradually revealed. Dysbiosis in the gut microbiota (GM) has been reported to have adverse health effects on the human body that lead to a variety of chronic diseases. Regulation of the GM can provide a potential target for the prevention and treatment of disease. The fermentation products of the gut microbiota are by far the most well studied, and they have been described to have a key role in the maintenance of the gut microbial ecology and the modulation of host immunity and metabolic disease
[8][9][10][11][12][8,9,10,11,12]. The major fermentation products of the GM that result from dietary fibers are short chain fatty acids (SCFAs), with the most abundant metabolites being acetate, propionate, and butyrate.
[12][13][12,13]. SCFAs can function as a macronutrient energy source and hormone-like signaling molecules that enter the portal circulation to signal through specific host receptor systems in order to regulate the innate immunity and host metabolism. Most studies linking the GM to disease designate SCFAs as potential disease-moderating or prevention factors in metabolic disease, intestinal immunity, cancer, and liver disease
[12][13][12,13]. Recent reports have indicated that dysbiosis is increased with aging, and that the GM of elderly people is enriched with pro-inflammatory commensals and fewer beneficial microbes
[14]. Dysbiosis is presumed to be the primary cause of age-associated morbidities, and, consequently, the premature death of elderly people
[14]. Gut dysbiosis leads to a disruption of the microbial metabolites, impaired function of the gastrointestinal tract, and increased leakage of the gut
[14]. These events enhance systemic inflammation which is associated with aging, termed inflammaging, and they, consequently, result in aging-associated pathologies
[14].
Trimethylamine (TMA) is a byproduct generated from the gut microbial metabolism of dietary amines such as choline, betaine, and carnitine that originate from animal-based foods
[15][16][15,16]. TMA is absorbed into the portal circulation and is oxidized by the liver into trimethylamine-N-oxide (TMAO) using the flavin monooxygenase enzyme (
Figure 1)
[16][17][16,17]. More attention has been directed upon circulating TMAO due to its pro-inflammatory, pro-atherogenic, and pro-thrombotic properties
[18][19][20][21][22][18,19,20,21,22]. Several factors, such as diet, gut microbial flora, drug administration, and liver flavin monooxygenase activity, influence the plasma TMAO level
[16]. TMAO has been described as vital for lipid balance and for the increase in scavenger receptors, such as CD36 and scavenger receptor class A type 1 (SR-A1), that contribute to the surge of fat accumulation in foam cells, which in turn play a major part in atherosclerotic plaque progress
[16][18][23][16,18,23]. In addition, the hepatic enzyme flavin-containing monooxygenase 3 (FMO3) is considered the most active in converting TMA into TMAO, leading to higher plasma TMAO levels. High levels of plasma TMAO have been linked to an alteration of reverse cholesterol transport
[16][24][16,24], hyperlipidemia and hyperglycemia
[16][24][25][16,24,25], and the overexpression of inflammatory markers including tumor necrosis factor alpha (TNF-α), interleukin-6 (IL-6), c-reactive protein (CRP)
[26][27][26,27], and insulin resistance
[16][25][16,25], which all lead to the promotion of atherosclerosis
[16][19][25][28][29][16,19,25,28,29]. It has also been reported that high plasma levels of TMAO metabolite are related to the prognosis of 5-year all-cause mortality in stable patients diagnosed with peripheral artery disease
[28][30][28,30]. Furthermore, Brunt et al. confirmed that circulating TMAO was high in older compared with younger adults, and that elevated TMAO was correlated with a higher carotid–femoral pulse wave velocity (PWV). Their findings in humans represented the first link between the age-related increase in circulating TMAO with higher aortic stiffness and blood pressure (BP). Nevertheless, as both aortic stiffness and BP are aging risk factors, it might be that both outcomes are in fact not causally associated with TMAO
[31].
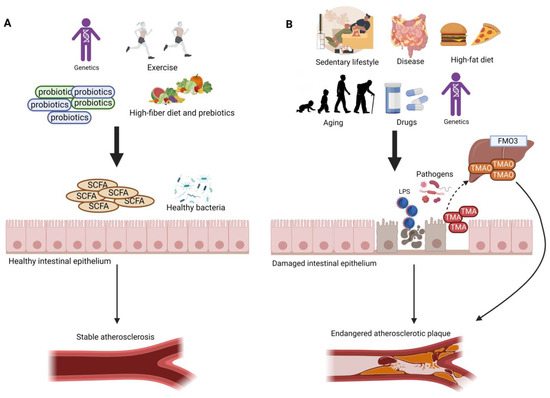
Figure 1. Different factors, such as the gut microbiome, diet, lifestyle, and genetics, play a major role in the development of atherosclerosis. (A) High-fiber diet, active lifestyle, and the intake of probiotics increase the abundance of beneficial gut bacteria and the amount of SCFAs, which support the tight junctions and protect the intestinal epithelium, thus preventing harmful metabolites from entering the circulation. This contributes to a stable atherosclerotic plaque. (B) High-fat diet, sedentary lifestyle, intestinal disease (e.g., Crohn’s disease, irritable bowel disease), the intake of different drugs (e.g., antibiotics), and aging lead to a disruption in the gut microbial profile, resulting in higher abundance of pathogenic bacteria and lower amounts of SCFAs. These effects lead to damage in the gut epithelium; thus, a leaky gut allows the translocation of LPS, TMA, and other damaging metabolites into the circulation. High amounts of plasma LPS and TMAO result in an endangered atherosclerotic plaque.
2. TMAO, TMA-Producing Gut Bacteria, and Atherosclerosis
Previous studies described that many human gut colonizing bacteria are capable of producing TMA, which leads to an increase of TMAO levels in plasma. These gut bacteria include
Streptococcus sanguinis, Desulfovibrio alaskensis, Desulfovibrio desulfuricans, Acinetobacter, Serratia, Escherichia coli, Citrobacter, Klebsiella pneumoniae, Providencia, Shigella, Achiomobacter, and
Sporosorcine, which belong to the Firmicutes and Actinobacteria phyla.
[16][23][16,23]. On the other hand, bacteria belonging to phylum Bacteroidetes are not able to produce TMA
[16][32][16,32]. Several previous studies have investigated the impact of host factors, such as diet and dietary compounds, on TMAO plasma levels, and it was reported that higher plasma TMAO levels have been linked to an animal-based diet
[33][34][33,34] compared to vegetarians (
Figure 1)
[19][35][19,35]. Mainly, TMA is generated by the enzymes produced by the gut microbiota, and its levels are dependent on the amount of precursors available and the abundance and activity of bacteria catalyzing TMA formation
[36], which compete with the host for these precursors that are usually absorbed as essential nutrients
[36]. As the quantification of TMA producers in the gut is limited, it is more difficult to understand the distribution of these bacteria in the gut and specify their niches, which is important for designing effective and sustainable treatment strategies to minimize TMAO plasma concentrations
[36]. In order to discover the abundance and diversity of TMA-forming gut bacteria, Rath et al. developed assays that targeted key genes encoding enzymes responsible for TMA formation from choline (choline-TMA lyase,
CutC), carnitine (carnitine monooxygenase,
CntA), and betaine (subunit B of betaine reductase,
grdH)
[17]. In addition to the association of TMAO with PAD and atherosclerosis, advancing age has also been strongly linked to TMAO levels. As age increases, the host’s physiology and function are altered. For example, the epithelial integrity of the colon, which is needed to promote the influx of bacterial metabolites, including TMAO, might be reduced
[37][38][37,38]. Rath et al. reported that there was an association between carotid intima-media thickness (IMT) and TMAO only in individuals above 65 years of age, which indicates that aging people are principally affected by this metabolite
[38]. Age-related associations between TMAO plasma levels and health parameters have not yet been reported in patients with PAD. Preclinical studies in both animal and in vitro models on human-derived material have emphasized the contribution of TMAO to endothelial senescence and vascular aging
[39][40][39,40]; however, age-specific effects of TMAO have still not been fully studied. Many studies have investigated the link between diet and TMAO levels, reporting different results; however, an analysis of gut microbiota has only recently been included
[38]. Rath et al. were able to provide important information regarding the formation of TMAO in the general population, and they have elaborated on the functional role of the gut microbiota and specific foods, clarifying the increased levels of TMAO with increasing age
[38].
3. Gut Dysbiosis, Aging, and TMAO Levels
Gut dysbiosis is the disruption in the gut microbiome that is associated with different diseases. Dysbiosis disturbs the gut barrier function, leading to the leakage of harmful metabolites, such as lipopolysaccharides (LPS), and other bacterial components, such as peptidoglycans, into the circulation, which triggers an inflammatory response leading to atherosclerosis (
Figure 1)
[41][42][43][41,42,43]. LPS can stimulate the uptake of modified low-density lipoprotein (LDL) and reduce the efflux of cholesterol from foam cells, promoting monocyte recruitment and macrophage foam cell formation
[41][43][44][41,43,44]. LPS can induce vascular inflammation directly or by producing pro-inflammatory factors from immune cells
[44]. The increased production of pro-inflammatory cytokines promotes oxidative stress and oxidized LDL (oxLDL), increasing the risk of hypertension via nitric oxide synthase inhibition. This effect reduces vasodilator nitric oxide levels and increases levels of vasoconstrictor endothelin-1
[41].
Aging, Gut Dysbiosis, and TMAO
Aging leads to several changes in cells, tissues, and organs
[45] and is influenced by an individual’s genetics, lifestyle, and environment
[46]. The term “immunoscence” first appeared a few decades ago to refer to impaired or faulty immune responses leading to a decrease in the ability to trigger the immune response and effectively produce antibodies against different pathogens
[47][48][47,48]. The gut microbiome undergoes dynamic changes through time, and gut dysbiosis is an age-related complication caused by host senescence, changes in nutritional behavior, drug use, and the lifestyle of aged people
[48]. The changes in the gut microbiome include shifts in bacterial composition and metabolic function
[49]. In humans, age-related gut dysbiosis is characterized by increased inter-individual variation and decreased species diversity; specifically, a loss of Clostridiales and Bifidobacterium, an enrichment of Proteobacteria, Lactobacilli, and an overrepresentation of pathobionts such as Enterobacteriaceae
[49][50][51][52][49,50,51,52]. However, the major gut microbiota aging feature is the decreased ratio of Firmicutes/Bacteroidetes
[53]. Schneeberger et al. reported that aged mice showed a decrease in beneficial gut bacteria, such as in Clostridium members of cluster IV that produce SCFAs and
Akkermansia muciniphila, and an increase in pro-inflammatory microbes
[14][54][14,54]. Overall, the decrease in intestinal commensal microbes diversity is associated with increased susceptibility to pathogen infection accompanied by disturbance of the gut mucosal barrier and enrichment in pro-inflammatory cytokines; all these events have a detrimental consequence in aging
[14][55][14,55]. Recent studies suggested that gut dysbiosis is associated with the development of several chronic diseases including cardiovascular disease and other metabolic disorders
[56].
Many studies have reported a close relationship between TMAO levels, aging, and age-related diseases. Several animal models have been used to identify mechanisms that underlie TMAO’s role in senescence
[57]. Cell senescence involves many processes including increased production of reactive oxygen species (ROS), mitochondrial dysfunction, and senescence-associated secretory phenotype (SASP)
[57]. Ke et al. reported that senescence-accelerated prone mouse strain 8 (SAMP8) and senescence-accelerated mouse resistant 1 (SAMR1) were treated with 1.5% (w/v) TMAO for 16 weeks to induce vascular aging and advanced vascular aging processes, respectively
[40]. Many potential mechanisms underlie TMAO’s role in aging, including the inhibition of sirtuin 1 (SIRT1) expression, which increases oxidative stress and results in the activation of the p53/p21/Rb pathway. Increased P53 and P21 acetylation and reduced CDK2, cyclinE1, and Rb phosphorylation are followed by enhanced endothelial cell senescence and vascular aging
[58]. In addition, TMAO increases the accumulation of ROS, matrix metalloproteinase 2 (MMP2), and matrix metalloproteinase 9 (MMP9) in vivo and in vitro, which are associated to oxidative stress in cells
[59]. Furthermore, high TMAO levels are linked to increased expression of pro-inflammatory cytokines, such as TNF-α and IL-1β, as well as decreased production of anti-inflammatory cytokines such as IL-10
[26].
4. Mechanisms Underlying the Therapeutic Effect of Probiotics in Atherosclerosis
The mechanisms underlying the protective effect of probiotics against atherosclerosis are not fully understood. Nevertheless, the action of probiotics at different steps is becoming clear as more studies are being conducted. To start with, it has been reported in previous studies how probiotics can combat gut dysbiosis through strengthening the epithelial tight junctions, preventing the translocation of damaging metabolites, such as LPS and TMAO, into the circulation, which can lead to a stable atherosclerotic plaque
[60][61]. Several studies have demonstrated the hypocholesterolemic effect of probiotics
[61][62][153,154]. Liong and Shah have pointed out the ability of Lactobacillus strains to reduce cholesterol in an in vitro model, and this was achieved through various mechanisms such as assimilation of cholesterol during growth, incorporation of cholesterol into the membrane of cells, the binding of cholesterol to the cell surface, and co-precipitation with deconjugated bile
[63][155]. These findings were backed up in another study by Zeng et al., who studied
Lactobacillus buchneri P2 and confirmed the cholesterol removal trait of this bacteria through an assimilation mechanism
[64][156]. Another study by Huang and Zheng reported the cholesterol lowering property of a probiotic strain through the inhibition of the gene expression of NPC1L1 in Caco-2 cells
[65][66][157,158]. The NPC1L1 protein plays a major role in cholesterol absorption, and it is considered to be a promising target for cholesterol-lowering medication
[67][159]. NPC1L1 has been identified by Duval et al. to be a novel target gene for the liver X receptors (LXRs), which support the crucial role of LXRs in intestinal cholesterol homeostasis
[68][160]. LXRs activation has been reported to reduce whole-body cholesterol levels and reduce atherosclerosis
[69][161]. In addition, VSL-3 was found to improve lipid profiles in mice
[70][162], and this was attained by promoting BA deconjugation and fecal excretion, and by increasing hepatic BA synthesis through the downregulation of the FXR/FGF15 axis
[71][163]. BAs can regulate cholesterol balance, and disruption in the circulation of enterohepatic BAs can lead to gall bladder
[72][164] and gastrointestinal diseases
[73][165]. The metabolism of BAs is also associated with obesity, diabetes, and cardiovascular diseases
[74][75][76][166,167,168]. BAs are synthesized from hepatic cholesterol, and they are further conjugated with amino acids glycine and taurine to form bile salts that are transferred to the intestine. Bile salts’ amphiphilic combination is crucial for fat absorption in the intestine, yet excessive bile salts are toxic to the gut bacteria
[77][169]. Bile salt hydrolase (BSH), which is present in the gut microbiome, is responsible for the catalysis of the conjugated bile salts into deconjugated BAs in order to maintain the balance of metabolism of BAs. Deconjugated BAs function as signaling molecules to aid in the secretion of GLP-1 hormone
[78][170], activate other receptors, and impact different metabolic processes involved in various diseases
[79][80][171,172]. The presence of BSH has been identified in different microbial genera such as
Lactobacillus, Bifidobacterium, Enterococcus, Clostridium spp., and
Bacteroides. It has also been reported that one bacterial strain can possess distinct BSHs that can have different properties
[81][173]. In addition, recent evidence suggested that TMAO can promote atherosclerosis, partially through inhibiting hepatic bile acid synthesis
[82][110].
In addition, probiotics can apply their anti-inflammatory actions through modulating the expression of key transcription factors or microRNAs (miRNAs), which are associated with pro-inflammatory signaling
[83][84][148,174]. For instance, Chen et al. reported that
L. acidophilus ATCC 4356 was able to reduce the levels of TNF-α and oxidative stress markers in addition to its ability of reversing the reduction in IL-10 levels via inhibiting the activation of NF-κB and its translocation to the nucleus
[83][148]. In the case of atherosclerosis, T lymphocytes and macrophages accumulate and proliferate at the atherosclerotic lesions, which leads to the secretion of inflammatory cytokines such as TNF-α and IL-10
[83][148]. These inflammatory cytokines can, in turn, activate intracellular NF-κB signaling pathways which can stimulate the production of more cytokines, leading to further inflammation
[85][175]. Activated NF-κB exists in the fibrotic thickened intima-media and atheromatous areas of the atherosclerotic lesion, smooth muscle cells, macrophages, and endothelial cells; however, little or no activated NF-κB can be detected in vessels lacking atherosclerosis
[86][87][176,177]. The activation of NF-κB is associated with the phosphorylation of IκB-α and subsequent degradation of IκB-α, which results in the translocation of NF-κB into the nucleus. It has also been reported that TNF-α is one of the inflammatory markers that can promote atherosclerosis. On the other hand, IL-10 is an anti-inflammatory marker that provides a crucial atheroprotective signal
[44]. As for the miRNAs,
Lactobacillus acidophilus has been shown to protect against apoptosis and necrosis in human endothelial cells, which is induced by LPS stimulation, and this in turn was associated with a decrease in the expression of pro-inflammatory miR-155 and increased expression of anti-apoptotic mIR-21
[43][84][43,174].