Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Vittoria Schirinzi and Version 2 by Beatrix Zheng.
Current evidence, both on animal and human models, indicates that several diet components may have beneficial effects on obesity by affecting brown adipose tissue (BAT) and energy metabolism, including polyunsaturated fatty acids, capsaicin and capsinoids, catechins, curcumin, resveratrol and berberine, oleuropein, anthocyanins, quercetin, gingerol, shogaol, 6-paradol, thiacremonone, cinnamaldehyde and menthol.
- brown adipose tissue
- obesity
- thermogenic nutraceuticals
1. Polyunsatured Fatty Acids
Fish oil is rich in the polyunsaturated fatty acids (PUFA) eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), which stimulate thermogenesis in BAT [1][2][3][41,42,43]. In the early 2000s, several studies in mouse models compared the effects of a diet enriched with n-3 PUFAs and a diet rich in n-6 PUFAs in order to study their impact on BAT. It turned out that fats from the n-3 series were much more effective in activating BAT than the n-6 series [4][44]. Therefore, subsequent research focused on the effects of n-3 PUFAs, especially EPA, and confirmed its anti-obesogenic effect, revealing that it can promote the browning of WAT. Rats fed with a supplementation of fish oil at a 20% EPA for 4 weeks showed an increased mRNA level of UCP1 in WAT [5][45]. Another study showed that supplementation with 27.5% fish oil, administered for one month, stimulates SNS-mediated mitochondrial and thermogenic activity in rats [6][46]. A diet enriched with a mixture of EPA and DHA increases the expression of UCP1 in BAT and reduces adipose accumulation via the induction of marked, non-shivering thermogenesis [7][8][47,48]. Further implications emerged from a study conducted in 2016, which revealed that a high-fat diet (HFD), enriched with fish oil (12% of total lipids) and administered to mice for 8 weeks, modulates BAT activation via epigenetic regulation [9][49]. EPA promotes both the adipogenesis of mature brown adipocytes [10][50] and the differentiation of pre-adipocytes into beige adipocytes, within WAT stores, particularly in the inguinal WAT [8][48]. It is important to note that the conversion of mature white adipocytes to beige adipocytes was achieved via supplementation with EPA in human adipocyte cultures [11][51] and that the same effect was not observed in rodents [3][43].
2. Capsaicin and Capsinoids
Capsaicin is an alkaloid found in chilli peppers that is mainly concentrated in the seeds of the fruit and is the main component responsible for the spicy flavour. Capsaicin has a role in preventing the obesogenic effects of diet, due to its ability to increase energy expenditure by activating BAT [12][52]. Capsaicin implements BAT function via several signalling pathways, primarily via the activation of Transient Receptor Potential Vanilloid 1 channels (TRPV1), which stimulate the central nervous system to produce catecholamines involved in thermogenesis. Capsaicin seems to regulate the epigenetic expression of the transcription factors involved in WAT browning [13][14][53,54]. Important evidence has also emerged from studies on humans: Yoneshiro et al. [15][55] examined the acute effect of an oral ingestion of a single dose of capsinoids (9 mg) on energy expenditure, in relation to BAT activity in humans; this was measured by 18F-FDG PET, a classical imaging technique used to visualize BAT activation. The ingestion of capsinoids resulted in a 3-fold increase in energy expenditure in the BAT-positive group compared to the BAT-negative group. Furthermore, the chronic intake of capsinoids (9 mg/day in capsule form) for 6 weeks promoted BAT activity and reduced fat mass after cold exposure, even in human subjects with low BAT activity. In 2018, Camps et al. [16][56] tested the same treatment used in mice in 2016 [17][57] on humans. They gave a group of normal-weight individuals a dose of capsinoids (12 mg), combined with exposure to cold (14.5 °C). The subjects underwent, before and after exposure, both 18F-FDG PET and indirect calorimetry, in order to assess the total energy expenditure. The results showed that capsinoids increased energy expenditure in BAT-positive participants and, when combined with cold, increased fat oxidation, improved insulin sensitivity and increased HDL-cholesterol [18][19][58,59].
3. Green Tea Catechins
Green tea consumption is associated with weight loss and the modulation of fat metabolism and energy expenditure [20][21][60,61]. Catechin supplementation for 8 weeks reduced the mass of perirenal WAT and increased the expression of mRNA coding for UCP1 in BAT, compared to the control group in rats [22][62]. Yan and al. [23][63] showed that the oral administration of catechins (100 mg/kg body weight) for 4 weeks significantly reduced the total fat mass (subcutaneous and visceral) and liver size in rats fed with HFD. No browning of the WAT was observed, but fatty acid oxidation in the BAT increased twofold [23][63]. Several experiments testing the effects of catechin supplementation on rodents were also conducted on humans. The habitual intake of green tea (>300 mg catechins/day) was found to be as effective as in rats in terms of reducing body weight and preventing weight regain [24][64]. Similar results were obtained in a study on subjects with metabolic syndrome, where catechin supplementation additionally improved the lipid profile and hypertension [25][65]. The consumption of 300 mg/day of Epigallocatechin Gallate (EGCG) or 200 mg of caffeine [26][66] for 3 days in subjects with obesity increased postprandial fat oxidation, but not the total energy expenditure [27][67]. A 12-week treatment with green tea extracts containing 583 mg catechins resulted in a small but significant reduction in body fat (3–5%) in healthy, normal-weight subjects [28][68]. A more recent study [29][69] suggested that catechins also act as inhibitors of catechol-O-methyltransferases (COMT), enzymes that degrade catecholamines. The inhibition of COMTs results in the increased metabolism of catecholamines, including noradrenaline, which, as thwe researchers hhave seen, is involved in thermogenesis and fat oxidation processes.
4. Curcumin
Curcumin is the most abundant and active element within the turmeric rhizome; it is a widely studied polyphenol with antioxidant and anti-inflammatory properties, also known for its anti-obesity effects [30][70]. Isolated WAT cells of obese rats, in response to treatment with 20 μM of curcumin for 6–8 days, showed an increase in the thermogenic markers of BAT and in the hormone-sensitive lipase (HSL), which is responsible for triglyceride mobilization and lipolysis in WAT. These results suggest curcumin’s role in improving the lipid profile in an obese condition [31][71]. The administration of high doses of curcumin (45 mg/kg body weight) causes an increase in energy expenditure via the induction of mitochondrial biogenesis in mice [32][72]. Experimental evidence on humans is still limited. However, one significant study in the literature stands out; this was conducted in 2019 on 60 participants [33][34][73,74], with the aim of assessing the effects of curcumin supplementation on cardiovascular risk factors among overweight adolescents. It was a randomized controlled trial involving 60 girls aged 13–18 years, randomly assigned to the intervention or control group. The intervention group was supplemented with 500 mg of curcumin (in bioavailable form) daily for 10 weeks, while the control group received a placebo. At the same time, they were asked to undergo a mild weight loss or weight maintenance diet, depending on the degree of overweight or obesity. The results revealed that curcumin supplementation induced significant improvements compared to the control group, such as a reduction in the body mass index, a reduction in waist circumference and hip circumference, and reduction in the triglyceride/HDL ratio; meanwhile, it induced an increase in the HDL cholesterol. However, clinical studies on a larger population and with a longer duration are needed to confirm the results of these studies [33][34][73,74] and to test the WAT darkening effects observed in rodents also in humans.
5. Resveratrol
Resveratrol is a polyphenol naturally contained in grape skins and other vegetables such as red fruits, cocoa and peanuts [35][75]. The supplementation of 30 mg/kg body weight of resveratrol in mouse models for 8 weeks showed a significant reduction in fat mass, plasma glucose concentrations and total cholesterol, compared to the control group [36][76]. There was also evidence of increased UCP1 expression in both BAT and skeletal muscle [37][77]. This is attributed, at least in part, to the ability of resveratrol to activate upstream AMPK, which promotes the production of PGC1α and Sirtuin 1 (SIRT1); these, in turn, promote mitochondrial biogenesis and WAT browning [38][78]. The effect of resveratrol on the neo-formation of beige adipocytes is thought to be mediated by the phosphorylation of AMPK, as the deletion of this protein in mice abolishes the browning effects of resveratrol [39][79]. A recent study [40][80] revealed the additional signalling pathways exerted by resveratrol. The results reported regarding the cell cultures and animal models suggest that high doses of resveratrol (100 and 200 μM) induce epigenetic regulatory mechanisms. Polyphenol can up-regulate the expression of the genes that code for the proteins involved in WAT adipogenesis, such as fatty acid synthase (FAS), the sterol response element-binding protein (SREBP1), lipoprotein lipase (LPL) and hormone-sensitive lipase (HSL) [41][81]. Overall, resveratrol oversees antiadipogenic and anti-inflammatory effects, which are dependent and independent of BAT activation; however, evidence supporting its thermogenic effects in humans is currently lacking. This is due to the poor bioavailability of polyphenolic compounds [42][82], so it is difficult to achieve effective doses that are able to stimulate BAT through food alone. Diet alone is not sufficient in order to mimic the efficacy of resveratrol observed in rodents.
6. Berberine
Berberine is an alkaloid compound found in the roots, rhizomes and bark of certain plants of the genus Berberis, primarily known for its anti-cancer activity [43][83]. Zhang et al. [44][84] proved that the intraperitoneal administration of berberine, administered at 5 mg/kg/day for 4 weeks in obesity-induced mice, played a key role in increasing energy expenditure and the mobilization of lipids. More specifically, the thermogenic effects attributed to berberine concerned the increase in the mitochondrial content and thermogenic markers in BAT (UCP1, PGC1α, Cidea). The increase in BAT was detected via PET-CT, while the increase in the resting energy expenditure was quantified by measuring the oxygen consumption (VO2) and carbon dioxide release (VCO2) before and after treatment. The total expenditure increased by 20%.
An interesting finding, which did not emerge from studies on the other phytochemical compounds, is that this treatment caused a significant reduction in the respiratory quotient, suggesting that berberine causes a shift in cell metabolism, taking energy from the oxidation of fatty acids, rather than from carbohydrates. Furthermore, mice berberine not only stimulates BAT activity but also induces the browning of the inguinal WAT, probably via the phosphorylation of AMPK, which leads to the increased expression of UCP1 and PGC1α [45][85]. An in vitro study was recently conducted on human preadipocytes, and it was found that the thermogenic effect induced by berberine on WAT depends on the recruitment of the AMPK-PRDM16 axis: the activation of AMPK could lead to DNA demethylation and thus upregulate the expression of PRDM16, which acts as a master regulator of the transdifferentiation of white adipocytes into beige adipocytes; this occurs via the direct modulation of the transcription factors PPARγ and PGC1α [46][47][86,87].
7. Other Nutraceutical Compounds
Many other dietary polyphenolic compounds have been found, in rodent studies, to influence BAT thermogenesis and WAT browning: oleuropein, anthocyanins, quercetin, and structural analogues of capsaicin (i.e., menthol, cinnamaldehyde, allyl and benzyl isothiocyanates, which are the spicy elements found in mustard and wasabi, and thiacremonone). Some of these compounds have also shown promising results in humans.
7.1. Oleuropein
Oleuropein is the main polyphenol contained in olive leaves and fruits, and is responsible for the pungent and bitter taste of raw olives. Olive leaf extract (3 mg of oleuropein, injected intravenously in mice for 7 weeks) increases UCP1 content in BAT by activating SIRT1, PPARγ and PGC1α. In addition, it stimulates the secretion of adrenalin and noradrenalin via the activation of TRP channels [48][88]. Oleuropein aglycone (the absorbed form of oleuropein) appears to be able to attenuate diet-induced obesity by supporting the expression of thermogenic genes and genes related to mitochondrial biogenesis in the BAT of overfed mice [49][89], and to promote the browning of adipose tissue via mesenchymal stem cells in humans [50][90].
7.2. Anthocyanins
Anthocyanins are the polyphenols mainly contained in red fruits, grapes, black soy, and red beans. Anthocyanins are known first and foremost for their strong antioxidant power, but they have also been re-evaluated for their thermogenic action in stimulating adrenalin secretion and energy metabolism in humans; meanwhile, in rodents, they exert an anti-obesity action that is related to BAT activation [51][52][91,92]. In humans, the intake of 150 mg/day of an extract of Aronia melanocarpa, a particular type of blueberry rich in anthocyanins, procyanidins and other flavonoids, increases the surface body temperature and plasma adrenalin levels, suggesting that it has a stimulating effect on the SNS [53][93]. Numerous studies have also shown that long-term treatment with cyanidin-3-glucoside, which is found in raspberry and mulberry extract, increases UCP1 expression and mitochondrial biogenesis during the adipogenic differentiation of brown and white preadipocytes [54][55][94,95]. Similarly, black soybean peel extract upregulates the expression of thermogenic genes in BAT, induces WAT browning and increases the lipid respiration quotient, thus preventing visceral fat accumulation in mice on a hyperlipidic diet [56][96].
7.3. Quercetin
Quercetin is a flavonoid contained in a wide variety of fruits (apples, grapes, olives, citrus fruits, berries) and vegetables (tomatoes, onions, broccoli, and capers). It has a thermogenic effect in mice and an ability to modulate the gut microbiota in mice. Obese mice fed a HFD supplemented with 1% quercetin for 16 weeks lost weight and had reduced plasma cholesterol levels, compared to mice fed a HFD alone [57][97]. The improvement in obesity is explained by the ability of quercetin to increase the expression of thermogenic genes in BAT (UCP1, PGC1α. FGF21), as well as and genes coding for β-adrenergic receptors and AMPK; this results in increased non-excitation thermogenesis. The increase in AMPK also suggests that quercetin may predispose these signalling pathways to WAT browning [58][98]. Other studies point out the flavonoid’s ability to modulate the intestinal microbial composition of mice. Quercetin reduces the ratio of Firmicutes to Bacteroidetes in the microbiota of HFD-fed mice, improving the obesity-related dysbiosis picture. In fact, a eubiotic microbiota allows greater energy extraction from the diet via the increased production of short-chain fatty acids, which have been found in the faeces of mice supplemented with quercetin [57][97].
7.4. Analogues of Capsaicin
Structural analogues of capsaicin include menthol, which is a cyclic monoterpene alcohol obtained from peppermint, the activator of TRPM8 receptors [59][60][99,100], cinnamaldehyde, a pungent compound that increases UCP1 expression in human white adipocytes [61][101] and is found in cinnamon, allyl and benzyl isothiocyanates, which are spicy elements found in mustard, ginger and wasabi (Japanese horseradish), and finally, thiacremonone, a sulphur compound isolated in garlic [62][102]. The compounds just mentioned can activate TRPV1 and TRPA1 channels (also belonging to the TRP receptor family, but activated by cold, mechanical stimuli and cooling nutritional compounds), which modulate thermoregulation. The combination of sub-effective doses of capsaicin, cinnamaldehyde and menthol induce the “brite” phenotype in the differentiation of the 3T3-L1 cells and subcutaneous white adipose tissue of HFD-fed obese mice [63][103]. The intervention prevented adipose tissue hypertrophy and weight gain, and enhanced the thermogenic potential, mitochondrial biogenesis, and overall activation of brown adipose tissue. These changes, observed in vitro as well as in vivo, were linked to the increased phosphorylation of kinases, AMPK and ERK. In the liver, this combination treatment enhanced insulin sensitivity, improved the gluconeogenic potential and lipolysis, prevented fatty acid accumulation and enhanced glucose utilization. Finally, it is worth mentioning that gingerol, shogaol and 6-paradol, which are all contained in ginger root, activate TRPV1 channels [64][65][66][104,105,106]. An interesting study in humans looked at a type of ginger native to West Africa (black ginger), from which the extract Kaempferia parviflora is derived. The administration of 100 mg/day of this extract appears to increase energy expenditure in BAT-positive subjects, and the activation of brown adipocytes is detectable via FDG-PET after exposure to cold [67][107].
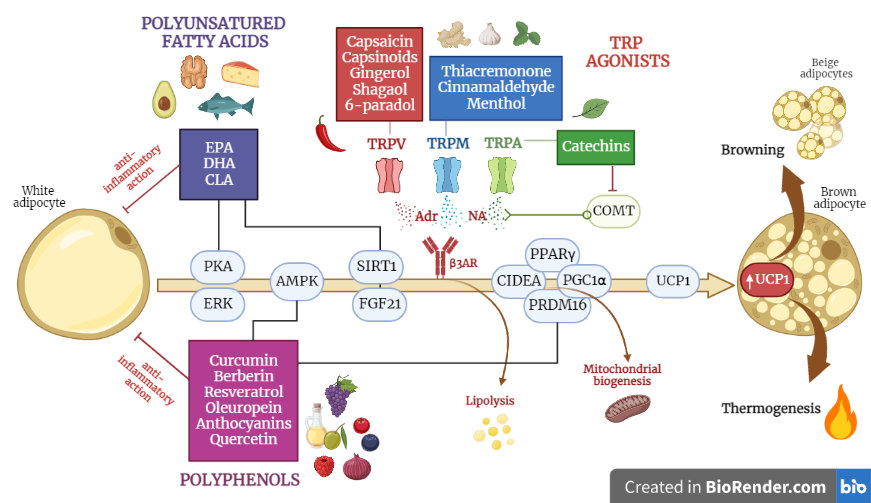
Table 13.
Mechanism of action of the thermogenic nutraceuticals examined.
| Thermogenic Nutraceuticals | Dose | Mechanism of Action | Experimental Model | Ref. | |||||
|---|---|---|---|---|---|---|---|---|---|
| EPA DHA | simulates thermogenesis in BAT | mouse human |
[1]] | [41 | [2] | ,42 | [3][4 | ,43,44] | |
| ↑ expression of UCP1 in BAT, | mouse | [7][8] | [47,48] | ||||||
| ↓ adipose accumulation via the induction of marked, non-shivering thermogenesis, | mouse | [68][69] | [36,37] | ||||||
| promotes the adipogenesis of mature brown adipocytes | mouse | [10] | [50] | ||||||
| promotes the differentiation of pre-adipocytes into beige adipocytes, particularly in the inguinal WAT | mouse | [8] | [48] | ||||||
| Capsaicin | activates TRPV1 channels: implements BAT function | in vitro and pre-clinical studies | [13][14] | [53,54] | |||||
| regulates the epigenetic expression of the transcription factors involved in WAT browning | in vitro and pre-clinical studies | [13][14] | [53,54] | ||||||
| Capsinoids | 9 mg/day in capsule form for 6 weeks | promotes BAT activity and reduces fat mass | human | [15] | [55] | ||||
| 12 mg combined with exposure to cold (14.5 °C) | ↑ energy expenditure and, when combined with cold, ↑ fat oxidation, ↑ insulin sensitivity and ↑ HDL-cholesterol | human | [16][17][18] | [56,57,58] | |||||
| Catechins | for 8 weeks | ↓ mass of perirenal WAT, ↑ expression of mRNA coding for UCP1 in BAT | rat | [22] | [62] | ||||
| 100 mg/kg body weight for 4 weeks | ↓ total fat mass (subcutaneous and visceral) and liver size, fatty acid oxidation in the BAT increased twofold | rat | [23] | [63] | |||||
| >300 mg catechins/day | ↓ body weight and prevents weight regain | human | [24] | [64] | |||||
| inhibits catechol-O-methyltransferases | human | [29] | [69] | ||||||
| Curcumin | 20 μM for 6–8 days | ↑ in thermogenic markers of BAT and in hormone-sensitive lipase (HSL), | isolated WAT cells of obese rats | [31] | [71] | ||||
| 45 mg/kg of body weight | ↑ energy expenditure via the induction of mitochondrial biogenesis | mouse | [33] | [73] | |||||
| 500 mg (in bioavailable form) daily for 10 weeks | ↓ in body mass index, waist circumference and hip circumference, and triglyceride/HDL ratio, and ↑ HDL cholesterol | human | [33][34] | [73,74] | |||||
| Resveratrol | 30 mg/kg of body weight for 8 weeks | ↓ fat mass, plasma glucose concentrations and total cholesterol | mouse | [36] | [76] | ||||
| ↑ UCP1 expression | mouse | [37] | [77] | ||||||
| activates upstream AMPK, which promotes the production of PGC1α, and SIRT1, which promotes mitochondrial biogenesis and WAT browning | mouse | [38] | [78] | ||||||
| up-regulates the expression of genes coding for proteins involved in WAT adipogenesis (FAS, SREBP1, LPL and HSL) | human | [41] | [81] | ||||||
| Berberine | 5 mg/kg/day for 4 weeks | ↑ energy expenditure and the mobilization of lipids | mouse | [44] | [84] | ||||
| stimulates BAT activity, and induces browning of the inguinal WAT | mouse | [45] | [85] | ||||||
| Oleuropein | 3 mg injected intravenously for 7 weeks | ↑ UCP1 content in BAT by activating SIRT1, PPARγ and PGC1α, stimulates the secretion of adrenalin and noradrenalin via the activation of TRP channels |
mouse | [48] | [88] | ||||
| Oleuropein aglycone (the absorbed form of oleuropein) | attenuates diet-induced obesity by supporting the expression of thermogenic genes and genes related to mitochondrial biogenesis in the BAT of overfed mice | mouse | [49] | [89] | |||||
| promotes the browning of adipose tissue from mesenchymal stem cells in humans | in vitro | [50] | [90] | ||||||
| Anthocyanins | 150 mg/day of an extract of Aronia melanocarpa | ↑ surface body temperature and plasma adrenalin levels, suggesting that it has stimulating effect on the SNS | human | [51] | [91] | ||||
| long-term treatment with cyanidin-3-glucoside, such as raspberry and mulberry extract | ↑ UCP1 expression and mitochondrial biogenesis during adipogenic differentiation of brown and white preadipocytes | rat | [52][53] | [92,93] | |||||
| black soybean peel extract | ↑ the expression of thermogenic genes in BAT, induces WAT browning and increases the lipid respiration quotient, preventing visceral fat accumulation on a hyperlipidic diet | mouse | [54] | [94] | |||||
| Quercetin | HFD supplemented with 1% quercetin for 16 weeks | reduces plasma cholesterol levels | mouse | [55] | [95] | ||||
| ↑ the expression of thermogenic genes in BAT (UCP1, PGC1α, FGF21) and genes coding for β-adrenergic receptors and AMPK | in vitro | [56] | [96] | ||||||
| Menthol | activates TRPM8 receptors | in vitro | [58] | [98] | |||||
| Cinnamaldehyde | ↑ UCP1 expression | in vitro | [57] | [97] | |||||
| Gingerol, Shogaol, 6-Paradol | activates TRPV1 channels | in vitro mouse |
[62]] | [102 | [63][ | ,103 | 64 | ,104] | |
| 100 mg/day of Kaempferia parviflora | ↑ energy expenditure in BAT-positive subjects; activates brown adipocytes | human | [67] | [107] |
BAT: brown adipose tissue; UCP1: uncoupling protein 1; WAT: white adipose tissue; TRPV1: transient receptor potential vanilloid 1; HSL: hormone-sensitive lipase; AMPK: adenosine monophosphate-activated protein kinase; PGC1α: peroxisome proliferator-activated receptor gamma coactivator 1-alpha; SIRT1: Sirtuin 1; FAS: fatty acid synthase; SREBP1: sterol response element-binding protein; LPL: lipoprotein lipase; PPAR: peroxisome proliferator-activated receptor; TRP: transient receptor potential; SNS: sympathetic nervous system; FGF21: fibroblast growth factor 21; TRPM8: transient receptor potential cation channel subfamily M member 8; ↑: increase, ↓ decrease.
References
- Minj, K.; Tsuyoshi, G.; Rina, Y. Fish oil intake induces UCP1 upregulation in brown and white adipose tissue via the sympathetic nervous system. Sci. Rep. 2016, 5, 186–197. Golden A. Obesity's Impact. Nurs Clin North Am. 2021 December; 4(56): 13-15.
- Matta, J.A.; Miyares, R.L.; Ahern, G.P.; Brothwell, S.L.C.; Barber, J.L.; Monaghan, D.T.; Jane, D.E.; Gibb, A.J.; Jones, S. TRPV1 is a novel target for omega-3 polyunsaturated fatty acids. J. Physiol. 2007, 578, 397–411. Blüher M. Obesity: global epidemiology and pathogenesis. Nat Rev Endocrinol. 2019 May; 5(15): 176-8.
- Ming, Z.; XIaoli, C. Eicosapentaenoic acid promotes thermogenic and fatty acid storage capacity in mouse subcutaneous adipocytes. Biochem. Biophys. Res. Commun. 2014, 4, 1446–1461. Safaei M, Sundararajan E, Driss M, Boulila W, Shapi A. A systematic literature review on obesity: Understanding the causes & consequences of obesity and reviewing various machine learning approaches used to predict obesity. Comput Biol Med. 2021 September;(136): 47-54.
- Tsuboyama, N.; Mayumi, K.; Hyounjulu, T. Up-Regulation of Liver Uncoupling Protein-2 mRNA by either Fish Oil Feeding or Fibrate Administration in Mice. Biochem. Biophys. Res. Commun. 2002, 3, 879–885. Navarro A, Villarroya J, Cereijo R, Giralt M, Villarroya F. The endocrine role of brown adipose tissue: An update on actors and actions. Rev Endocr Metab Disord. 2022 February; 1(23): 40-46.
- Takahashi, Y.; Ide, T. Dietary n-3 fatty acids affect mRNA level of brown adipose tissue uncoupling protein 1, and white adipose tissue leptin and glucose transporter 4 in the rat. Br. J. Nutr. 2000, 2, 175–184. Kuryłowicz A, Puzianowska-Kuźnicka M. Induction of Adipose Tissue Browning as a Strategy to Combat Obesity. Int J Mol Sci. 2020 August; 21(17): 6241.
- Oudart, H.; Groscolas, R.; Calgari, C.; Nibbelink, M.; Leray, C.; Le Maho, Y.; Malan, A. Brown fat thermogenesis in rats fed high-fat diets enriched with n-3 polyunsaturated fatty acids. Int. J. Obes. 1997, 21, 955–962. Armani A, Feraco A, Camajani E, Gorini G, Lombardo M, Caprio M. Nutraceuticals in Brown Adipose Tissue Activation. Cells. 2022 December; 11(24): 3996.
- Sadurskis, A.; Dicker, A.; Cannon, B.; Nedergaard, J. Polyunsaturated fatty acids recruit brown adipose tissue: Increased UCP content and NST capacity. Am. J. Physiol. Metab. 1995, 269, E351–E360. Martins F, Souza-Mello V, Aguila M, Mandarim-de-Lacerda C. Brown adipose tissue as an endocrine organ: updates on the emerging role of batokines. Horm Mol Biol Clin Investig. 2022 October; 10(15): 22-44.
- Bargut, T.C.L.; Silva-E-Silva, A.C.A.G.; Souza-Mello, V.; Mandarim-De-Lacerda, C.A.; Aguila, M.B. Mice fed fish oil diet and upregulation of brown adipose tissue thermogenic markers. Eur. J. Nutr. 2016, 55, 159–169. Negrea M, Neamtu B, Dobrotă I, Sofariu CCR, al. e. Causative Mechanisms of Childhood and Adolescent Obesity Leading to Adult Cardiometabolic Disease: A Literature Review. Appl. Sci.. 2021 December; 11(23): 1123-1156.
- Eckel, M.; Fleckenstein-Elsen, D.; Dennis, T. Eicosapentaenoic acid and arachidonic acid differentially regulate adipogenesis, acquisition of a brite phenotype and mitochondrial function in primary human adipocytes. Mol. Nutr. Food Res. 2016, 9, 2065–2075. Chait A, Den Hartigh L. Adipose Tissue Distribution, Inflammation and Its Metabolic Consequences, Including Diabetes and Cardiovascular Disease. Front Cardiovasc Med. 2020 February; 25(7): 22.
- Kim, J.; Okla, M.; Erickson, A.; Carr, T.; Natarajan, S.K.; Chung, S. Eicosapentaenoic Acid Potentiates Brown Thermogenesis through FFAR4-dependent Up-regulation of miR-30b and miR-378. J. Biol. Chem. 2016, 291, 20551–20562. Zhang F, Hao G, Shao M, Nham K, An Y, Wang Q, et al. An adipose tissue atlas: an image-guided identification of human-like bat and beige depots in rodents. Cell Metab.. 2018 October; 27(253): 252–62.
- Laiglesia, L.; Lorente-Cebrián, S.; Prieto-Hontoria, P.; Fernández-Galilea, M.; Ribeiro, S.; Sáinz, N.; Martínez, J.; Moreno-Aliaga, M. Eicosapentaenoic acid promotes mitochondrial biogenesis and beige-like features in subcutaneous adipocytes from overweight subjects. J. Nutr. Biochem. 2016, 37, 76–82. Srivastava S, Veech R. Brown and Brite: The Fat Soldiers in the Anti-obesity Fight. Front Physiol. 2019 January; 10(38): 33-89.
- Ludy, M.-J.; Moore, G.E.; Mattes, R.D. The Effects of Capsaicin and Capsiate on Energy Balance: Critical Review and Meta-analyses of Studies in Humans. Chem. Senses 2011, 37, 103–121. Gesta S, Tseng Y, Kahn R. Developmental origin of fat: tracking obesity to its source. Cell. 2007 October; 131(2): 242-256.
- Panchal, S.K.; Bliss, E.; Brown, L. Capsaicin in Metabolic Syndrome. Nutrients 2018, 10, 630. Long J, Svensson K, Tsai L, Zeng X, al e. A smooth muscle-like origin for beige adipocytes. Cell Metab. 2014 May; 5(19): 810-820.
- Baskaran, P.; Krishnan, V.; Ren, J.; Thyagarajan, B. Capsaicin induces browning of white adipose tissue and counters obesity by activating TRPV1 channel-dependent mechanisms. Br. J. Pharmacol. 2016, 173, 2369–2389. Rosenwald M, Wolfrum C. The origin and definition of brite versus white and classical brown adipocytes. Adipocyte. 2014 January; 1(3): 4-9.
- Yoneshiro, T.; Aita, S.; Matsushita, M.; Kayahara, T.; Kameya, T.; Kawai, Y.; Iwanaga, T.; Saito, M. Recruited brown adipose tissue as an antiobesity agent in humans. J. Clin. Investig. 2013, 123, 3404–3408. Bartness T, Vaughan H, Song C. Sympathetic and sensory innervation of brown adipose tissue. In J Obes. 2010 October; 1(10): 36-42.
- Sun, L.; Camps, S.G.; Goh, H.J.; Govindharajulu, P.; Schaefferkoetter, J.D.; Townsend, D.W.; Verma, S.K.; Velan, S.S.; Sun, L.; Sze, S.K.; et al. Capsinoids activate brown adipose tissue (BAT) with increased energy expenditure associated with subthreshold 18-fluorine fluorodeoxyglucose uptake in BAT-positive humans confirmed by positron emission tomography scan. Am. J. Clin. Nutr. 2018, 107, 62–70. Cinti S. La transdifferenziazione dell'organo adiposo.. Ancona: Istituto di Morfologia Umana, Università Politecnica delle Marche, Società Italiana dell'Obesità; ; 2008.
- Ohyama, K.; Nogusa, Y.; Shinoda, K.; Suzuki, K.; Bannai, M.; Kajimura, S. A synergistic anti-obesity effect by a combination of capsinoids and cold temperature through promoting beige adipocyte biogenesis. Diabetes 2016, 1, 1410–1423. Widmaier E, Raff H, Strang T, Vander J. Vander's human physiology : the mechanisms of body function. 14th ed. NY: New York McGraw-Hill Education; 2016.
- Maliszewska, K.; Kretowski, A. Brown Adipose Tissue and Its Role in Insulin and Glucose Homeostasis. Int. J. Mol. Sci. 2021, 22, 1530. Perez L, Perez L, Nene Y, Umpierrez G. Interventions associated with brown adipose tissue activation and the impact on energy expenditure and weight loss. Front. Endocrinol. 2022 December; 13.
- Schaik, L.; Kettle, C.; Green, R. Both caffeine and Capsicum annuum fruit powder lower blood glucose levels and increase brown adipose tissue temperature in healthy adult males. Front. Physiol. 2022, 13, 20–27. Van der Lans A, Hoeks J, Brans B. Cold acclimation recruits human brown fat and increases nonshivering thermogenesis. J Clin Invest. 2013 August; 8(123): 395-403.
- Gosselin, C.; Haman, F. Effects of green tea extracts on non-shivering thermogenesis during mild cold exposure in young men. Br. J. Nutr. 2012, 110, 282–288. Cannon B, Nedergaard J. Metabolic consequences of the presence or absence of the thermogenic capacity of brown adipose tissue in mice (and probably in humans). Int J Obes. 2010 October; 1(34): 7-16.
- Kurogi, M.; Kawai, Y.; Nagatomo, K.; Tateyama, M.; Kubo, Y.; Saitoh, O. Auto-oxidation Products of Epigallocatechin Gallate Activate TRPA1 and TRPV1 in Sensory Neurons. Chem. Senses 2014, 40, 27–46. Wankhade U, Shen M, Yadav H, Thakali K. Novel Browning Agents, Mechanisms, and Therapeutic Potentials of Brown Adipose Tissue. Biomed Res Int. 2016 December; 1(11): 65-69.
- Nomura, S.; Ichinose, T.; Jinde, M.; Kawashima, Y.; Tachiyashiki, K.; Imaizumi, K. Tea catechins enhance the mRNA expression of uncoupling protein 1 in rat brown adipose tissue. J. Nutr. Biochem. 2008, 19, 840–847. Morrison S, Madden C, Tupone D. Central control of brown adipose tissue thermogenesis. Front Endocrinol. 2012 January; 3(5): 33-39.
- Yan, J.; Zhao, Y.; Zhao, B. Green tea catechins prevent obesity through modulation of peroxisome proliferator-activated receptors. Sci. China Life Sci. 2013, 56, 804–810. Pereira R, McFarlane S. The Role of Brown Adipose Tissue in Cardiovascular Disease Protection: Current Evidence and Future Directions. Int J Clin Res Trials. 2019 September; 4(2): 136.
- Hursel, R.; Viechtbauer, W.; Dulloo, A.; Tremblay, A.; Tappy, L.; Rumpler, W.; Westerterp-Plantenga, M.S. The effects of catechin rich teas and caffeine on energy expenditure and fat oxidation: A meta-analysis. Obes. Rev. 2011, 12, e573–e581. Tseng Y, Cypess A, Kahn R. Cellular bioenergetics as a target for obesity therapy. Nat Rev Drug Discov. 2010 June; 9(6): 465-82.
- Basu, A.; Sanchez, K.; Leyva, M.J.; Wu, M.; Betts, N.M.; E Aston, C.; Lyons, T.J. Green Tea Supplementation Affects Body Weight, Lipids, and Lipid Peroxidation in Obese Subjects with Metabolic Syndrome. J. Am. Coll. Nutr. 2010, 29, 31–40. Preusch M, Baeuerle M, Albrecht C, Blessing E. GDF-15 protects from macrophage accumulation in a mousemodel of advanced atherosclerosis. Eur J Med Res. 2013 June; 18(1): 19.
- Van Schaik, L.; Kettle, C.; Green, R.; Irving, H.R.; Rathner, J.A. Effects of Caffeine on Brown Adipose Tissue Thermogenesis and Metabolic Homeostasis: A Review. Front. Neurosci. 2021, 15, 56–62. Stanford K, Middelbeek R, Townsend R, An D, Nygaard E. Brown adipose tissue regulates glucose homeostasis and insulin sensitivity. J Clin Invest. 2013 January; 123(1): 215-23.
- Thielecke, F.; Rahn, G.; Böhnke, J.; Adams, F.; Birkenfeld, A.L.; Jordan, J.; Boschmann, M. Epigallocatechin-3-gallate and postprandial fat oxidation in overweight/obese male volunteers: A pilot study. Eur. J. Clin. Nutr. 2010, 64, 704–713. Aldiss P, Davies G, Woods R, Budge H, Sacks H, Symonds M. 'Browning' the cardiac and peri-vascular adipose tissues to modulate cardiovascular risk. Int J Cardiol. 2017 February; 1(228): 265-274.
- Yoneshiro, T.; Matsushita, M.; Hibi, M.; Tone, H.; Takeshita, M.; Yasunaga, K.; Katsuragi, Y.; Kameya, T.; Sugie, H.; Saito, M. Tea catechin and caffeine activate brown adipose tissue and increase cold-induced thermogenic capacity in humans. Am. J. Clin. Nutr. 2017, 105, 873–881. Armani A, Feraco A, Camajani E, Gorini S. Nutraceuticals in Brown Adipose Tissue Activation. Cells. 2022 December;(24): 10-11.
- Akhtar, J.; Yar, M.S.; Grover, G.; Nath, R. Neurological and psychiatric management using COMT inhibitors: A review. Bioorganic Chem. 2019, 94, 103418. Sun L, Goh H, Verma S. Metabolic effects of brown fat in transitioning from hyperthyroidism to euthyroidism. Eur J Endocrinol. 2021 August; 4(185): 553-563.
- Di Pierro, F.; Bressan, A.; Ranaldi, D.; Rapacioli, G.; Giacomelli, L.; Bertuccioli, A. Potential role of bioavailable curcumin in weight loss and omental adipose tissue decrease: Preliminary data of a randomized, controlled trial in overweight people with metabolic syndrome. Preliminary study. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 4195–4202. Loeliger R, Maushart C, Gashi G. Relation of diet-induced thermogenesis to brown adipose tissue activity in healthy men. Am J Physiol Endocrinol Metab. 2021 January; 1(320): 93-101.
- Kim, S.W.; Choi, J.H.; Mukherjee, R.; Hwang, K.-C.; Yun, J.W. Proteomic identification of fat-browning markers in cultured white adipocytes treated with curcumin. Mol. Cell. Biochem. 2016, 415, 51–66. Wouter D, Vanhommerig J, Smulders N. Cold-activated brown adipose tissue in healthy men. N Engl J Med. 2013 April; 15(360): 1500-8.
- Nishikawa, S.; Kamiya, M.; Aoyama, H.; Nomura, M. Highly Dispersible and Bioavailable Curcumin but not Native Curcumin Induces Brown-Like Adipocyte Formation in Mice. Mol. Nutr. Food. Res. 2018, 5, 700–731. Labbè S, Mouchiroud M, Caron A, Secco B, E F. mTORC1 is Required for Brown Adipose Tissue Recruitment and Metabolic Adaptation to Cold. Sci Rep. 2016 November; 2(6): 33-37.
- Nishikawa, S.; Kamiya, M.; Aoyama, H.; Yoshimura, K.; Miyata, R.; Kumazawa, S.; Tsuda, T. Co-Administration of Curcumin and Artepillin C Induces Development of Brown-Like Adipocytes in Association with Local Norepinephrine Production by Alternatively Activated Macrophages in Mice. J. Nutr. Sci. Vitaminol. 2019, 65, 328–334. Van der Lans A, Hoeks J, Brans B, Vijgen G. Cold acclimation recruits human brown fat and increases nonshivering thermogenesis. J Clin Invest. 2013 August; 8(123): 3395-403.
- Saraf-Bank, S.; Ahmadi, A.; Paknahad, Z.; Maracy, M.; Nourian, M. Effects of curcumin on cardiovascular risk factors in obese and overweight adolescent girls: A randomized clinical trial. Sao Paulo Med. J. 2019, 137, 414–422. Singh R, Barrios A, Dirakvand G, Pervin S. Human Brown Adipose Tissue and Metabolic Health: Potential for Therapeutic Avenues. Cells. 2021 November;(5): 10-11.
- Glinjak, S.; Aebisher, D.; Bartusik, B. Health benefits of resveratrol administration. Acta Biochim. Pol. 2019, 1, 13–21. Orava J, Nuutila P, Noponen T, al. e. Blunted metabolic responses to cold and insulin stimulation in brown adipose tissue of obese humans. Obesity (Silver Spring). 2013 November; 21(11): 2279-87.
- Andrade, J.M.O.; Frade, A.C.M.; Guimarães, J.B.; Freitas, K.M.; Lopes, M.T.P.; Guimaraes, A.; De Paula, A.M.B.; Coimbra, C.C.; Santos, S.H.S. Resveratrol increases brown adipose tissue thermogenesis markers by increasing SIRT1 and energy expenditure and decreasing fat accumulation in adipose tissue of mice fed a standard diet. Eur. J. Nutr. 2014, 53, 1503–1510. Vosselman M, Van der Lans A, Brans B, Wierts R. Systemic β-adrenergic stimulation of thermogenesis is not accompanied by brown adipose tissue activity in humans. Diabetes. 2012 December; 12(61): 3106-13.
- Alberdi, G.; Rodríguez, V.M.; Miranda, J.; Macarulla, M.T.; Churruca, I.; Portillo, M.P. Thermogenesis is involved in the body-fat lowering effects of resveratrol in rats. Food Chem. 2013, 141, 1530–1535. Chitraju C, Fischer A, Farese R. Lipid Droplets in Brown Adipose Tissue Are Dispensable for Cold-Induced Thermogenesis. Cell Rep. 2020 November; 5(3): 108-34.
- Wang, S.; Liang, X.; Yang, Q.; Fu, X.; Rogers, C.J.; Zhu, M.; Rodgers, B.D.; Jiang, Q.; Dodson, M.V.; Du, M. Resveratrol induces brown-like adipocyte formation in white fat through activation of AMP-activated protein kinase (AMPK) α1. Int. J. Obes. 2015, 39, 967–976. Scheele C, Wolfrum C, al. e. Brown Adipose Crosstalk in Tissue Plasticity and Human Metabolism. Endocr Rev. 2020 January; 1(41): 53-65.
- Um, J.-H.; Park, S.-J.; Kang, H.; Yang, S.; Foretz, M.; McBurney, M.W.; Kim, M.K.; Viollet, B.; Chung, J.H. AMP-Activated Protein Kinase–Deficient Mice Are Resistant to the Metabolic Effects of Resveratrol. Diabetes 2010, 59, 554–563. Olza J, Sanchez G, Gil A, Ruiz J. Role of Exercise in the Activation of Brown Adipose Tissue. Annals of Nutrition and Metabolism. 2015;(67): 21-23.
- Milton-Laskíbar, I.; Gómez-Zorita, S.; Arias, N.; Romo-Miguel, N.; González, M.; Fernández-Quintela, A.; Portillo, M.P. Effects of resveratrol and its derivative pterostilbene on brown adipose tissue thermogenic activation and on white adipose tissue browning process. J. Physiol. Biochem. 2020, 76, 269–278. Ruiz , J , Tellez B, Delgado G, Aguilera C. Regulation of energy balance by brown adipose tissue: at least three potential roles for physical activity. British Journal of Sport Medicine. 2015 March;(10): 37-45.
- Scarano, F.; Gliozzi, M.; Zito, M.C.; Guarnieri, L.; Carresi, C.; Macrì, R.; Nucera, S.; Scicchitano, M.; Bosco, F.; Ruga, S.; et al. Potential of Nutraceutical Supplementation in the Modulation of White and Brown Fat Tissues in Obesity-Associated Disorders: Role of Inflammatory Signalling. Int. J. Mol. Sci. 2021, 22, 3351. Roberts L, O'Sullivan J, Schinzel R, Lewis G. β-Aminoisobutyric Acid Induces Browning of White Fat and Hepatic β-oxidation and is Inversely Correlated with Cardiometabolic Risk Factors. Cell Metab. 2014 January;(19): 96-108.
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. Bostrom P, Wu J, Jedrychowski P, Korde A, Ye L. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature. 2012; 4(81): 463- 468.
- Pinto-Garcia, L.; Efferth, T.; Torres, A.; Hoheisel, J.D.; Youns, M. Berberine Inhibits Cell Growth and Mediates Caspase-Independent Cell Death in Human Pancreatic Cancer Cells. Planta Medica 2010, 76, 1155–1161. Minj K, Tsuyoshi G, Rina Y. Fish oil intake induces UCP1 upregulation in brown and white adipose tissue via the sympathetic nervous system. 2015;. Scientific Reports. 2016 December; 5: 186-197.
- Zhang, Z.; Zhang, H.; Li, B.; Meng, X.; Wang, J.; Zhang, Y.; Yao, S.; Ma, Q.; Jin, L.; Yang, J.; et al. Berberine activates thermogenesis in white and brown adipose tissue. Nat. Commun. 2014, 5, 5493. Matta J, Miyares R, Ahern G. TRPV1 is a novel target for omega-3 polyunsaturated fatty acids. J. Physiol. 2007 January; 45(578): 397-411.
- van Dam, A.D.; Kooijman, S.; Schilperoort, M.; Rensen, P.C.; Boon, M.R. Regulation of brown fat by AMP-activated protein kinase. Trends Mol. Med. 2015, 21, 571–579. Ming Z, XIaoli C. Eicosapentaenoic acid promotes thermogenic and fatty acid storage capacity in mouse subcutaneous adipocytes. Biochemical and Biophysical Research Communications. 2014; 4(450): 1446-1461.
- Wu, L.; Xia, M.; Duan, Y.; Zhang, L.; Jiang, H.; Hu, X.; Yan, H.; Zhang, Y.; Gu, Y.; Shi, H.; et al. Berberine promotes the recruitment and activation of brown adipose tissue in mice and humans. Cell Death Dis. 2019, 10, 468. Tsuboyama N, Mayumi K, Hyounjulu T. Up-Regulation of Liver Uncoupling Protein-2 mRNA by either Fish Oil Feeding or Fibrate Administration in Mice. Biochemical and Biophysical Research Communications. 2002; 3(257): 879-885.
- Jiang, N.; Yang, M.; Han, Y.; Zhao, H.; Sun, L. PRDM16 Regulating Adipocyte Transformation and Thermogenesis: A Promising Therapeutic Target for Obesity and Diabetes. Front. Pharmacol. 2022, 13, 25–47. Takahashi Y, Ide T, al. e. Dietary n-3 fatty acids affect mRNA level of brown adipose tissue uncoupling protein 1, and white adipose tissue leptin and glucose transporter 4 in the rat. Br J Nutr. 2000 August; 2(84): 175-84.
- Oi-Kano, Y.; Iwasaki, Y.; Nakamura, T.; Watanabe, T.; Goto, T.; Kawada, T.; Watanabe, K.; Iwai, K. Oleuropein aglycone enhances UCP1 expression in brown adipose tissue in high-fat-diet-induced obese rats by activating β-adrenergic signaling. J. Nutr. Biochem. 2017, 40, 209–218. Oudart H, Groscolas R, Calgari C, Nibbelink M, Leray C. Brown fat thermogenesis in rats fed high-fat diets enriched with n-3 polyunsaturated fatty acids. Int J Obes Relat Metab Disord. 1997; 2(21): 955-962.
- Oi-Kano, Y.; Kawada, T.; Watanabe, T.; Koyama, F. Oleuropein, a phenolic compound in extra virgin olive oil, increases uncoupling protein 1 content in brown adipose tissue and enhances noradrenaline and adrenaline secretions in rats. J Nutr Sci Vitaminol 2018, 5, 363–370. Sadurskis A, Dicker A, Cannon B, Nedergaard J. Polyunsaturated fatty acids recruit brown adipose tissue: increased UCP content and NST capacity. Am J Physio. 1995; 3(269): 351- 360.
- Palmeri, R.; Monteleone, J.I.; Spagna, G.; Restuccia, C.; Raffaele, M.; Vanella, L.; Volti, G.L.; Barbagallo, I. Olive Leaf Extract from Sicilian Cultivar Reduced Lipid Accumulation by Inducing Thermogenic Pathway during Adipogenesis. Front. Pharmacol. 2016, 7, 1–10. Bargut T, Silva E, Souza-Mello V, Mandarim-de-Lacerda C. Mice fed fish oil diet and upregulation of brown adipose tissue thermogenic markers. Eur J Nutr. 2016 March; 2(55): 159-169.
- Pei, L.; Wan, T.; Wang, S.; Ye, M.; Qiu, Y.; Jiang, R.; Pang, N.; Huang, Y.; Zhou, Y.; Jiang, X.; et al. Cyanidin-3-O-β-glucoside regulates the activation and the secretion of adipokines from brown adipose tissue and alleviates diet induced fatty liver. Biomed. Pharmacother. 2018, 105, 625–632. Eckel M, Fleckenstein-Elsen D, Dennis T. Eicosapentaenoic acid and arachidonic acid differentially regulate adipogenesis, acquisition of a brite phenotype and mitochondrial function in primary human adipocytes. Molecular Nutrition & Food Research. 2016 May; 9(60): 2065-2075.
- Chen, K.; Kortesniemi, M.K.; Linderborg, K.M.; Yang, B. Anthocyanins as Promising Molecules Affecting Energy Homeostasis, Inflammation, and Gut Microbiota in Type 2 Diabetes with Special Reference to Impact of Acylation. J. Agric. Food Chem. 2022, 71, 1002–1017. Kim J, Okla M, Erickson A, Natarajan S, Chung S. Eicosapentaenoic acid potentiates brown thermogenesis through FFAR4-dependent up-regulation of miR-30b and miR-378. J Biol Chem.. 2016; 39(291): 20551-62.
- Zhu, Y.; Zhang, J.; Wei, Y.; Hao, J. The polyphenol-rich extract from chokeberry (Aronia melanocarpa L.) modulates gut microbiota and improves lipid metabolism in diet-induced obese rats. Nutr. Metab. 2020, 17, 50–54. Aguila T, Cristina L, Bargut A. Mice fed fish oil diet and upregulation of brown adipose tissue thermogenic markers. Eur J Nutr. 2016; 1(55): 159-169.
- Matsukawa, T.; Villareal, M.O.; Motojima, H.; Isoda, H. Increasing cAMP levels of preadipocytes by cyanidin-3-glucoside treatment induces the formation of beige phenotypes in 3T3-L1 adipocytes. J. Nutr. Biochem. 2017, 40, 77–85. Laiglesia L, Lorente-Cebrian S, Prieto-Hontoria P, Fernandez-Galilea M. Eicosapentaenoic acid promotes mitochondrial biogenesis and beige-like features in subcutaneous adipocytes from overweight subjects. J Nutr Biochem. 2016; 4(37): 76-82.
- Sheng, Y.; Liu, J.; Zheng, S.; Liang, F.; Luo, Y.; Huang, K.; Xu, W.; He, X. Mulberry leaves ameliorate obesity through enhancing brown adipose tissue activity and modulating gut microbiota. Food Funct. 2019, 10, 4771–4781. Ming Z, Xiaoli C. Eicosapentaenoic acid promotes thermogenic and fatty acid storage capacity in mouse subcutaneous adipocytes. Biochem Biophys Res Commun. 2014 August; 4(450): 1446-51.
- Lee, M.; My, L. The Effects of C3G and D3G Anthocyanin-Rich Black Soybean on Energy Metabolism in Beige-like Adipocytes. J. Agric Food Chem. 2020, 43, 12011–12018. Lud M, Moore G, Mattes R. The effects of capsaicin and capsiate on energy balance: critical review and meta-analyses of studies in humans. Chem Senses. 2012; 1(37): 103-121.
- Pei, Y.; Otieno, D.; Gu, I.; Lee, S.-O.; Parks, J.S.; Schimmel, K.; Kang, H.W. Effect of quercetin on nonshivering thermogenesis of brown adipose tissue in high-fat diet-induced obese mice. J. Nutr. Biochem. 2020, 88, 108532. Pancha S, Bliss E, Brown L. Capsaicin in Metabolic Syndrome. Nutrients. 2018; 5(10): 630.
- Gil Lee, S.; Parks, J.S.; Kang, H.W. Quercetin, a functional compound of onion peel, remodels white adipocytes to brown-like adipocytes. J. Nutr. Biochem. 2017, 42, 62–71. Baskaran P, Krishnan V, Ren J, Thyagarajan B. Capsaicin induces browning of white adipose tissue and counters obesity by activating TRPV1 channel-dependent mechanisms. Br J Pharmacol. 2017 May; 15(173): 2369-89.
- Typolt, O.; Filingeri, D. Evidence for the involvement of peripheral cold-sensitive TRPM8 channels in human cutaneous hygrosensation. Am. J. Physiol. Integr. Comp. Physiol. 2020, 318, R579–R589. Yoneshiro T, Aita S, Matsushita M, Kayahara T, Kameya T, Kawai Y, et al. Recruited brown adipose tissue as an antiobesity agent in humans. J Clin Invest. 2013; 7(123): 3404-.
- Vizin, R.C.L.; Motzko-Soares, A.C.P.; Armentano, G.M.; Ishikawa, D.T.; Cruz-Neto, A.P.; Carrettiero, D.C.; Almeida, M.C.; Motzko-Soares, A.C.P. Short-term menthol treatment promotes persistent thermogenesis without induction of compensatory food consumption in Wistar rats: Implications for obesity control. J. Appl. Physiol. 2018, 124, 672–683. Camps S, Sun L, Goh H, Govindharajulu P. Capsinoids activate brown adipose tissue (BAT) with increased energy expenditure associated with subthreshold 18-fluorine fluorodeoxyglucose uptake in BAT-positive humans confirmed by positron emission tomography scan. Am J Clin Nutr. 2018 January; 1(107): 62-70.
- Jiang, J.; Emont, M.; Jun, H.; Qiao, X.; Liao, J.; Kim, D.-I.; Wu, J. Cinnamaldehyde induces fat cell-autonomous thermogenesis and metabolic reprogramming. Metabolism 2017, 77, 58–64. Ohyama K, Nogusa Y, Shinoda K, Suzuki K, Bannai M, Kajimura S. A synergistic anti-obesity effect by a combination of capsinoids and cold temperature through promoting beige adipocyte biogenesis. Diabetes. 2016; 1(65): 1410-23.
- Young, Y.; Eun, J.; Kima, D. Thiacremonone, a sulfur compound isolated from garlic, attenuates lipid accumulation partially mediated via AMPK activation in 3T3-L1 adipocytes. J. Nutr. Biochem. 2012, 12, 1552–1558. Maliszewska K, Kretowski A. Brown Adipose Tissue and Its Role in Insulin and Glucose Homeostasis. Int J Mol Sci. 2021 February; 4(22): 15-30.
- Kaur, J.; Singh, D.P.; Kumar, V.; Kaur, S.; Bhunia, R.K.; Kondepudi, K.K.; Kuhad, A.; Bishnoi, M. Transient Receptor Potential (TRP) based polypharmacological combination stimulates energy expending phenotype to reverse HFD-induced obesity in mice. Life Sci. 2023, 12, 1704. Schaik L, Kettle C, Green R. Both caffeine and Capsicum annuum fruit powder lower blood glucose levels and increase brown adipose tissue temperature in healthy adult males. Front. Physiol. 2022 August; 13(12): 20-27.
- Deng, X.; Zhang, S.; Wu, J.; Sun, X. Promotion of Mitochondrial Biogenesis via Activation of AMPK-PGC1α Signaling Pathway by Ginger (Zingiber officinale Roscoe) Extract, and Its Major Active Component 6-Gingerol. J. Food Sci. 2019, 84, 2101–2111. Gosselin C, Haman F. Effects of green tea extracts on non-shivering thermogenesis during mild cold exposure in young men. Br J Nutr. 2016 March; 2(110): 282–8.
- Konstantinidi, M.; Koutelidakis, A.E. Functional Foods and Bioactive Compounds: A Review of Its Possible Role on Weight Management and Obesity’s Metabolic Consequences. Medicines 2019, 6, 94. Kurogi M, Kawai Y, Nagatomo K, Tateyama M, Kubo Y. Auto-oxidation products of epigallocatechin gallate activate TRPA1 and TRPV1 in sensory neurons. Chem Senses. 2015 August; 1(40): 27-46.
- Hattori, H.; Yamauchi, K.; Onwona-Agyeman, S.; Mitsunaga, T. Identification of vanilloid compounds in grains of paradise and their effects on sympathetic nerve activity. J. Sci. Food Agric. 2018, 98, 4742–4748. Nomura S, Ichinose T, Jinde M, Kawashima Y, Tachiyashi K. Tea catechins enhance the mRNA expression of uncoupling protein 1 in rat brown adipose tissue. Nutr Biochem. 2008 April; 1(19): 840-7.
- Matsushita, M.; Yoneshiro, T.; Aita, S.; Kamiya, T.; Kusaba, N.; Yamaguchi, K.; Takagaki, K.; Kameya, T.; Sugie, H.; Saito, M. Kaempferia parviflora Extract Increases Whole-Body Energy Expenditure in Humans: Roles of Brown Adipose Tissue. J. Nutr. Sci. Vitaminol. 2015, 61, 79–83. Ya J, Zhao Y, Zhao B. Green tea catechins prevent obesity through modulation of peroxisome proliferator-activated receptors. Sci China Life Sci. 2013; 4(56): 804–10.
- Scheele, C.; Wolfrum, C. Brown Adipose Crosstalk in Tissue Plasticity and Human Metabolism. Endocr. Rev. 2019, 41, 53–65. Hursel R, Viechtbauer W, Dulloo A, Tremblay A, Tappy L. The effects of catechin rich teas and caffeine on energy expenditure and fat oxidation: a meta-analysis. Obes Rev. 2011 December; 3(12): 573-81.
- Sanchez-Delgado, G.; Martinez-Tellez, B.; Olza, J.; Aguilera, C.M.; Gil, Á.; Ruiz, J.R. Role of Exercise in the Activation of Brown Adipose Tissue. Ann. Nutr. Metab. 2015, 67, 21–32. Basu A, Sanchez K, Leyva M, Wu M, Betts C. Green Tea Supplementation Affects Body Weight, Lipids, and Lipid Peroxidation in Obese Subjects with Metabolic Syndrome. Journal of the American College of Nutrition. 2010; 1(29): 31-40.
- Schaik L, Kettle C, Green R, Irving H. Effects of Caffeine on Brown Adipose Tissue Thermogenesis and Metabolic Homeostasis. Front. Neurosci. 2021 February; 15: 56-62.
- Thielecke F, Rahn G, Bohnke J, Adams F, Birkenfeld A. Epigallocatechin-3-gallate and postprandial fat oxidation in overweight/obese male volunteers: a pilot study. Eur J Clin Nutr. 2010 May; 2(64): 704–13.
- Yoneshiro T, Matsushita M, Hibi M, Tone H. Tea catechin and caffeine activate brown adipose tissue and increase cold-induced thermogenic capacity in humans. Am J Clin Nutr. 2017 January; 3(105): 873-81.
- Akhtar M, Yar M, Grover G, Nath R. Neurological and psychiatric management using COMT inhibitors: A review. Bioorg Chem. 2020 October; 1(94): 410-18.
- Di Pierro F, Bressan A, Ranaldi D, Rapacioli G GLBA. Potential role of bioavailable curcumin in weight loss and omental adipose tissue decrease: preliminary data of a randomized, controlled trial in overweight people with metabolic syndrome. Preliminary study. Eur Rev Med Pharmacol Sci. 2015; 1(19): 4195–202.
- Kim S, Choi J, Mukherjee R, Hwang K. Proteomic identification of fat-browning markers in cultured white adipocytes treated with curcumin. ;(415): 51-66. Mol Cell Biochem. 2016 March; 2(415): 51-66.
- Nishikawa S, Kamiya M, Aoyama H, Nomura M. Highly Dispersible and Bioavailable Curcumin but not Native Curcumin Induces Brown-Like Adipocyte Formation in Mice. Mol Nutr Food Res. 2018 Dec; 5(10): 700-731.
- Nishikawa S, Aoyama H, Yoshimura K, Miyata R. Co-Administration of Curcumin and Artepillin C Induces Development of Brown-Like Adipocytes in Association with Local Norepinephrine Production by Alternatively Activated Macrophages in Mice. Nutr Sci Vitaminol. 2019 January; 5(64): 328-334.
- Saraf-Bank S, Ahmadi A, Paknahad Z, Maracy M, Nourian M. Effects of curcumin on cardiovascular risk factors in obese and overweight adolescent girls: a randomized clinical trial. Sao Paulo Med. J. 2019 October; 5(137): 14-2.
- Glinjak S, Aebisher D, Bartusik B. Health benefits of resveratrol administration. Acta Biochim Pol. 2019 February; 1(66): 13-21.
- Andrade J, Frade A, Guimaraes J, Freitas K. Resveratrol increases brown adipose tissue thermogenesis markers by increasing SIRT1 and energy expenditure and decreasing fat accumulation in adipose tissue of mice fed a standard diet. Eur J Nutr. 2014;(53):. Eur J Nutr. 2014; 4(53): 1503-10.
- Alberdi G, Rodriguez V, Miranda J, Macarulla M. Thermogenesis is involved in the body- fat lowering effects of resveratrol in rats. Food Chem. 2013 November; 1(141): 1530-35.
- Wang S, Liang X, Yang Q, Fu X, Rogers C, Zhu M, et al. Resveratrol induces brown-like adipocyte formation in white fat through activation of AMP-activated protein kinase (AMPK) alpha1. Int J Obes. 2015; 3(39): 967-76.
- Um J, Park S, Kang H, Yang S, Foretz M, McBurney M. AMP-activated protein kinase- deficient mice are resistant to the metabolic effects of resveratrol. Diabetes. 2010 October; 4(59): 554-63.
- Laskíbar M, Gómez-Zorita S, Romo-Miguel N, Arias N. Effects of resveratrol and its derivative pterostilbene on brown adipose tissue thermogenic activation and on white adipose tissue browning process. J Physiol Biochem. 2020 May; 2(76): 269-278.
- Scarano f, Gliozzi M, Zito M, Guarnieri L. Potential of Nutraceutical Supplementation in the Modulation of White and Brown Fat Tissues in Obesity-Associated Disorders: Role of Inflammatory Signalling. Int J Mol Sci. 2021 March; 22(7): 33-51.
- Manach C, Williamson G, Morand C, Scalbert A. Bioavailability and bioefficacy of polyphenols in humans. Review of 97 bioavailability studies. Am J Clin Nutr. 2005 March; 1(81): 1230-42.
- Pinto-Garcia L, Efferth T, Torres A, Hoheisel J, Youns M. Berberine Inhibits Cell Growth and Mediates Caspase-Independent Cell Death in Human Pancreatic Cancer Cells. Planta Medica. 2010; 11(76): 1155-61.
- Zhang Z, Zhang H, Li B, Meng X, Wang J, Zhang Y, et al. Berberine activates thermogenesis in white and brown adipose tissue. Nat Commun. 2014 October; 1(5): 5493.
- Van Dam A, Kooijman S, Schilperoort M, Rensen P, Boon M. Regulation of brown fat by AMP-activated protein kinase. Trends Mol Med. 2015 September; 9(21): 571–9.
- Wu L, Xia M, Duan Y, Zhang L, Jiang H, Hu X. Berberine promotes the recruitment and activation of brown adipose tissue in mice and humans. Cell Death Dis. 2019 June; 10(468): 706-19.
- Jiang N, Yang M, Han Y, Zhao H. PRDM16 Regulating Adipocyte Transformation and Thermogenesis: A Promising Therapeutic Target for Obesity and Diabetes. Front Pharmacol. 2022 April; 13(87): 25-47.
- Oi-Kano W, Iwasaki Y, Nakamura T, Watanabe T. Oleuropein aglycone enhances UCP1 expression in brown adipose tissue in high-fat-diet-induced obese rats by activating β- adrenergic signaling.. J Nutr Biochem. 2017 February; 9(40): 209-218.
- Oi-Kano Y, Kawada T, Watanabe T, Koyama F. Oleuropein, a phenolic compound in extra virgin olive oil, increases uncoupling protein 1 content in brown adipose tissue and enhances noradrenaline and adrenaline secretions in rats. J Nutr Sci Vitaminol. 2018 October; 5(54): 363-70.
- Palmeri R, Monteleone J, Restuccia C, Raffaele M. Olive Leaf Extract from Sicilian Cultivar Reduced Lipid Accumulation by Inducing Thermogenic Pathway during Adipogenesis. Front Pharmacol. 2016 May; 7(143).
- Pei L, Wan T, Wang S, Ye M, Qiu Y, Jiang R, et al. Cyanidin-3-O-β-glucoside regulates the activation and the secretion of adipokines from brown adipose tissue and alleviates diet induced fatty liver. Biomed Pharmacother. 2018 September; 105: 625-632.
- Chen K, Kortesniemi M, Linderborg K, Yang B. Anthocyanins as Promising Molecules Affecting Energy Homeostasis, Inflammation, and Gut Microbiota in Type 2 Diabetes with Special Reference to Impact of Acylation. J Agric Food Chem. 2023 January; 71(2): 1002-1017.
- Zhu Y, Zhang J, Wei Y, Hao J. The polyphenol-rich extract from chokeberry (Aronia melanocarpa L.) modulates gut microbiota and improves lipid metabolism in diet-induced obese rats. Nutr Metab. 2020 July; 17: 50-54.
- Matsukawa T, Villareal M, Motojima O, Isoda H. Increasing cAMP levels of preadipocytes by cyanidin-3-glucoside treatment induces the formation of beige phenotypes in 3T3-L1 adipocytes. J Nutr Biochem. 2017 February; 18(40): 77-85.
- Liu J, Sheng Y, Zheng L, Liang F, Luo Y. Mulberry leaves ameliorate obesity through enhancing brown adipose tissue activity and modulating gut microbiota. Food Funct. 2019; 8(10): 4771-4781. Food Funct.. 2019 August; 8(10): 4771-4781.
- Lee M, My L. The Effects of C3G and D3G Anthocyanin-Rich Black Soybean on Energy Metabolism in Beige-like Adipocytes. J Agric Food Chem. 2020 October; 43(68): 12011-12018.
- Pei Y, Otieno D, Gu I, Lee S, Parks J. Effect of quercetin on nonshivering thermogenesis of brown adipose tissue in high-fat diet-induced obese mice. J Nutr Biochem.. 2021 February;(88): 108- 116.
- Lee S, Parks J, Who Kang H. Quercetin, a functional compound of onion peel, remodels white adipocytes to brown-like adipocytes. J Nutr Biochem. 2017 April;(42): 62-71.
- Otieno D PWY,GILS. Effect of quercetin on nonshivering thermogenesis of brown adipose tissue in high-fat diet-induced obese mice. J Nutr Biochem. 2021 February; 10(88): 10-16.
- Typolt O, Filingeri D. Evidence for the involvement of peripheral cold-sensitive TRPM8 channels in human cutaneous hygrosensation. Am J Physiol Regul Integr Comp Physiol. 2020 March; 3(318): 579-589.
- Vizin R, Motzko-Soares A, Armentano G, Ishikawa D, Cruz-Neto A, al. e. Short-term menthol treatment promotes persistent thermogenesis without induction of compensatory food consumption in Wistar rats: implications for obesity control. J Appl Physiol. 2018 March; 3(124): 672-683.
- Jiang J, Emont M, Jun H, Qiao X. Cinnamaldehyde induces fat cell-autonomous thermogenesis and metabolic reprogramming. Metabolism. 2017 December; 77: 59-64.
- Young Y, Eun J, Kima D. Thiacremonone, a sulfur compound isolated from garlic, attenuates lipid accumulation partially mediated via AMPK activation in 3T3-L1 adipocytes. The Journal of Nutritional Biochemistry. 2012 December; 12(23): 1552-1558.
- Kaur J, Singh D, Kumar V, Kaur S, Bhunia R. Transient Receptor Potential (TRP) based polypharmacological combination stimulates energy expending phenotype to reverse HFD-induced obesity in mice. Life Sci. 2023 April; 12: 1704.
- Deng X, Zhang S, Wu J, Sun X. Promotion of Mitochondrial Biogenesis via Activation of AMPK-PGC1ɑ Signaling Pathway by Ginger (Zingiber officinale Roscoe) Extract, and Its Major Active Component 6-Gingerol. J Food Sci. 2019 August; 84(8): 2101-2111.
- Konstantinidi M, Koutelidakis A. Functional Foods and Bioactive Compounds: A Review of Its Possible Role on Weight Management and Obesity's Metabolic Consequences. Medicines (Basel). 2019 September; 6(3): 94.
- Hattori H, Yamauchi K, Onwona-Agyeman S, Mitsunaga T. Identification of vanilloid compounds in grains of paradise and their effects on sympathetic nerve activity. J Sci Food Agric. 2018 September; 98(12): 4742-4748.
- Matsushita M, Yoneshiro T, Aita S. Kaempferia parviflora Extract Increases Whole-Body Energy Expenditure in Humans: Roles of Brown Adipose Tissue. Journal of Nutritional Science and Vitaminol. 2015 May; 1(61): 79-83.
- Saito M, Okamatsu-Ogura Y, Matsushita M, Watanabe K. Human brown adipose tissue: regulation and anti-obesity potential. Endocrine Journal. 2014 May; 2(13): 15-27.
- Poekes L, Lanthier N, Leclercq I. Brown adipose tissue: a potential target in the fight against obesity and the metabolic syndrome. Clin Sci (Lond). 2015 Dec; 11(129): 933–949.
- Wang X, Xu M, Li Y. Adipose Tissue Aging and Metabolic Disorder, and the Impact of Nutritional Interventions. Nutrients. 2022 July; 14(15): 31-34.
- Wu J, Cohen P, Spiegelman B. Adaptive thermogenesis in adipocytes: is beige the new brown? Genes Dev. 2013 February; 27(3): 234-50.
- Machado S, Pasquarelli-do-Nascimento G, Santos da Silva D, al e. Browning of the white adipose tissue regulation: new insights into nutritional and metabolic relevance in health and diseases. Nutr Metab (Lond). 2022 September; 19(1): 22-69.
- Pilkington A, Paz H, Wankhade U. Beige Adipose Tissue Identification and Marker Specificity. Front Endocrinol. 2021 March; 12.
- Wu J, Cohen P, Spiegelman B. Adaptive thermogenesis in adipocytes: is beige the new brown? Genes Dev. 2013 February; 3(27): 234-50.
- Rajakumari S, Wu J, Ishibashi J, Lim H, Giang A. EBF2 Determines and Maintains Brown Adipocyte Identit. Cell Metab.. 2013 December;(17): 562–574.
- Seale P, Bjork B, Yang W, Kajimura S, Chin S, Kuang S. PRDM16 controls a brown fat/skeletal muscle switch. Nature. 2018 October; 454: 961–967.
- Seale P, Kajimura S, Spiegelman B. Transcriptional control of brown adipocyte development and physiological function-of mice and men. Genes Dev. 2019 March; 23: 788- 797.
- Altınova A. Beige Adipocyte as the Flame of White Adipose Tissue: Regulation of Browning and Impact of Obesity. J Clin Endocrinol Metab. 2022 April; 5(105): 1778-1788.
- Mohamed-Ali V, Pinkney J, Coppack S. Adipose tissue as an endocrine and paracrine organ. International Journal of Obesity. 1998; 22: 1145-1158.
- Sanchez-Gurmaches J, Guertin D. Adipocytes arise from multiple lineages that are heterogeneously and dynamically distributed. Nature Communications. 2014 June; 5(4099): 18-30.
- Pošćić N, Montanari T, Colitti M. Factors involved in white-to-brown adipose tissue conversion and in thermogenesis: a review. Obese Rev. 2017 May; 5(18): 495-513.
- Baker M, Gaukrodger N, Bongani M, Imrie H. Association between common polymorphisms of the proopiomelanocortin gene and body fat distribution: a family study. Diabetes. 2005 August; 8(54): 2492-6.
- Navarro A, Villarroya J, Cereijo R, Giralt M, Villarroya F. The endocrine role of brown adipose tissue: An update on actors and actions. Endocrine and Metabolic Disorders. 2021 March; 23(1): 31-41.
More