Chytridiomycota (zoosporic true fungi) have a consistent presence in soils and have been frequently identified within many diverse terrestrial environments. However, Chytridiomycota and other early-diverging fungi have low representation in whole-genome sequencing databases compared to Dikarya. New molecular techniques have provided insights into the diversity and abundance of chytrids in soils and the changes in their populations both spatially and temporally. Chytrids complete their life cycle within rapidly changing soil environments where they may be more common within micropores due to protection from predation, desiccation, and extreme temperatures. Reproductive and morphological changes occur in response to environmental changes including pH, fluctuating nutrient concentrations, and metals at levels above toxic thresholds. Rhizoids share some features of hyphae, including the spatial regulation of branching and the ability to attach, adapt to, and proliferate in different substrates, albeit on a microscale. Soil chytrids provide a pool of novel enzymes and proteins which enable a range of lifestyles as saprotrophs or parasites, but also can be utilised as alternative tools with some biotechnological applications. Thus, 3D live-cell imaging and micromodels such as MicroCT may provide insight into zoospore functions and rhizoid plasticity, respectively, in response to various conditions. A combination of classical techniques of soil chytrid baiting with simultaneous molecular and ecological data will provide insights into temporal population changes in response to environmental change. The authors emphasise the need to review and improve DNA-based methodologies for identifying and quantifying chytrids within the soil microbiome to expand our knowledge of their taxonomy, abundance, diversity, and functionality within soil environments.
Chytridiomycota have been isolated from many environments, studied, and described
[1][2][1,2]; however, their ecological importance within soil environments has been neglected. Despite this, it is apparent that these fungi play a keystone role in many aquatic environments
[3]. The taxonomic classification and organisation of the phylogenetic tree of the fungal kingdom is under constant review and evolution
[4][5][6][7][4,5,6,7]. In particular, significant caveats in phylogenetic and evolutionary knowledge concerning basal lineages of fungi still exist. Early classification of zoosporic true fungi (chytrids) combined the group of fungus-like species within the aquatic phycomycetes (
sensu Sparrow 1960)
[1]. Further revision of the taxonomy of eucaryotic organisms assigned chytrids to the supergroup Opisthokonta
[8][9][8,9]. Later, zoosporic true fungi were split from the fungus-like zoosporic members of the Opisthokonta, with chytrids being assigned to the kingdom Fungi
[10]. Chytridiomycota (chytrids), Neocallimastigomycota (neocallimastigos), and Monoblepharidomycota (monoblephs) are zoosporic early-diverging phyla under the sub-kingdom Chytridiomyceta
[11][12][11,12] (
Figure 1), although the exact relationships between these phyla are still to be determined
[7].
Within Chytridiomycota there are currently 14 described orders: Caulochytriales, Chytridiales, Cladochytriales, Gromochytriales, Lobulomycetales, Mesochytriales, Polychytriales, Polyphagales, Rhizophydiales, Rhizophlyctidales, Spizellomycetales, Synchytriales, Zygophlyctidales, and Zygorhizidiales
[7]. However, more than half of these orders still lack sequence data. Nephridiophagids have also been recently shown to sit within the Chytridiomycota phylum
[13] and may form an additional order. For the anaerobic fungi phylum Neocallimastigomycota, there is only one order, Neocallimastigales. For the Monoblepheridiomycota there is also one order, Monoblepheridales. Previously, Blastocladiomycota was grouped within Chytridiomyceta but is now recognised as phylogenetically distinct based on morphological and molecular analyses
[14][15][16][17][14,15,16,17]. However, the position of Blastocladiomycota on the fungal evolutionary tree has not yet been resolved
[7]. More work is needed to fully understand the taxonomy of early-diverging fungi. Recently, increased phylogenetic diversity within the early-diverging taxa
[7][11][7,11] has further highlighted the need to resolve these relationships, including within Chytridiomyceta. This review focuses on Chytridiomycota.
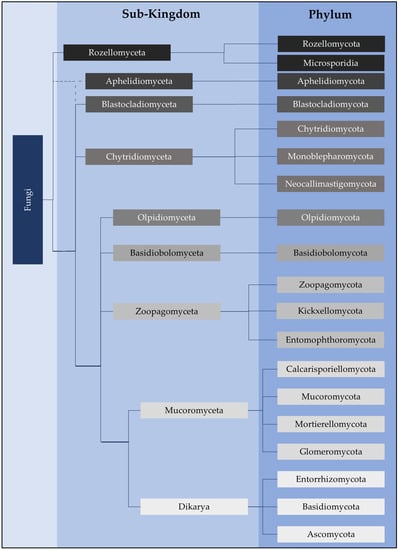
Figure 1. Fungi classification at the higher taxonomic ranks of sub-kingdom and phylum as per Powell and Letcher 2014
[12], Tedersoo et al. (18S and 28S rRNA sequence data) (2018)
[11], and James et al. (2020)
[7]. Although our current understanding of classifications has been updated, James et al. (2006a, 2006b)
[15][18][15,18] has provided foundational phylogenetic analyses using the 18S, 28S, 5.8S, ITS, EF1α, RPB1 and RPB2 markers. The exact branching of Aphelidomyceta is still unclear (dotted lines).
The morphological and physiological characteristics which allow chytrids to grow and disperse in soil environments are distinct and have been previously discussed
[19]. The characteristic features of chytrids include chemotactic responses
[20][21][20,21], motile zoospores propelled by a posterior flagellum
[1][22][1,22], the presence of cholesterol rather than ergosterol as the major sterol
[23], and morphological adaptations to environmental cues at different growth stages of the life cycle (
Figure 2). These adaptations include changes in zoospore size, shape, and release during zoosporulation, changes in the size of the sporangium, and changes in the length and branching of the rhizoids after encystment
[19] (
Figure 3). A further feature is the ability of the rhizoid to attach to and penetrate highly resistant solid substrates, such as the hard outer casings of diatoms
[24] and pollen grains
[25] (
Figure 3e). Stress tolerance and ruderal ecological strategies may both be adopted by individual chytrid species
[19] in order to survive, grow, and complete their life cycle under a wide range of environmental conditions. There are a number of species in Chytridiomycota that are pathogenic in aquatic environments.
Batrachochytrium dendrobatidis (order Rhizophydiales) can be considered one of the most important infectious diseases impacting the biodiversity of the amphibian population
[26][27][28][29][30][26,27,28,29,30]. Members of Chytridiomycota have been associated with phytoplankton, including diatoms, dinoflagellates, and cyanobacteria, as microparasites
[31][32][33][34][31,32,33,34]. They are well documented in bloom events in lakes
[35][36][35,36] and algal blooms in the Mediterranean Sea
[37]. Data indicate that fungal parasites can potentially control the fate of phytoplankton-derived organic matter by enhancing remineralisation and reducing sedimentation in freshwater and coastal systems
[31].
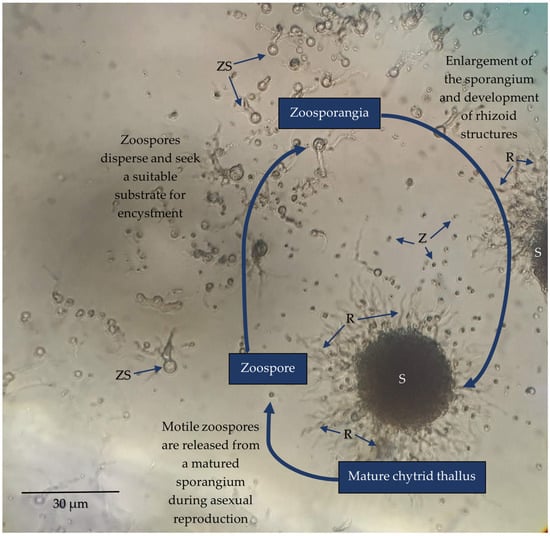
Figure 2. Light micrograph of Spizellomyces sp. (order Spizellomycetales) culture capturing three characteristic stages of development; a mature thallus containing a large sporangium (S) and rhizoid (R) extensions (not all chytrid species produce rhizoids), sporulation to disperse motile zoospores (Z), and zoospore encystment which initiates maturation from a zoosporangium (ZS) into a mature thallus. Strains were revived from long-term stock cultures by aseptically removing a segment of agar containing mature sporangium and placing it inoculated side down onto fresh peptone, yeast, and glucose (PYG) medium (2.5 mM peptone, 4.56 mM yeast extract, 27.7 mM glucose, 2% w/v agar). A few drops of sterile deionised water were then added over the inoculated agar to promote sporulation. Cultures were incubated at room temperature over the course of a week to observe the chytrid life cycle.
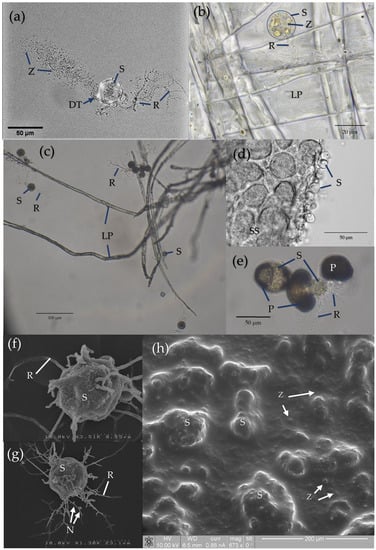
Figure 3. Microscopic images of Chytridiomycota (
a) captured with a confocal microscope, (
b–
e) taken using a light microscope, and (
f–
h) captured via scanning electron microscopy (SEM). As per
Figure 2, strains were cultured and maintained on PYG. (
a)
Spizellomyces sp. (order Spizellomycetales) chytrid during sporulation, identifying thallus structures including the sporangium (S), rhizoids (R), discharge tube (DT), and released zoospores (Z).
Spizellomyces sp. was cultured and prepared for microscopy as per Hanrahan-Tan et al. (2019)
[38]. (
b)
Rhizophlyctis rosea (order Rhizophlyctidales) attached to lens paper (LP) as a cellulose substrate. Adapted with permission from Henderson et al. (2019)
[39]. Copyright 2019 Nova Hedwigia. (
c)
Spizellomyces sp. attached to lens paper as a cellulose substrate. Culture plates were flooded with sterile deionised water from which an aliquot of active zoospores could be taken approximately 2 h later. The aliquot was then used to inoculate a liquid medium containing autoclaved lens paper baits (1.5 mm diameter). (
d)
Terramyces sp. (order Rhizophydiales) attached to snake skin (SS) as a keratin substrate. (
e)
Spizellomyces sp. thalli attached to and penetrating the tough exine layer of pollen grains (P) to access nutrients. As per (c), an inoculum of fresh zoospores was added to liquid medium containing autoclaved pollen grains. (
f)
R. rosea thallus under normal conditions on PYG prepared as per Henderson et al. (2017, 2019)
[39][40][39,40]. Adapted with permission from Henderson et al. (2017, 2019)
[39][40][39,40] and reproduced with permission from The Licensor through PLSclear. Copyright 2017 Taylor and Francis Group LLC (Books) US and copyright 2019 Nova Hedwigia. (
g)
R. rosea thallus. Incubation on media containing 60 ppm copper over 5 days shortens the length of rhizoids, increases the degree of rhizoid branching, and can lead to nodulation (N). Image sourced from Henderson et al. (2017)
[40]. Reproduced with permission from The Licensor through PLSclear. Copyright 2017 Taylor and Francis Group LLC (Books) US. (
h) Environment SEM of
Terramyces sp. colony grown on PYG medium. Sporangia appear as larger bumps (>20 µm) under the surface of the biofilm while zoospores are encysted as small spots (<10 µm) in the biofilm surface.
This review examines the recent evidence for the presence and relative abundance of chytrids in different soil environments. The authors examine the soil environment and the functions and adaptive strategies of soil chytrid populations, particularly in light of the recent additional knowledge on their morphology and functionality. The authors then highlight the current research discussing potential applications in biotechnology. Finally, the authors recommend innovative methods for further investigating the distribution and ecology of chytrids in soil environments.