 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Renuka Attanayake | + 3461 word(s) | 3461 | 2021-02-26 04:20:14 | | | |
| 2 | Peter Tang | Meta information modification | 3461 | 2021-03-07 08:47:37 | | |
Video Upload Options
Anaerobic soil disinfection (ASD) has been identified as an alternative soil-borne pathogen control strategy to chemical fumigation. ASD involves the application of an easily liable carbon source followed by irrigation to field capacity and maintenance of an anaerobic condition for a certain period.
1. Introduction
Crops are often attacked by various plant pathogens, plant-parasitic nematodes, insect pests, and weeds causing great economic losses around the world. Among diverse groups of plant pathogens, soil-borne phytopathogens pose a great threat to crop production [1][2][3]. Although soil is a home for billions of living organisms (both macro and microorganisms), they must face a multitude of challenges such as flood, drought, and agricultural practices. However, soil-borne pathogens can survive under these challenges and cause serious crop damage around the world. For example, waterlogged agricultural fields may be unfavourable for many organisms but favourable for root-infecting fungi and oomycetes such as Pythium and Phytophthora spp. [4][5][6]. Although drought conditions are unfavourable for most of the organisms, soil-borne pathogen species such as Fusarium spp. and Verticillium spp. [5] manage to cause severe infections. Hence, soil-borne phytopathogens show a great deal of evolutionary adaptations. They can survive in the soils for many years in the absence of host plants through the formation of resistant structures such as microsclerotia (Verticillium spp.), sclerotia (Sclerotinia spp.), chlamydospores (Fusarium spp.), or oospores (Phytophthora spp.) [7][8][9][10]. Microsclerotia and sclerotia have the same anatomical structure, consisting of outer melanized parenchyma cells and inner colorless medullary cells, and are asexual in nature. Chlamydospores are thick-walled asexual survival structures whereas oospores are thick-walled sexual structures with food reserves for better survival. These structures may be melanised or non-melanised. Melanisation of survival structures has several evolutionary advantages such as protection from UV radiation, successful penetration during infection, long-term survival, growth, and development [11][12]. Wilhelm [13] found the persistence of microsclerotia of Verticillium alboatrum for 14 years in soil, which were viable even after the exposure to desiccation at high temperatures. Ben-Yephet et al. [14] reported the survival of sclerotia of soil-borne Sclerotinia sclerotiorum declined after an outbreak of lettuce drop, nevertheless, about 5.5% were viable even after seven years. Babadoost and Pavon [15] assessed the survival of Phytophthora capsici oospores in the soil in Illinois (USA) and found three to four years of survivability. Apart from soil-borne fungal plant pathogens, plant-parasitic nematodes have been recognized as another group of challenging pathogens to manage [16].
Besides, each plant can be infected by several pathogen species and the complex nature of the soil environment, it is difficult to control diseases caused by soil-borne pathogens. Hence, successful control of soil-borne pathogens is a major challenge due to inherent difficulties of disease prediction, early detection, and accurate diagnosis [2]. Some modern crop production systems are based on raised-bed, plasticulture, and limited or short crop rotation-lengths, probably with the unavoidable application of broad-spectrum soil fumigants to manage pests and diseases [1]. Since the mid-20th century, synthetic chemicals have been used to control many plant diseases including a broad spectrum soil fumigant, methyl bromide (MeBr) [17][18][19]. Since then, MeBr has been heavily applied worldwide primarily to control soil-borne pathogens as well as the nematodes [20]. For example, five million kg of MeBr were used only in California in the year 2000 [21]. MeBr has been identified as a stratospheric ozone-depleting component by the U.S. Environmental Protection Agency (EPA) and the United Nations Environment Program (UNEP). Bolstered by the 1994 UNEP Montreal Protocol on Substances that Deplete the Ozone Layer, MeBr was identified as a major ozone-depleting compound [22]. Thereafter, MeBr was completely banned by the 1 January 2005 with few exceptions [19][21][23][24].
Alternative synthetic fumigants such as 1,3-dichloropropene, 1,3-D, chloropicrin, trichloronitromethane, methyl isothiocyanate, allyl isothiocyanate (AITC), and dazomet were tested and applied by the farming communities around the world yet were poorly accepted due to geographic limitations, reduced efficacy, and regulatory constraints [25][26][27]. Moreover, many criticisms have been generated from the public and from the scientific communities against the use of such chemical soil disinfestation methods due to their toxicity on humans and undesirable effects on non-target organisms such as beneficial microflora, groundwater pollution, and development of resistance [19][28][29][30][31][32].
Therefore, farmers were compelled to use non-chemical approaches. Traditionally a number of environmental friendly approaches such as mixed cropping, crop rotation, resistant cultivars/selective breeding, application of biocontrol agents, flooding, solarisation, steaming, pasteurisation, hot water treatment, and bio-fumigation have been applied by farmers around the world to mitigate soil-borne diseases [19][33][34][35]. Nevertheless, these applications were not as popular as chemical fumigants due to several limitations [19]. Application of mixed cropping systems may be helpful in increasing the crop yield while addressing some of the soil-borne pathogen problems [36], yet it is not always economically feasible when the rotation is done with low economical value crops [35]. Although selective breeding shows some level of effectiveness against soil-borne pathogens, host resistance breakdown has been reported, and no completely resistant cultivars are available for all the crops [35] Another option would be the use of biocontrol agents, however, these are highly specific for particular pathogen species if not for strains, and effectiveness is greatly dependent on the environmental factors [37]. Similarly, other non-chemical approaches have their own disadvantages, hence there have been limited applications [33][36][38][39][40][41].
1.1. Anaerobic Soil Disinfestation (ASD)
To minimize the above drawbacks of chemical and non-chemical methods of soil-borne pathogen control, researchers found alternative methods, and one such promising approach is anaerobic soil disinfestation (ASD), also called biological soil disinfection (BSD) or reductive soil disinfection (RDS). This method was first described independently by researchers in Japan [42][43] and in the Netherlands [44] and was later adapted to the USA [45] to control soil-borne pathogens in strawberry and vegetable fields. Thereafter, researchers around the world started applying this method, showing a great potential to control various soil-borne phytopathogens [44][45][46][47][48][49][50][51][52][53][54][55].
The method is characterized by non-chemical pre-plant control of soil-borne phytopathogens using few simple steps [29][56]. The first step of ASD is the incorporation of organic amendments (usually an easily labile carbon source) to the topsoil. The soil is later wetted to field capacity and covered with a clear (preferably black) and gas-impermeable polyethylene sheet for a defined period of time to maintain an anaerobic condition [57]. The effectiveness of ASD has been evaluated against soil-borne diseases such as potato brown rot [46], spinach and tomato wilt diseases [48], Prunus [58] and apple replant disease [50], Fusarium wilt of banana [59], root and crown rot diseases of pepper [60], etc., with promising results. ASD has now become popular in organic agriculture worldwide and is practiced under greenhouse and field conditions as well [47][51][61]. There is some evidence that ASD also can contribute to the development of disease-suppressive soils [57].
2. Trends and Gaps in Application of ASD
2.1. Geographical Projection
When ASD was first introduced in Japan, it was initially suggested to be used with organic materials such as wheat bran, molasses, rice straw, and rice bran specifically at 1 to 2 tons per 0.1 ha, followed by flooding and plastic film covering of the soil surface [42]. In Netherlands, Blok et al. [44] carried out a two-year ASD field experiment in 1994 and 1995 using fresh broccoli or grass (3.4 to 4.0 kg fresh weight m−2) as C sources. They came up with the landmark finding that there was a significant control of soil-born fungal pathogens: Fusarium oxysporum, Rhizoctonia solani, and Verticillium dahliae. The study was published in 2000 and concluded that this novel method could control a wide range of phytopathogens [44]. Based on the published data, it is clear that the initial development of ASD was restricted to the Netherlands and Japan and was later expanded to the USA. However, beyond this point, ASD research showed slow progress until 2014, in which the number of publications were more than doubled (Figure 1). During the past few years, several other countries have also attempted to mitigate soil-borne diseases through ASD.
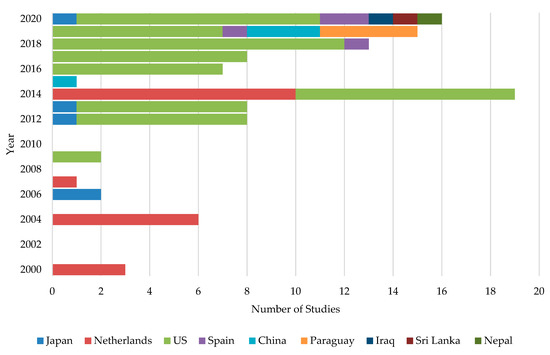
Figure 1. International distribution of ASD studies conducted in each year.
It is interesting to note that almost all the studies have been restricted to nine countries—primarily the USA (63.3%) followed by the Netherlands (18.3%) and Japan (4.6%). Spain, China, and Paraguay shared about 11.1% of ASD studies equally. The rest of the studies were conducted in Iraq, Sri Lanka, and Nepal, where only one study has been conducted in each country.
2.2. Application of ASD to Control Pathogens, Weeds, and Effect on Crop Yield
Initially, ASD studies were applied to control soil-borne phytopathogenic fungi [44]. Later on, it expanded towards control of nematodes, oomycetes, weeds, and protozoans. However, studies on ASD targeting soil borne-fungi have been extensively carried out mainly due to their broad host range, enormous losses in crop yield and quality, worldwide distribution, management difficulties, and extensive use of synthetic fungicides [62]. For example, 46.8% of the studies were concentrated on the control of fungal pathogens followed by 26.6%, 5.5%, and 4.6% of studies dedicating to testing the effects on nematodes, yield increase, and weed control, respectively. Moreover, about 12.8% of studies have been carried out with different aspects such as evaluating the effect of ASD on soil microflora and cost benefits of the application of ASD. Figure 2 shows the number of studies conducted in each year targeting soil-borne pathogens and other aspects. A majority (63%) of ASD studies were carried out under field conditions. About 35.1% of studies were performed as greenhouse or growth chamber experiments, while about 1.9% of the studies were conducted as lab experiments.
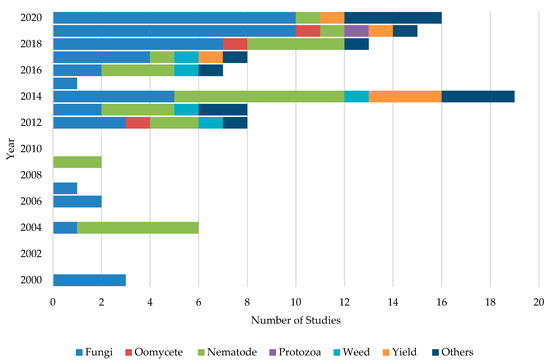
Figure 2. Different ASD studies conducted during the past two decades targeting each group of soil-borne pathogens and other aspects of crop production systems.
About 28.7% of the ASD studies were targeted to control tomato pathogens while 13%, 9.3%, and 7.4% of the studies were targeted to control strawberry, potato, and bell pepper pathogens, respectively. About 12.9% of the studies did not report the target crop or the intended pathogen to control. The remaining studies were carried out to control soil-borne pathogens associated with lettuce, mustard green, spinach, carrot, cabbage, cauliflower, eggplant, lily bulb, and common bean production fields. Studied organisms included pathogenic fungi: Fusarium oxysporum, Verticillium dahlia, Colletotrichum coccodes, Sclerotinia sclerotiorum, and S. rolfsii, nematodes: Meloidogyne hapla, M. incognita and Pyrenochaeta lycopersici, oomycetes: Phytophthora capsici and protist: Plasmodiophora brassicae etc.
2.3. C Source Dependency of ASD
The effectiveness of ASD predominantly depends on the selection of C source, C:N ratio, rate of its application, and anaerobic period. However, soil temperature, water holding capacity of soil, and climatic conditions should also be considered before implementing ASD [63][64][65]. C sources should be easily applicable, readily available/locally available, easily degradable, affordable, and able to control a broad spectrum of phytopathogens [65]. Careful selection of C source seems to play the key role in ASD since several studies have shown the emission of volatile compounds with strong pathogen inhibitory activities. Use of Brassica juncea cv. Pacific Gold seed meal (seed meal is a waste product of the oil extraction process) as the C source caused the release of isothiocyanates, alcohols, organic acids, organic sulphides, and esters, while application of orchard grass residues released organic sulfides, ketones, organic acids, and hydrocarbons. Similarly, the application of rice bran-treated soils emitted a spectrum of volatile compounds containing organic acids, alcohols, and esters [50][66]. Mahalingam et al. [55] conducting a gas chromatography-mass spectrometry (GC-MS) analysis of cabbage and leek cull piles reported the presence of antifungal volatiles.
Moreover, fresh and dried plant materials and composted broiler litter have been tested in multiple studies as the C source in ASD-based studies [54][65][67]. Ethanol has been incorporated as a C source in controlling phytopathogens due to the inefficiency of some of the commonly used C sources. As an example, Momma et al. [68] found that the use of wheat bran alone is not effective in controlling Fusarium oxysporum infection of tomato. However, once the soil is saturated with 1% ethanol solution (ethanol medicated ASD treatment), high levels of suppression of F. oxysporum were observed. Hewavitharana et al. [53] also reported that ethanol (10%) mediated ASD effectively controlled apple root infection caused by Rhizoctonia solani AG-5 and Pratylenchus penetrans. In addition, it has been reported that ethanol temporarily increased the anaerobic bacterial population [68]. A summary of C source, application rate, target pathogen group, and optimum temperature along with the reference are shown in the Table 1 below.
Table 1. Details of successful ASD experiments conducted during the past few years.
|
C Source |
Application Rate of C Sources (t ha−1) |
Pathogens Suppressed |
Mean Soil Temperature/Range (°C) |
Treatment Period |
Crop |
Field/Greenhouse |
Country | Reference |
|---|---|---|---|---|---|---|---|---|
|
Fresh broccoli (Brassica oleracea) |
34,38 |
Fusarium oxysporum, Rhizoctonia solani, Verticillium dahliae |
25–32, 29–39 |
15 weeks |
N/A |
Field, plot |
Netherlands | [44] |
|
Perennial ryegrass (Lolium perenne) |
40 |
Fusarium oxysporum, Rhizoctonia solani, Verticillium dahliae |
25–32, 29–39 |
15 weeks |
N/A |
Field, plot |
Netherlands | [44] |
|
Grass or potato haulms |
30 |
Ralstonia solanacearum |
N/A |
6 weeks |
Potato |
Laboratory, field |
Netherlands | [46] |
|
Wheat bran |
2 |
Meloidogyne incognita |
35.0 |
24 days |
Tomato |
Greenhouse, plot |
Japan | [47] |
|
Cereal rye (Secale cereale) |
0.134 |
Rhizoctonia solani |
20.8 |
4 weeks |
Tomato, bell pepper |
Field, plot |
USA | [49] |
|
Mustard (Brassica juncea) seed meal |
4.9 |
Rhizoctonia solani, Pythium ultimum, Fusarium oxysporum |
18–24 |
2 weeks |
Apple |
Growth chamber, pot |
USA | [50] |
|
Grass residues |
40.0 |
Rhizoctonia solani, Pythium ultimum, Fusarium oxysporum |
18–24 |
2 weeks |
Apple |
Growth chamber, pot |
USA | [50] |
|
Rice bran |
20 |
Verticillium dahliae |
21–23 |
4 weeks |
Strawberries |
Field |
USA | [50] |
|
Radish roots |
100 |
Fusarium oxysporum |
33.1 |
3 weeks |
Spinach |
Greenhouse, field |
Japan | [48] |
|
Mixture of fresh rye-grass species |
50 |
Verticillium dahliae, Pasteuria penetrans |
N/A |
12 weeks |
N/A |
Field |
Netherlands | [48] |
|
Mustard (Brassica juncea) |
50 |
Fusarium oxysporum |
33.1 |
3 weeks |
Spinach |
Greenhouse, pots, field |
Japan | [48] |
|
Wheat bran |
20 |
Fusarium oxysporum |
33.1 |
3 weeks |
Spinach |
Green house, pots, field |
Japan | [48] |
|
Rice bran |
4.4 |
Rhizoctonia solani, Pratylenchus penetrans |
18–24 |
2 weeks |
Apple |
Growth chamber, pot |
USA | [53] |
|
Fresh orchard grass residues |
20 |
Rhizoctonia solani, Pratylenchus penetrans |
18–24 |
2 weeks |
Apple |
Growth chamber, pot |
USA | [53] |
|
Mustard (Brassica juncea) seed meal |
4.4 |
Rhizoctonia solani, Pratylenchus penetrans |
18–24 |
2 weeks |
Apple |
Growth chamber, pot |
USA | [53] |
|
Rice bran |
20 |
Phytophthora nicotianae |
15–35 |
4 weeks |
Pepper |
Field |
Spain | [54] |
|
Rapeseed cake |
20 |
Phytophthora nicotianae |
15–35 |
4 weeks |
Pepper |
Field, plot |
Spain | [54] |
|
Grape pomace |
40 |
Phytophthora nicotianae |
15–35 |
4 weeks |
Pepper |
Field, plot |
Spain | [54] |
|
Rice bran |
20 |
Fusarium oxysporum |
18–24 |
15 days |
Strawberry |
Growth chamber, pot |
USA | [69] |
N/A: not available or not reported.
2.4. ASD against Nematodes
Plant pathogenic nematodes are another group of organisms posing a severe threat to worldwide agriculture, especially in developing countries. With the limited availability of nematicides, negative impacts of available chemistries and resistance development have always demanded alternative management options [70]. ASD has shown promising results in controlling plant-parasitic nematodes in several studies conducted in the USA [71][72] and in the Netherlands [52]. A study conducted by Mazzola et al. [73] in the USA found successful control of Pratylenchus penetrans in strawberry fields when Brassica juncea seed meal was used as the C source, whereas Testen and Miller [74] reported reduction of M. hapla when wheat bran and molasses were used. Similarly, tomato plant residues with fresh sheep manure were effective in controlling M. incognita [75]. Korthals et al. [52] reported that ASD was more effective and longer-lasting against P. penetrans and V. dahliae than chemical control, and Di Gioia et al. [76] also reported ASD was effective as chemical soil fumigation against Meloidogyne sp. However, it should be noted that the selection of C source should be done carefully, and targeted organism should be taken into account. As an example, Korthals et al. [52] demonstrated that B. junceae leaf incorporation (no anaerobic condition was imposed) increased P. penetrans density in soil.
2.5. Effect of ASD on Weed Control and Yield
Weed control is one of the major requirements, especially in the tropics, where year-round cultivation is practiced, favouring accumulation of weed seed banks. Each year, billions of dollars are spent on herbicide development and applications. Application of one of the most controversial yet most effective herbicides, glyphosate, has dramatically increased in the last two decades and by the end of 2014, 8.6 billion kg of glyphosate had been applied globally [77]. However, recently, there are serious concerns over the use of glyphosate and its negative impacts on human and environmental health. Mixed results have been presented in terms of the effect of ASD on weed control. While Shennan et al. [66] and Guo et al. [64] reported a low success rate or failure of weed control during ASD with either rice bran or molasses as C sources, McCarty et al. [49] reported effective weed control with cereal rye and mustard/arugula in Tennessee, USA. Lamers et al. [78] emphasized that green manure crops should be amended at least at 40 t ha−1 rate to achieve weed control in the Netherlands. Although not as effective as chemical treatments, Brassicaceae residues are also effective in weed management [79]. However, Amaranthus retroflexus (an opportunistic annual weed of many cropping systems) is reported to be one of the most challenging weed to control through ASD [19][80]. It appears that the weed control ability of ASD is due to phytotoxicity/phytotoxic volatiles generated by microbial activities.
Only six studies have been conducted to determine the effect of ASD on the crop yield. Korthals et al. [52] determined the effect of nine different treatments including ASD on crop yields of potato, carrot, and lily bulb and reported that ASD produced higher yields in all the crops compared to the untreated control. However, Di Gioia et al. [81] reported that ASD had no significant effect on tomato yield when composted poultry litter (22 Mg ha−1) and molasses (13.9 and 27.7 m3 ha−1) were used as the C sources. Nevertheless, plant nutrients such as potassium, calcium, magnesium, and iron accumulation had improved in ASD treated plants. Yield improvement might have resulted due to the combined effects of disease control, weed control, and improved soil nutrients.
2.6. Mechanism of ASD
Only 34% of studies have reported the mechanism of ASD. Nevertheless, the exact mechanism of ASD is still not clear, and further studies are necessary. In ASD, the use of different carbon sources helps boosting soil microbial biomass and enzyme activities [54]. Covering with a plastic trap as well as the utilization of available oxygen by the aerobic microorganisms ultimately create an anaerobic soil condition. Figure 3 shows possible soil pathogen control mechanism(s) by ASD.
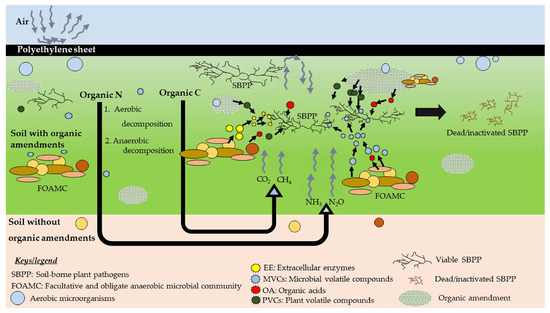
Figure 3. Proposed pathogen control mechanism(s) (simplified) during ASD. The decomposition of organic amendments is initiated by the activities of aerobic microorganisms (e.g., Bacillus spp.). Later, the growth of anaerobic bacteria (e.g., Clostridium spp.) is stimulated with the depletion of oxygen, and anaerobic decomposition of organic matter is initiated. This pathway is more complex and less energy demanding than that of the aerobic decomposition. Facultative and obligate anaerobic microbial communities (FOAMC) decompose the added organic C and produce several gases such as CO2, CH4, and volatile compounds. Decomposition of organic N leads to produce soil ammonium (NH4+) via mineralization. Finally, due to series of activities, CO2, CH4, N2O, and NH3 are released, and these gases have toxic effect on living matter. Combined effects of above released gases along with organic acids (OA), microbes released extracellular enzymes (EE), microbial volatile compounds (MVCs), and plant volatile/non-volatile compounds (PVCs) along with the change of soil physical properties may cause the inhibition of soil-borne phytopathogens (SBPP).
Polyethylene sheets prevent further penetration of oxygen to the treatment creating a conducive environment for anaerobic microorganisms (e.g., Clostridial species). These anaerobic decomposers use C source to respire while releasing toxic anaerobic by-products such as CO2, NH3, H2S, CH4, and N2O [19]. However, these by-products are released to the atmosphere quickly, as soon as the tarp is removed or the holes are punched [79]. Researchers predicted that the limitation of oxygen along with the trapping of toxic compounds and lowered pH could control soil-borne phytopathogens [44]. Under the flooded conditions, microbes decompose liable C sources and release gases (or by-products) suppressing some of the phytopathogens [19]. ASD has shown significant changes in the whole soil microbial communities [46][65]. Mowlick et al. [82] reported the changes in microbial community structures (through clone library analysis) after ASD treatment. They observed ASD caused a reduction in diversity of bacterial communities of various phylogenetic groups and a domination of anaerobic clostridial class bacteria.
In ASD, accumulation of various volatile compounds with the potential to control phytopathogens greatly depends on the C source used [50][55][66][83][84]. In addition to pathogen control, plant growth promotion abilities of microbial volatile compounds (MVCs) have also been extensively reported [84][85][86]. These volatile compounds spread through soil by diffusion, and efficacy of volatile compounds is greater than non-volatile compounds [87][88][89]. Compared to the other MVCs such as enzymes, antibiotics, and toxins, microbial organic volatiles are typically small in size (up to 20 carbon atoms) with molecular mass ranging from 100 to 500 Daltons [85]. MVCs have a good diffusing ability under normal temperatures and pressures [90]. Volatile compounds produced by the bacteria are dominated by alkenes, alcohols, ketones, terpenes, benzenoids, pyrazines, acids, and esters, while fungal volatiles are dominated by alcohols, benzenoids, aldehydes, alkenes, acids, esters, and ketones [85]. Antifungal compounds such as dimethyl disulfide, dimethyl trisulfide, and acetoin are well reported [90]. Studies found that the fungal species such as Aspergillus giganteus, Fusarium oxysporum, Penicillium viridicatum, Trichoderma viride, and Zygorhynchus vuilleminii have abnormal morphologies in their conidiophores and hyphae when exposed to VOCs from bacteria and actinomycetes [91]. Rather than the production of volatile compounds [48], soil anaerobic bacterial communities could kill the phytopathogenic fungi through extracellular enzymes such as 1,3-glucanase and chitosanase, whereas obligate anaerobic Clostridium beijerinckii could suppress the spinach wilt fungi, F. oxysporum and F. spinaciae [61]. However, the prevailing groups of the microorganisms may be different based on the type of C source used and the treatment period of ASD, since some microorganisms are responsive to fluctuating redox potentials [45][92].
References
- Butler, D.M.; Rosskopf, E.N.; Kokalis-Burelle, N.; Albano, J.P.; Muramoto, J.; Shennan, C. Exploring Warm-season Cover Crops as Carbon Sources for Anaerobic Soil Disinfestation (ASD). Plant. Soil. 2012, 355, 149–165.
- Mihajlovic, M.; Rekanovic, E.; Hrustic, J.; Grahovac, M.; Tanovic, B. Methods for Management of Soil-borne Plant Pathogens. Pestic. Fitomed. 2017, 32, 9–24.
- Panth, M.; Hassler, S.C.; Baysal-Gurel, F. Methods for Management of Soil-borne Diseases in Crop Production. Agriculture 2020, 10, 16.
- Duncan, J.M.; Kennedy, D.M. The Effect of Waterlogging on Phytophthora Root Rot of Red Raspberry. Plant Pathol. 1989, 38, 161–168.
- Meisner, A.; de Boer, W. Strategies to Maintain Natural Biocontrol of Soil-borne Crop Diseases during Severe Drought and Rainfall Events. Front. Microbiol. 2018, 9, 1–8.
- Punja, Z.; Collyer, D.; Scott, C.; Lung, S.; Holmes, J.; Sutton, D. Pathogens and Molds Affecting Production and Quality of Cannabis sativa L. Front. Plant. Sci. 2019, 10, 1–23.
- Ho, H. The Taxonomy and Biology of Phytophthora and Pythium. JBMOA 2018, 6, 1–7.
- Niu, X.; Ah-Fong, A.; Lopez, L.; Judelson, H. Transcriptomic and Proteomic Analysis Reveals Wall-associated and Glucan-degrading Proteins with Potential Roles in Phytophthora infestans Sexual Spore Development. PLoS ONE 2018, 13, 198186.
- Prova, A.; Akanda, A.M.; Islam, S.; Hossain, M.M. Characterization of Sclerotinia sclerotiorum, an Emerging Fungal Pathogen Causing Blight in Hyacinth Bean (Lablab purpureus). Plant Pathol. J. 2018, 34, 367–380.
- Taylor, A.; Coventry, E.; Handy, C.; West, J.S.; Young, C.S.; Clarkson, J.P. Inoculum Potential of Sclerotinia sclerotiorum Sclerotia Depends on Isolate and Host Plant. Plant Pathol. 2018, 67, 1286–1295.
- Cheung, N.; Tian, L.; Liu, X.; Li, X. The Destructive Fungal Pathogen Botrytis cinerea-Insights from Genes Studied with Mutant Analysis. Pathogens 2020, 9, 923.
- Xia, S.; Xu, Y.; Hoy, R.; Zhang, J.; Qin, L.; Li, X. The Notorious Soil-borne Pathogenic Fungus Sclerotinia sclerotiorum: An Update on Genes Studied with Mutant Analysis. Pathogens 2019, 9, 27.
- Wilhelm, S. Longevity of the Verticillium Wilt Fungus in the Laboratory and Field. Phytopathology 1955, 45, 180–181.
- Ben-Yephet, Y.; Genizi, A.; Siti, E. Sclerotial Survival and Apothecial Production by Sclerotinia sclerotiorum Following Outbreaks of Lettuce Drop. Phytopathology 1993, 83, 509–513.
- Babadoost, M.; Pavon, C. Survival of Oospores of Phytophthora capsici in Soil. Plant Dis. 2013, 97, 1478–1483.
- Basyony, A.G.; Abo-Zaid, G.A. Biocontrol of the Root-knot Nematode, Meloidogyne incognita, Using an Eco-friendly Formulation from Bacillus subtilis, lab. and Greenhouse Studies. Egypt. J. Biol. Pest Control. 2018, 28, 1–13.
- Hancock, T.L.C.; Costello, A.M.; Lidstrom, M.E.; Oremland, R.S. Strain IMB-1, A Novel Bacterium for the Removal of Methyl Bromide in Fumigated Agricultural Soils. Appl. Environ. Microbiol. 1998, 64, 2899–2905.
- Barry, K.H.; Koutros, S.; Lubin, J.H.; Coble, J.B.; Barone-Adesi, F.; Beane Freeman, L.E.; Sandler, D.P.; Hoppin, J.A.; Ma, X.; Zheng, T.; et al. Methyl Bromide Exposure and Cancer Risk in the Agricultural Health Study. Cancer Causes Control 2012, 23, 807–818.
- Shrestha, U.; Augé, R.M.; Butler, D.M.A. Meta-analysis of the Impact of Anaerobic Soil Disinfestation on Pest Suppression and Yield of Horticultural Crops. Front. Plant Sci. 2016, 7, 1–20.
- Mao, L.; Jiang, H.; Zhang, L.; Zhang, Y.; Sial, M.U.; Yu, H.; Cao, A. Replacing Methyl Bromide with a Combination of 1,3-dichloropropene and Metam Sodium for Cucumber Production in China. PLoS ONE 2017, 12, 188137.
- Gemmill, A.; Gunier, R.B.; Bradman, A.; Eskenazi, B.; Harley, K.G. Residential Proximity to Methyl Bromide use and Birth Outcomes in an Agricultural Population in California. Environ. Health Perspect. 2013, 121, 737–743.
- Thomas, W.B. Methyl Bromide: Effective Pest Management Tool and Environmental Threat. J. Nematol. 1996, 28, 586–589.
- Theis, J.A.; Fery, R.L. Host Plant Resistance as an Alternative to Methyl Bromide for Managing Meloidogyne incognita in Pepper. J. Nematol. 2002, 34, 374–377.
- Velders, G.J.M.; Andersen, S.O.; Daniel, J.S.; Fahey, D.W.; McFarland, M. The Importance of the Montreal Protocol in Protecting Climate. Proc. Natl. Acad. Sci. USA 2007, 104, 4814–4819.
- Martin, F.N. Development of Alternatives Strategies for Management of Soil-borne Pathogens Currently Controlled with Methyl Bromide. Annu. Rev. Phytopathol. 2003, 41, 325–350.
- Sande, D.; Mullen, J.; Wetzstein, M.; Houston, J. Environmental Impacts from Pesticide Use: A Case Study of Soil Fumigation in Florida Tomato Production. IJERPH 2011, 8, 4649–4661.
- Desaeger, J.; Dickson, D.W.; Locascio, S.J. Methyl Bromide Alternatives for Control of Root-knot Nematode (Meloidogyne spp.) in Tomato Production in Florida. J. Nematol. 2017, 49, 140–149.
- Yoon, M.Y.; Cha, B.; Kim, J.C. Recent Trends in Studies on Botanical Fungicides in Agriculture. Plant Pathol. J. 2013, 29, 1–9.
- Strauss, S.L.; Kluepfel, D.A. Anaerobic Soil Disinfestation: A chemical-independent approach to Pre-plant Control of Plant Pathogens. J. Integr. Agric. 2015, 14, 2309–2318.
- Ghosh, S.K. Application of Synthetic Chemicals in Agriculture and their Toxic Effect on the Environment. Bull. Env. Pharmacol. Life Sci. 2015, 4, 1–6.
- Jayaraj, R.; Megha, P.; Sreedev, P. Organochlorine Pesticides, their Toxic Effects on Living Organisms and their Fate in the Environment. Interdiscip. Toxicol. 2016, 9, 90–100.
- Kaur, T.; Rani, R.; Manhas, R.K. Biocontrol and Plant Growth Promoting Potential of Phylogenetically New Streptomyces sp. MR14 of Rhizospheric Origin. AMB. Expr. 2019, 9, 1–14.
- Runia, W.T.; Molendijk, L.P.G. Physical Methods for Soil Disinfestation in Intensive Agriculture: Old Methods and New Approaches. Acta. Hortic. 2010, 249–258.
- Weerakoon, D.M.N.; Reardon, C.L.; Paulitz, T.C.; Izzo, A.D.; Mazzola, M. Long-term Suppression of Pythium abappressorium Induced by Brassica juncea Seed Meal Amendment is Biologically Mediated. Soil Biol. Biochem. 2012, 51, 44–52.
- Newitt, J.; Prudence, S.; Hutchings, M.; Worsley, S. Biocontrol of Cereal Crop Diseases Using Streptomycetes. Pathogens 2019, 8, 78.
- Li, M.; Zhang, J.; Liu, S.; Ashraf, U.; Zhao, B.; Qiu, S. Mixed-cropping Systems of Different Rice Cultivars Have Grain Yield and Quality Advantages over Mono-cropping Systems. J. Sci. Food. Agric. 2019, 99, 3326–3334.
- Vitorino, L.C.; Silva, F.; Cruvinel, B.G.; Bessa, L.A.; Rosa, M.; Souchie, E.L.; Silva, F.G. Biocontrol Potential of Sclerotinia sclerotiorum and Physiological Changes in Soybean in Response to Butia archeri Palm Rhizobacteria. Plants 2020, 9, 64.
- Ploeg, A. Biofumigation to Manage Plant-parasitic Nematodes. In Integrated Management and Biocontrol of Vegetable and Grain Crops Nematodes; Ciancio, A., Mukerji, K.G., Eds.; Springer: Dordrecht, The Netherlands, 2008; Volume 2, pp. 239–248.
- Chandrashekara, K.N.; Manivannan, S.; Chandrashekara, C.; Chakravarthi, M. Biological Control of Plant Diseases. In Ecofriendly Innovative Approaches in Plant Disease Management; Vaibhav, K.S., Yogendra, S., Akhilesh, S., Eds.; International Book Distributors: New Delhi, India, 2012; pp. 147–166.
- Capstaff, N.M.; Miller, A.J. Improving the Yield and Nutritional Quality of Forage Crops. Front. Plant Sci. 2018, 9, 1–18.
- Michel, V.; Cara-García, M.D. Bio-fumigation: Practical Information, Advantages and Disadvantages. Best4soil Facts Sheet 2020. Available online: https://www.best4soil.eu/assets/factsheets/11.pdf (accessed on 2 May 2020).
- Momma, N. Biological Soil Disinfestation (BSD) of Soil-borne Pathogens and its Possible Mechanisms. JARQ 2008, 42, 7–12.
- Momma, N.; Kobara, Y.; Uematsu, S.; Kita, N.; Shinmura, A. Development of Biological Soil Disinfestations in Japan. Appl. Microbiol. Biotechnol. 2013, 97, 3801–3809.
- Blok, W.J.; Lamers, J.G.; Termorshuizen, A.J.; Bollen, G.J. Control of Soil-borne Plant Pathogens by Incorporating Fresh Organic Amendments Followed by Trapping. Phytopathology 2000, 90, 253–259.
- Shennan, C.; Muramoto, J.; Lamers, J.; Mazzola, M.; Rosskopf, E.N.; Kokalis-Burelle, N.; Momma, N.; Butler, D.M.; Kobara, Y. Anaerobic Soil Disinfestation for Soil Borne Disease Control in Strawberry and Vegetable Systems: Current Knowledge and Future Directions. Acta Hortic. 2014, 1044, 165–175.
- Messiha, N.A.S.; van Diepeningen, A.D.; Wenneker, M.; van Beuningen, A.R.; Janse, J.D.; Coenen, T.G.C.; Termorshuizen, A.J.; Van Bruggen, A.H.C.; Blok, W.J. Biological Soil Disinfestation (BSD), a New Control Method for Potato Brown Rot, Caused by Ralstonia solanacearum race 3 biovar 2. Eur. J. Plant Pathol. 2007, 117, 403–415.
- Katase, M.; Kubo, C.; Ushio, S.; Ootsuka, E.; Takeuchi, T.; Mizukubo, T. Nematicidal Activity of Volatile Fatty Acids Generated from Wheat Bran in Reductive Soil Disinfestation. Jpn. J. Nematol. 2009, 39, 53–62.
- Mowlick, S.; Yasukawa, H.; Inoue, T.; Takehara, T.; Kaku, N.; Ueki, K.; Ueki, A. Suppression of Spinach Wilt Disease by Biological Soil Disinfestation Incorporated with Brassica juncea Plants in Association with Changes in Soil Bacterial Communities. Crop Prot. 2013, 54, 185–193.
- McCarty, D.G.; Eichler Inwood, S.E.; Ownley, B.H.; Sams, C.E.; Wszelaki, A.L.; Butler, D.M. Field Evaluation of Carbon Sources for Anaerobic Soil Disinfestation in Tomato and Bell Pepper Production in Tennessee. Hort. Sci. 2014, 49, 272–280.
- Hewavitharana, S.S.; Ruddell, D.; Mazzola, M. Carbon Source-dependent Antifungal and Nematicidal Volatiles Derived during Anaerobic Soil Disinfestation. Eur. J. Plant Pathol. 2014, 140, 39–52.
- Muramoto, J.; Shennan, C.; Baird, G.; Zavatta, M.; Koike, S.T.; Bolda, M.P.; Daugovish, O.; Dara, S.K.; Klonsky, K.; Mazzola, M. Optimizing Anaerobic Soil Disinfestation for California Strawberries. Acta Hortic. 2014, 215–220.
- Korthals, G.W.; Thoden, T.C.; van den Berg, W.; Visser, J.H.M. Long-term Effects of Eight Soil Health Treatments to Control Plant-parasitic Nematodes and Verticillium dahliae in Agro-ecosystems. Appl. Soil Ecol. 2014, 76, 112–123.
- Hewavitharana, S.S.; Mazzola, M. Carbon Source-dependent Effects of Anaerobic Soil Disinfestation on Soil Microbiome and Suppression of Rhizoctonia solani AG-5 and Pratylenchus penetrans. Phytopathology 2016, 106, 1015–1028.
- Serrano-Pérez, P.; Rosskopf, E.; De Santiago, A.; del Carmen Rodríguez-Molina, M. Anaerobic Soil Disinfestation Reduces Survival and Infectivity of Phytophthora nicotianae Chlamydospores in Pepper. Sci. Hortic. 2017, 215, 38–48.
- Mahalingam, T.; Rajapakse, C.S.K.; Somachandra, K.P.; Attanayake, R.N. Carbon Source Dependent-anaerobic Soil Disinfestation (ASD) Mitigates the Sclerotial Germination of Sclerotinia sclerotiorum. Trop. Plant Pathol. 2020, 45, 13–24.
- Núñez-Zofío, M.; Garbisu, C.; Larregla, S. Application of Organic Amendments Followed by Plastic Mulching for the Control of Phytophthora Root Rot of Pepper in Northern Spain. Acta Hortic. 2010, 353–360.
- Rosskopf, E.N.; Serrano-Pérez, P.; Hong, J.; Shrestha, U.; Rodríguez-Molina, M.d.C.; Martin, K.; Kokalis-Burelle, N.; Shennan, C.; Muramoto, J.; Butler, D. Anaerobic Soil Disinfestation and Soil-borne Pest Management. In Soil Biology; Springer: Cham, Switzerland, 2015; pp. 277–305.
- Browne, G.; Ott, N.; Poret-Peterson, A.; Gouran, H.; Lampinen, B. Efficacy of Anaerobic Soil Disinfestation for Control of Prunus Replant Disease. Plant Dis. 2018, 102, 209–219.
- Wen, T.; Huang, X.; Zhang, J.; Zhu, T.; Meng, L.; Cai, Z. Effects of Water Regime, Crop Residues, and Application Rates on Control of Fusarium oxysporum f. sp. cubense. J. Environ Sci. 2015, 31, 30–37.
- Gandariasbeitia, M.; Ojinaga, M.; Orbegozo, E.; Ortíz-Barredo, A.; Núñez-Zofío, M.; Mendarte, S.; Larregla, S. Winter Bio-disinfestation with Brassica Green Manure is a Promising Management Strategy for Phytophthora capsici Control of Protected Pepper Crops in Humid Temperate Climate Regions of Northern Spain. Span. J. Agric. Res. 2019, 17, 1–11.
- Ueki, A.; Kaku, N.; Ueki, K. Role of Anaerobic Bacteria in Biological Soil Disinfestation for Elimination of Soil-borne Plant Pathogens in Agriculture. Appl. Microbiol. Biotechnol. 2018, 102, 6309–6318.
- Yang, J.; Hsiang, T.; Bhadauria, V.; Chen, X.-L.; Li, G. Plant Fungal Pathogenesis. BioMed Res. Int. 2017, 1–2.
- Butler, D.M.; Ownley, B.H.; Dee, M.E.; Eichler Inwood, S.E.; McCarty, D.G.; Shrestha, U.; Kokalis-Burelle, N.; Rosskopf, E.N. Low Carbon Amendment Rates During Anaerobic Soil Disinfestation (ASD) at Moderate Soil Temperatures Do Not Decrease Viability of Sclerotinia Sclerotiorum Sclerotia or Fusarium Root Rot of Common Bean. Acta Hortic. 2014, 203–208.
- Guo, H.; Di Gioia, F.; Zhao, X.; Ozores-Hampton, M.; Swisher, M.E.; Hong, J.; Kokalis-Burelle, N.; DeLong, A.N.; Rosskopf, E.N. Optimizing Anaerobic Soil Disinfestation for Fresh Market Tomato Production: Nematode and Weed Control, Yield, and Fruit Quality. Sci. Hortic. 2017, 218, 105–116.
- Poret-Peterson, A.T.; Albu, S.; McClean, A.E.; Kluepfel, D.A. Shifts in Soil Bacterial Communities as a Function of Carbon Source Used During Anaerobic Soil Disinfestation. Front. Environ. Sci. 2019, 6, 1–15.
- Shennan, C.; Muramoto, J.; Koike, S.; Baird, G.; Fennimore, S.; Samtani, J.; Bolda, M.; Dara, S.; Daugovish, O.; Lazarovits, G.; et al. Anaerobic Soil Disinfestation is an Alternative to Soil Fumigation for Control of Some Soil-borne Pathogens in Strawberry Production. Plant Pathol. 2017, 67, 51–66.
- Achmon, Y.; Harrold, D.R.; Claypool, J.T.; Stapleton, J.J.; Vander Gheynst, J.S.; Simmons, C.W. Assessment of Tomato and Wine Processing Solid Wastes as Soil Amendments for Biosolarization. Waste Manag. 2016, 48, 156–164.
- Momma, N.; Momma, M.; Kobara, Y. Biological Soil Disinfestation Using Ethanol: Effect on Fusarium oxysporum f. sp. lycopersici and Soil Microorganisms. J. Gen. Plant Pathol. 2010, 76, 336–344.
- Hewavitharana, S.S.; Klarer, E.; Reed, A.J.; Leisso, R.; Poirier, B.; Honaas, L.; Rudell, D.R.; Mazzola, M. Temporal Dynamics of the Soil Metabolome and Microbiome During Simulated Anaerobic Soil Disinfestation. Front. Microbiol. 2019, 10, 1–17.
- Collange, B.; Navarrete, M.; Peyre, G.; Mateille, T.; Tchamitchian, M. Root-knot Nematode (Meloidogyne) Management in Vegetable Crop Production: The Challenge of an Agronomic System Analysis. Crop Protection 2011, 30, 1251–1262.
- Butler, D.M.; Kokalis-Burelle, N.; Muramoto, J.; Shennan, C.; McCollum, T.G.; Rosskopf, E.N. Impact of Anaerobic Soil Disinfestation Combined with Soil Solarization on Plant-parasitic Nematodes and Introduced Inoculum of Soil-borne Plant Pathogens in Raised-bed Vegetable Production. Crop Prot. 2012, 39, 33–40.
- Testen, A.L.; Miller, S.A. Carbon Source and Soil Origin Shape Soil Microbiomes and Tomato Soil-borne Pathogen Populations during Anaerobic Soil Disinfestation. Phytobiomes J. 2018, 2, 138–150.
- Mazzola, M.; Hewavitharana, S.S.; Strauss, S.L.; Shennan, C.; Muramoto, J. Anaerobic Soil Disinfestation and Brassica Seed Meal Amendment Alter Soil Microbiology and System Resistance. Int. J. Fruit Sci. 2016, 16, 47–58.
- Testen, A.L.; Miller, S.A. Anaerobic Soil Disinfestation to Manage Soil-borne Diseases in Muck Soil Vegetable Production Systems. Plant Dis. 2019, 103, 1757–1762.
- Gómez-Tenorio, M.A.; Lupión-Rodríguez, B.; Boix-Ruiz, A.; Ruiz-Olmos, C.; Marín-Guirao, J.I.; Tello-Marquina, J.C.; Camacho-Ferre, F.; de Cara-García, M. Meloidogyne-infested Tomato Crop Residues are a Suitable Material for Biodisinfestation to Manage Meloidogyne sp. in Greenhouses in Almería (South-east Spain). Acta Hortic. 2018, 217–222.
- Di Gioia, F.; Ozores-Hampton, M.; Hong, J.; Kokalis-Burelle, N.; Albano, J.; Zhao, X.; Black, Z.; Gao, Z.; Wilson, C.; Thomas, J.; et al. The Effects of Anaerobic Soil Disinfestation on Weed and Nematode Control, Fruit Yield, and Quality of Florida Fresh-market Tomato. Hort. Sci. 2016, 51, 703–711.
- Benbrook, C.M. Trends in Glyphosate Herbicide use in The United States and Globally. Environ Sci. Eur. 2016, 28, 1–15.
- Lamers, J.G.; Runia, W.T.; Molendijk, L.P.G.; Bleeker, P.O. Perspectives of Anaerobic Soil Disinfestation. Acta Hortic. 2010, 277–283.
- Fennimore, S.A.; Serohijos, R.; Samtani, J.B.; Ajwa, H.A.; Subbarao, K.V.; Martin, F.N.; Daugovish, O.; Legard, D.; Browne, G.T.; Muramoto, J.; et al. TIF Film, Substrates and Non-fumigant Soil Disinfestation Maintain Fruit Yields. Cal. Ag. 2013, 67, 139–146.
- Karimmojeni, H.; Bazrafshan, A.H.; Majidi, M.M.; Torabian, S.; Rashidi, B. Effect of Maternal Nitrogen and Drought Stress on Seed Dormancy and Germinability of Amaranthus retroflexus. Plant Species Biol. 2013, 29, E1–E8.
- Di Gioia, F.; Ozores-Hampton, M.; Zhao, X.; Thomas, J.; Wilson, P.; Li, Z.; Hong, J.; Albano, J.; Swisher, M.; Rosskopf, E. Anaerobic Soil Disinfestation Impact on Soil Nutrients Dynamics and Nitrous Oxide Emissions in Fresh-market Tomato. Agric. Ecosyst. Environ. 2017, 240, 194–205.
- Mowlick, S.; Inoue, T.; Takehara, T.; Kaku, N.; Ueki, K.; Ueki, A. Changes and Recovery of Soil Bacterial Communities Influenced by Biological Soil Disinfestation as Compared with Chloropicrin-treatment. AMB Express. 2013, 3, 1–12.
- Liu, W.; Mu, W.; Zhu, B.; Du, Y.; Liu, F. Antagonistic Activities of Volatiles from Four Strains of Bacillus spp. and Paenibacillus spp. against Soil-borne Plant Pathogens. Agr. Sci. China. 2008, 7, 1104–1114.
- Piechulla, B.; Degenhardt, J. The Emerging Importance of Microbial Volatile Organic Compounds. Plant Cell Environ. 2014, 37, 811–812.
- Schmidt, R.; De Jager, V.; Zühlke, D.; Wolff, C.; Bernhardt, J.; Cankar, K.; Beekwilder, J.; Van Ijcken, W.; Sleutels, F.; De Boer, W.; et al. Fungal Volatile Compounds Induce Production of the Secondary Metabolite Sodorifen in Serratia plymuthica PRI-2C. Sci. Rep. 2017, 7, 1–14.
- Sharifi, R.; Ryu, C.M. Biogenic Volatile Compounds for Plant Disease Diagnosis and Health Improvement. Plant Pathol. J. 2018, 34, 459–469.
- Garbeva, P.; Hordijk, C.; Gerards, S.; de Boer, W. Volatile-mediated Interactions between Phylogenetically Different Soil Bacteria. Front. Microbiol. 2014, 5, 1–9.
- Kanchiswamy, C.N.; Malnoy, M.; Maffei, M.E. Chemical Diversity of Microbial Volatiles and their Potential for Plant Growth and Productivity. Front. Plant Sci. 2015, 6, 1–23.
- Schulz-Bohm, K.; Martín-Sánchez, L.; Garbeva, P. Microbial Volatiles: Small Molecules with an Important Role in Intra- and Inter-kingdom Interactions. Front. Microbiol. 2017, 8, 1–10.
- Morita, T.; Tanaka, I.; Ryuda, N.; Ikari, M.; Ueno, D.; Someya, T. Antifungal Spectrum Characterization and Identification of Strong Volatile Organic Compounds Produced by Bacillus pumilus TM-R. Heliyon 2019, 5, 1–8.
- Wang, C.; Wang, Z.; Qiao, X.; Li, Z.; Li, F.; Chen, M.; Wang, Y.; Huang, Y.; Cui, H. Antifungal Activity of Volatile Organic Compounds from Streptomyces alboflavus TD-1. FEMS Microbiol. Lett. 2013, 341, 45–51.
- Van Agtmaal, M.; Van Os, G.J.; Hol, W.H.G.; Hundscheid, M.P.J.; Runia, W.T.; Hordijk, C.A.; de Boer, W. Legacy Effects of Anaerobic Soil Disinfestation on Soil Bacterial Community Composition and Production of Pathogen-suppressing Volatiles. Front. Microbiol. 2015, 6, 1–12.

