 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Rui M. Gil da Costa | + 6378 word(s) | 6378 | 2021-02-08 04:23:17 | | | |
| 2 | Dean Liu | -3282 word(s) | 3096 | 2021-02-22 05:02:39 | | |
Video Upload Options
Penile cancer is an uncommon and understudied malignancy that is most commonly diagnosed in developing countries. A large proportion of these tumors are caused by infection with high-risk human papillomavirus (HPV), while others are HPV-negative and linked with chronic inflammatory conditions. Relevant cell-based and in vivo animal models are essential to drive research on this type of cancer.
1. Cell-Based Models of Penile Cancer
A number of cell lines representing primary penile squamous cell carcinomas and lymph node metastases have been reported and are summarized in Table 1. The first successful efforts to culture penile cancer cells in vitro were reported in the 1960s[1], and since 2010, a growing number of cell lines—some of which extensively characterized at the molecular and morphological levels—has been described. All of these are able to grow as xenografts in mice (see Table 1), allowing researchers to use those cells for in vivo applications. While the pre-2000 studies do not report the HPV status of cells lines or their original tumors, it is remarkable that all cell lines with defined HPV status are HPV-negative. This lack of representation of HPV-positive disease constitutes a major gap in the array of cell-based models available and additional efforts are needed to develop cell lines representing HPV-positive tumors. The use of these cell-based models for basic and translational research increased in recent years, with multiple publications reporting the use of cell lines developed by Chinese researchers at the Sun Yat-Sen University Cancer Center. Zhou et al. (2018), using a panel of five cell lines, reported their sensitivity to cisplatin and resistance to therapy directed against the epithelial growth factor receptor 1 (EGFR), in association with frequent EGFR amplification[2]. The Penl1 cell line was also used to study the effects of overexpressing inhibitor of DNA binding 1 (ID1), which was found to promote tumor progression[3]. In the same year and using the same cell line, the overexpression of the carcinoembryonic antigen-related cell adhesion molecule 19 (CEACAM19) was found to promote tumorigenesis via activation of small mothers against decapentaplegic (Smad) 2 and 3 and increased metalloproteinase (MMP) 2 and 9 secretion[4]. Conversely, knockdown of the insulin-like growth factor binding protein 2 (IGFBP2) suppressed cell growth, clonogenesis and migration[5][6]. Knockdown of chemokine C-X-C motif ligand 13 (CXCL13) in the Penl1, Penl2, 149RCa and LM156 cell lines suppressed cell proliferation and survival, clonogenesis, migration and invasion via reduced MMP2/9 secretion [7]. These are recent examples of how such well-characterized cell lines can prove to be useful for researchers wishing to explore the biopathology of penile cancer or to test potential new therapies, either using in vitro settings or xenograft models. It is also worth mentioning that the first mouse syngeneic cell-based models were recently reported[8]. These syngeneic models offer an opportunity to study penile cancer cells in a fully immunocompetent murine host and may complement studies done in vitro or with xenografted human cells. The authors established two cell lines from murine penile carcinomas occurring in C57Bl/6 mice that were either Smad4/Apc null (SA1 cells) or Smad4/Apc/Pten null (SAP1 cells). The sensitivity of these cells towards cisplatin and a panel of 42 small molecules selected based on previous proteomic analysis was also tested[8]. This growing array of human and mouse cell-based systems offers opportunities to explore basic and translational aspects of penile cancer and will be essential to drive pre-clinical research in coming years.
Table 1. Penile cancer cell lines and their characteristics.
| Cell Lines | Tissue of Origin | HPV Status | Morphology | Other Characteristics | References Publication Year |
|---|---|---|---|---|---|
| First reported penile cancer cell line | Primary tumor | Not reported | Epithelial | Cytogenetic characterization reported | [1]1966 |
| TSUS-1 | Negative | Not reported | Epithelial | Epithelial morphology, cytogenetic characterization reported, mean doubling time 38 hours | [9] 1983 |
| PCA-5 | Negative | Not reported, human herpesvirus detected | Epithelial | Epithelial morphology, cytogenetic characterization | [10] 1987 |
| KU-8 | Lymph node metastasis | Not reported | Epithelial | Epithelial morphology, cytogenetic characterization reported, mean doubling time 20 hours, EGFR-positive | [11] 1989 |
| Ki-PeCa-L1, Ki-PeCa-P1 | Primary tumor (Ki-PeCa-P1), lymph node metastasis (Ki-PeCa-L1) |
Not reported, positive for p16INK4A | Epithelial | Chemokine profiles available | [12] 2012 |
| P5 | Negative | Negative | Epithelial morphology but sarcomatoid when cultured in vivo | Genomic and transcriptomic characterization | [13] 2016 |
| Penl1, Penl2, 149RM, 149RCa, LM156 | lymph node metastases (Penl1, Penl2, LM156), locally recurrent lesion (149RM) scrotal invasion lesion (149RCa) | Negative | Epithelial | penl2 doubling time: 28 hours, 149RM 26 hours, 149RM and 149RCa 26 hours, LM156 34 hours. All cell lines: genomic characterization available, sensitive to cisplatin, resistant to anti-EGFR therapy | [2][14] 2016, 2018 |
| SA1 | C57Bl/6 mouse primary tumor | Negative | Epithelial | Smad4 and Apc null, cisplatin-sensitive. Genomic, methylation and transcriptomic characterization | [15] 2020 |
| SAP1 | C57Bl/6 mouse primary tumor | Negative | Epithelial | Smad4, Apc and Pten null, cisplatin-resistant. Genomic, methylation and transcriptomic characterization | [8] 2020 |
2. In Vivo Animal Models
The first mouse models for penile cancer were recently reported using either mice transgenic for HPV16[16] or knockout mice for the tumor suppressor genes Smad4 and Adenomatous polyposis coli (Apc), with or without deletion of Phosphatase and tensin homolog (Pten)[8][17]. These models represent major advances for pre-clinical research on penile cancer and will be discussed in detail in the next paragraphs. Table 2 summarizes the main features of the in vivo models for studying penile cancer that were reported so far.
Table 2. In vivo models for HPV-positive and HPV-negative penile cancer.
| Species/Strain | HPV Status | Genetic Modifications | Other Characteristics | Reference Publication Year |
|---|---|---|---|---|
| Horse | HPV status: negative but most are EcPV2-positive. | None. | Spontaneous model. Occurs infrequently in horses. Intraepithelial and pre-malignant lesions: papillomatous lesions. Metastasis: yes, to lymph nodes. | [18] 2014 |
| C57Bl/6 mouse | HPV status: negative. | Based on targeted deletion of Apc/Smad4 with or without Pten deletion. | 100% SCC incidence. Pten deletion confers cisplatin resistance. Intraepithelial and pre-malignant lesions: not described. Metastasis: no. |
[8] 2020 |
| FVB/N mouse | HPV status: Positive for HPV16. | Based on targeted expression of the entire HPV16 early region. | Requires exposure to DMBA.29.6% SCC incidence. Intraepithelial and pre-malignant lesions: yes, condylomas and penile intraepithelial neoplasia. Metastasis: no. |
[16] 2020 |
2.1. Mouse Penis: Anatomy and Histology
The choice of an animal model is based on similarities with human anatomy, physiology and pathology. When considering mouse models of penile cancer, it is necessary to consider similarities and differences between these animals and humans. Keeping this in mind, the next paragraphs and Figure 1A–D present a short comparison between the human and mouse penis, before discussing the new mouse models of penile cancer. As in men, the penis of mice has the main functions of urination, copulation and placement of sperm in the female reproductive tract. In mice, the primordial tubercle that ultimately originates the male penis and the female clitoris is formed between the 12th and 16th gestational days and the differentiation of the female and male organs occurs from the 16th day onwards[19]. After birth, the morphological distinction of female and male external genitalia becomes feasible at approximately 4 weeks of age[19]. The mouse penis consists of a proximal body and distal glans that connect in a right-angle curve [19]. The penile body begins near the pelvic outlet, where the urethra bends at a right angle. At this point, the corpora cavernosa leaves the penile body and diverge laterally to attach to the pubic bone. Thus, the penile body contains the urethra and the right and left cavernous body that will merge in the midline. The penile body ends distally in a right-angle curve, where the penis body joins the glans. The glans contains the glandular spongiosa and cavernosa bodies, the urethral spongiosa body, and the skeletal elements, including a transverse element containing bone and a growth plate of hyaline cartilage and a distal element composed of fibrocartilage that also ossifies after puberty. These skeletal elements allow for the necessary rigidity for a successful mating and account for a major anatomical difference compared with the human penis[20]. The surface of the penile glans is covered by a stratified squamous epithelium showing light keratinization, which is histologically similar to that of the human penis, where most penile squamous cell carcinomas arise. This is an important similarity, as genetic modifications intended to induce squamous cell carcinoma in mouse models are all directed at this epithelial layer. However, this epithelium contains epithelial spines, which constitute another morphological difference when compared with the human penile mucosa. The distal part of the glans is designated the Male Urogenital Mating Bulge (MUMP) [21]. The central element of MUMP is a 1.7 mm long fibrocartilaginous piece. Centrally, the penile bone extends from the right-angle curve of the penis to the MUMP and measures about 3.8 mm in length. In its distal portion, the bone is overlapped dorsally by the MUMP fibrocartilage (Figure 1A,C). The glandular cavernous body is circumferential, while the urethral spongiosa body is linear, runs ventrally to the urethra and appears to be homologous to the corpus spongiosum in humans (Figure 1C–D)[19]. Different to humans, mice have 2 preputial anatomical compartments, the inner covered by smooth non hairy mucosa and the outer covered by hairy skin (Figure 1A–B). The human inner foreskin may be considered an equivalent of the inner mouse foreskin, and the human outer dermal and epidermal foreskin may be equivalent to the mouse outer hairy foreskin. Externalization of the penis in mice can be achieved by applying light abdominal pressure, which allows the glans to leave the external foreskin, extending away from the proximal body. As the glans extends, the inner foreskin protrudes next to it, individualizing the glans, inner foreskin (entirely covered by a keratinizing mucosa) and outer foreskin (covered by haired skin on the external surface)[19]. Human penile anatomy, which is considered to be complex by pathologists, is simpler than that of experimental mice. The glans and inner foreskin surfaces are covered by a stratified squamous epithelium where PeIN develops. Below is the lamina propria, corpus spongiosum or dartos and corpora cavernosa. Tumors progresses vertically along these anatomical levels. In the foreskin, tumors progress from the epithelium to the lamina propria to dartos to dermis to epidermis[20].
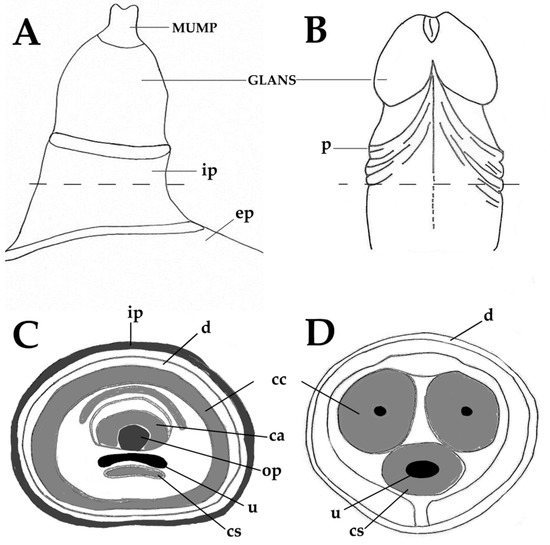
Figure 1. Anatomy of the adult murine (A,C) and human (B,D) penis. Male Urogenital Mating Bulge (MUMP)-male urogenital mating bulge, p-prepuce, ep-external prepuce, ip-internal prepuce, ca-MUMP cartilage, cc-corpus cavernosum, cs-corpus spongiosum, d-dartos, op-os penis, u-urethra.
2.2. HPV-Negative Penile Cancer in SMAD4/APC Double Knockout Mice
Penile squamous cell carcinomas were obtained in C57Bl/6 mice by targeted deletion of the Smad4 and Apc tumor suppressor genes on the penile epithelium. A preliminary report on this model was released in 2017[17] and a full characterization was recently published, revealing its great potential for pre-clinical research[8]. This model was developed using the androgen receptor (AR)-responsive probasin gene promoter, which is commonly used in mouse models of prostate cancer [22] since the penile epithelium is positive for AR[8]. The probasin gene promoter was used to drive the expression of a Cre recombinase and delete Smad4 and Apc in PB-Cre4+ Smad4L/L and/or ApcL/L mice. The deletion of each gene individually was insufficient to induce cancer, but their co-deletion was found to induce squamous cell carcinoma of the penis at 100% penetrance (median age 17.2 weeks-old). Although the tumors were initially induced via AR-driven mutagenesis, they were found to be AR-independent, and tumor progression was unaffected by castration. Transcriptomic analysis of these lesions confirmed the deregulation of the Wnt/β-catenin and the fibroblast growth factor (FGF) pathways induced by their genetic alterations. Importantly, the penile tumors transcriptome also revealed marked pro-inflammatory signaling pivoted by cyclooxigenase-2 (COX-2) and massive infiltration by immunosuppressive myeloid cells. This animal model was used to test the efficacy of rational drug combinations based on immune checkpoint inhibitors (anti-PD1/anti-CTLA4 antibodies) combined with either the selective COX-2 inhibitor celecoxib or with the multi-target tyrosine kinase inhibitor cabozantinib. The drug combinations proved to be significantly more effective than each drug alone and their anti-tumor effects were associated with reduced infiltration of myeloid cells and regulatory T lymphocytes. These results highlight the potential of this model for developing innovative combination therapies, especially those involving immune checkpoints blockade. Interestingly, celecoxib was also shown to increase the activation of cytotoxic T lymphocytes in a different model of epithelial carcinogenesis induced by HPV16, further supporting the use of COX-2 inhibition for potentiating immunological therapies[23]. Considering the role of Pten in the resistance of penile cancer to platinum-based therapy, the authors also developed a triple knockout, Smad4/Apc/Pten null mouse model. While the double knockout tumors remained sensitive to cisplatin, the triple knockout lesions were largely resistant, corroborating the role of Pten loss in driving resistance to platinum-based therapies in human patients. These findings validate those mouse models for studying strategies to overcome cisplatin resistance. The syngeneic cell lines developed from both the double and the triple knockout models are equally powerful tools for translational studies. Importantly, the authors reported that orthotopic injection of these cells lines was able to replicate penile squamous cell carcinoma. This is more realistic than implanting cell-based models heterotopically (most often subcutaneously) and should allow for drug development as well as for identifying key mechanisms involved in the invasion of penile structures. One important caution for researchers working in this field is that these models seem more adequate for representing HPV-negative disease than HPV-positive disease. Although the authors present their mouse models in the context of HPV-positive penile cancer and highlight similarities with HPV-positive disease, several key differences exist: (1) the absence of HPV oncogenes in these mouse models; (2) one key HPV target, pRb, remains present and phosphorylated at high levels; and (3) the gene mutations employed to generate this mouse models are found in HPV-positive and negative tumors and in many other epithelial neoplasms[24][25]. Overall, the models by Huang et al. (2020) are very promising tools to study HPV-negative disease and should be used in combination with other models that can adequately mimic HPV-positive lesions[8].
2.3. In Vivo Models for HPV-Positive Penile Cancer
An ideal model for studying HPV-positive penile cancer would be driven by key oncogenes from high-risk HPVs (e.g., HPV16 E6 and E7), would develop lesions specifically at the penis and would reproduce the main morphological and molecular features of HPV-induced carcinogenesis (e.g., basaloid PeIN, HPV-associated SCC subtypes followed by lymph node metastasis)[26]. Parts of these aims have been recently achieved by different research teams using complementary models (Table 2). A recently reported mouse model for HPV-positive penile cancer employs mice carrying the whole HPV16 early region (containing the key oncogenes E5, E6 and E7) in an FVB/N genetic background[16]. In this model, the expression of HPV oncogenes is targeted to basal keratinocytes by the cytokeratin 14 (Krt14) gene promoter[27][28] and the mouse strain is often referred to as K14-HPV16. This mouse strain was originally developed in the 1990s[27][28] and other related strains, carrying only the E6 and or E7 oncogenes, were used to study a number of HPV-induced cancers, including cervical cancer[27] anal cancer[29] oral, esophageal and oropharyngeal cancers[30][31]. These mice are also useful for studying interactions between HPV and the immune system—as they are fully immunocompetent—as well as with hormonal and other microenvironmental factors and with potential chemical co-carcinogens (e.g., tobacco toxins)[23][32][33]. One limitation of this model system is the fact that expression of HPV oncogenes is not regulated by the viral long control region or by cellular mechanisms normally involved in naturally infected patients, instead is driven by the Krt14 gene promoter[27][28]. Another limitation is the widespread occurrence of intraepithelial hyperplastic and dysplastic lesions throughout the skin and keratinizing mucosae, which may reduce the specificity of models intended solely for penile cancer[16]. This difficulty may be partially overcome by obtaining syngeneic cell lines that can be injected orthotopically or heterotopically into matched FVB/N HPV-negative mice. Finally, HPV-transgenic mice are not good models to study the mechanisms of HPV cell entry and infection. Models based on natural infection by the murine papillomavirus (MmuPV1) are a promising alternative in this regard, but no penile lesions caused by mmuPV1 have been reported so far[6][34]. HPV16-transgenic mice were found to develop PeIN and condylomas at 30 weeks-old, but no squamous cell carcinomas[16]. These were obtained at a 29.6% incidence by exposing HPV16-transgenic mice to the tobacco-related carcinogen dimethylbenz(a)anthracene (DMBA) topically in the penile mucosa weekly, for 16 weeks. Importantly, the presence of the HPV oncogenes was associated with deregulated cell proliferation, as assessed immunohistochemically using Ki-67, a marker for proliferative cells. This protein is present in the G1, S, G2 and mitotic phases of the cell cycle, and is used to infer the growth fraction of a given cell population[35][36][37]. The intraepithelial and invasive lesions observed in this animal model closely mimicked the histological features of HPV-positive penile lesions in human patients. PeIN lesions predominantly showed basaloid morphology and koilocytosis, typical of HPV-induced pathology. Invasive squamous cell carcinomas also corresponded to histological subtypes usually associated with HPV positivity like basaloid, warty-basaloid and solid medullary-like tumors[26][38][39][40]. DMBA application in this model was not associated with increased incidence of SCC on cutaneous locations as assessed histologically nor with systemic genotoxicity, according to screens using the micronucleus and the comet assays. Importantly, these observations provided the first experimental demonstration of the etiological role of HPV16 in penile cancer. Additionally, this study also supports the role of tobacco toxins as important promoters of penile carcinogenesis. While this model reproduced the whole spectrum of HPV-induced penile lesions, no metastases were observed, limiting its usefulness to study the more advanced stages of the disease. Another interesting model for studying penile cancer induced by papillomaviruses may be found in horses. These animals develop penile papillomas, intraepithelial neoplastic lesions and squamous cell carcinomas which are associated with infection by equine papillomavirus type 2 (EcPV2)[18][41][42][43]. Importantly, some equine penile squamous cell carcinomas are able to metastasize to regional lymph nodes, something that has not yet been observed when studying the newly available mouse models[18]. Interestingly, the characterization of these tumors yielded some markers that are similar to those observed in human patients[44]. Suárez–Bonnet et al. (2018) found that COX-2 was neo-expressed in 86% of cases, and the expression was higher in squamous cell carcinomas than in papillomas[44]. In squamous cell carcinomas, E-cadherin was present in 65% of cases, and vimentin was neo-expressed in 65% of poorly differentiated cases[44]. The cytoplasmic expression of 14-3-3σ protein was observed in 42% of squamous cell carcinomas[44]. Pten expression tended to be decreased or lost in squamous cell carcinomas[44], which correlates with cisplatin resistance in human patients and in another animal model, as previously discussed[8]. MMP1 is also expressed in equine penile lesions[43], which is in line with the expression of MMPs reported in human and murine penile[8]. These lesions also show nuclear p53 accumulation, revealed by immunohistochemical techniques, suggesting that p53 gene mutations are common despite a papillomaviral etiology[18]. This may be a specificity of the equine model and deserves further elucidation. Taken together, these observations suggest that equine penile lesions may be a useful spontaneous model of penile neoplasia, from which there is much to be learned.
References
- Yamane, I.; Tsuda, T. Establishment of a cell line in vitro from the lesion of a clinical case of penis cancroid. Tohoku J. Exp. Med. 1966, 88, 9–20.
- Zhou, Q.H.; Deng, C.Z.; Li, Z.S.; Chen, J.P.; Yao, K.; Huang, K.B.; Liu, T.Y.; Liu, Z.W.; Qin, Z.K.; Zhou, F.J.; et al. Molecular characterization and integrative genomic analysis of a panel of newly established penile cancer cell lines. Cell Death Dis. 2018, 9, 684.
- Hu, X.; Chen, M.; Li, Y.; Wang, Y.; Wen, S.; Jun, F. Overexpression of ID1 promotes tumor progression in penile squamous cell carcinoma. Oncol. Rep. 2019, 41, 1091–1100.
- Hu, X.; Chen, M.; Li, Y.; Wang, Y.; Wen, S.; Jun, F. Aberrant CEACAM19 expression is associated with metastatic phenotype in penile cancer. Cancer Manag. Res. 2019, 11, 715–725.
- Hu, X.; Chen, M.; Liu, W.; Li, Y.; Fu, J. Preoperative plasma IGFBP2 is associated with nodal metastasis in patients with penile squamous cell carcinoma. Urol. Oncol. 2019, 37, 452–461.
- Ingle, A.; Ghim, S.; Joh, J.; Chepkoech, I.; Bennett Jenson, A.; Sundberg, J.P. Novel laboratory mouse papillomavirus (MusPV) infection. Vet. Pathol. 2011, 48, 500–505.
- Mo, M.; Tong, S.; Li, T.; Zu, X.; Hu, X. Serum CXCL13 Level is Associated with Tumor Progression and Unfavorable Prognosis in Penile Cancer. OncoTargets Ther. 2020, 13, 8757–8769.
- Huang, T.; Cheng, X.; Chahoud, J.; Sarhan, A.; Tamboli, P.; Rao, P.; Guo, M.; Manyam, G.; Zhang, L.; Xiang, Y.; et al. Effective combinatorial immunotherapy for penile squamous cell carcinoma. Nat. Commun. 2020, 11, 2124.
- Ishikawa, S.; Kanoh, S.; Nemoto, S. Establishment of a cell line (TSUS-1) derived from a human squamous cell carcinoma of the penis. Hinyokika Kiyo 1983, 29, 373–376.
- Gentile, G.; Giraldo, G.; Stabile, M.; Beth-Giraldo, E.; Lonardo, F.; Kyalwazi, S.K.; Perone, L.; Ventruto, V. Cytogenetic study of a cell line of human penile cancer. Ann. Genet. 1987, 30, 164–169.
- Tsukamoto, T. Establishment and characterization of a cell line (KU-8) from squamous cell carcinoma of the penis. Keio J. Med. 1989, 38, 277–293.
- Naumann, C.M.; Sperveslage, J.; Hamann, M.F.; Leuschner, I.; Weder, L.; Al-Najar, A.A.; Lemke, J.; Sipos, B.; Junemann, K.P.; Kalthoff, H. Establishment and characterization of primary cell lines of squamous cell carcinoma of the penis and its metastasis. J. Urol. 2012, 187, 2236–2242.
- Munoz, J.J.; Drigo, S.A.; Kuasne, H.; Villacis, R.A.; Marchi, F.A.; Domingues, M.A.; Lopes, A.; Santos, T.G.; Rogatto, S.R. A comprehensive characterization of cell cultures and xenografts derived from a human verrucous penile carcinoma. Tumor Biol. 2016, 37, 11375–11384.
- Chen, J.; Yao, K.; Li, Z.; Deng, C.; Wang, L.; Yu, X.; Liang, P.; Xie, Q.; Chen, P.; Qin, Z.; et al. Establishment and characterization of a penile cancer cell line, penl1, with a deleterious TP53 mutation as a paradigm of HPV-negative penile carcinogenesis. Oncotarget 2016, 7, 51687–51698.
- Bravo, I.G.; Felez-Sanchez, M. Papillomaviruses: Viral evolution, cancer and evolutionary medicine. Evol. Med. Public Health 2015, 2015, 32–51.
- Medeiros-Fonseca, B.; Mestre, V.F.; Estevao, D.; Sanchez, D.F.; Canete-Portillo, S.; Fernandez-Nestosa, M.J.; Casaca, F.; Silva, S.; Brito, H.; Felix, A.; et al. HPV16 induces penile intraepithelial neoplasia and squamous cell carcinoma in transgenic mice: First mouse model for HPV-related penile cancer. J. Pathol. 2020, 251, 411–419.
- Sarhan, A.; Shang, X.; Tamboli, P.; Rao, P.; Pettaway, C.; Wang, A.; DePinho, R.; Lu, X. MP81-07 penile carcinoma: Genetically engineered models for novel therapeutics identification. J. Urol. 2017, 197, e1091.
- van den Top, J.G.; Harkema, L.; Lange, C.; Ensink, J.M.; van de Lest, C.H.; Barneveld, A.; van Weeren, P.R.; Grone, A.; Martens, A. Expression of p53, Ki67, EcPV2- and EcPV3 DNA, and viral genes in relation to metastasis and outcome in equine penile and preputial squamous cell carcinoma. Equine Vet. J. 2015, 47, 188–195.
- Rodriguez, E., Jr.; Weiss, D.A.; Yang, J.H.; Menshenina, J.; Ferretti, M.; Cunha, T.J.; Barcellos, D.; Chan, L.Y.; Risbridger, G.; Cunha, G.R.; et al. New insights on the morphology of adult mouse penis. Biol. Reprod. 2011, 85, 1216–1221.
- Cubilla, A.L.; Lloveras, B.; Alejo, M.; Clavero, O.; Chaux, A.; Kasamatsu, E.; Monfulleda, N.; Tous, S.; Alemany, L.; Klaustermeier, J.; et al. Value of p16(INK)(4)(a) in the pathology of invasive penile squamous cell carcinomas: A report of 202 cases. Am. J. Surg. Pathol. 2011, 35, 253–261.
- Phillips, T.R.; Wright, D.K.; Gradie, P.E.; Johnston, L.A.; Pask, A.J. A Comprehensive Atlas of the Adult Mouse Penis. Sex. Dev. 2015, 9, 162–172.
- Grabowska, M.M.; DeGraff, D.J.; Yu, X.; Jin, R.J.; Chen, Z.; Borowsky, A.D.; Matusik, R.J. Mouse models of prostate cancer: Picking the best model for the question. Cancer Metastasis Rev. 2014, 33, 377–397.
- Santos, C.; Neto, T.; Ferreirinha, P.; Sousa, H.; Ribeiro, J.; Bastos, M.; Faustino-Rocha, A.I.; Oliveira, P.A.; Medeiros, R.; Vilanova, M.; et al. Celecoxib promotes degranulation of CD8(+) T cells in HPV-induced lesions of K14-HPV16 transgenic mice. Life Sci. 2016, 157, 67–73.
- Polakis, P. The many ways of Wnt in cancer. Curr. Opin. Genet. Dev. 2007, 17, 45–51.
- Arya, M.; Thrasivoulou, C.; Henrique, R.; Millar, M.; Hamblin, R.; Davda, R.; Aare, K.; Masters, J.R.; Thomson, C.; Muneer, A.; et al. Targets of Wnt/ss-catenin transcription in penile carcinoma. PLoS ONE 2015, 10, e0124395.
- Cubilla, A.L.; Velazquez, E.F.; Amin, M.B.; Epstein, J.; Berney, D.M.; Corbishley, C.M.; Members of the ISUP Penile Tumor Panel. The World Health Organisation 2016 classification of penile carcinomas: A review and update from the International Society of Urological Pathology expert-driven recommendations. Histopathology 2018, 72, 893–904.
- Arbeit, J.M.; Munger, K.; Howley, P.M.; Hanahan, D. Progressive squamous epithelial neoplasia in K14-human papillomavirus type 16 transgenic mice. J. Virol. 1994, 68, 4358–4368.
- Coussens, L.M.; Hanahan, D.; Arbeit, J.M. Genetic predisposition and parameters of malignant progression in K14-HPV16 transgenic mice. Am. J. Pathol. 1996, 149, 1899–1917.
- Stelzer, M.K.; Pitot, H.C.; Liem, A.; Schweizer, J.; Mahoney, C.; Lambert, P.F. A mouse model for human anal cancer. Cancer Prev. Res. (Phila.) 2010, 3, 1534–1541.
- Strati, K.; Pitot, H.C.; Lambert, P.F. Identification of biomarkers that distinguish human papillomavirus (HPV)-positive versus HPV-negative head and neck cancers in a mouse model. Proc. Natl. Acad. Sci. USA 2006, 103, 14152–14157.
- Mestre, V.F.; Medeiros-Fonseca, B.; Estevao, D.; Casaca, F.; Silva, S.; Felix, A.; Silva, F.; Colaco, B.; Seixas, F.; Bastos, M.M.; et al. HPV16 is sufficient to induce squamous cell carcinoma specifically in the tongue base in transgenic mice. J. Pathol. 2020, 251, 4–11.
- Paiva, I.; Gil da Costa, R.M.; Ribeiro, J.; Sousa, H.; Bastos, M.M.; Faustino-Rocha, A.; Lopes, C.; Oliveira, P.A.; Medeiros, R. MicroRNA-21 expression and susceptibility to HPV-induced carcinogenesis—role of microenvironment in K14-HPV16 mice model. Life Sci. 2015, 128, 8–14.
- Santos, C.; Ferreirinha, P.; Sousa, H.; Ribeiro, J.; Bastos, M.M.; Neto, T.; Oliveira, P.A.; Medeiros, R.; Vilanova, M.; Gil da Costa, R.M. Ptaquiloside from bracken (Pteridium spp.) inhibits tumour-infiltrating CD8(+) T cells in HPV-16 transgenic mice. Food Chem. Toxicol. 2016, 97, 277–285.
- Cladel, N.M.; Budgeon, L.R.; Balogh, K.K.; Cooper, T.K.; Hu, J.; Christensen, N.D. Mouse papillomavirus MmuPV1 infects oral mucosa and preferentially targets the base of the tongue. Virology 2016, 488, 73–80.
- Diorio, G.J.; Leone, A.R.; Spiess, P.E. Management of Penile Cancer. Urology 2016, 96, 15–21.
- Scholzen, T.; Gerdes, J. The Ki-67 protein: From the known and the unknown. J. Cell. Physiol. 2000, 182, 311–322.
- Berdjis, N.; Meye, A.; Nippgen, J.; Dittert, D.; Hakenberg, O.; Baretton, G.B.; Wirth, M.P. Expression of Ki-67 in squamous cell carcinoma of the penis. BJU Int. 2005, 96, 146–148.
- Sanchez, D.F.; Rodriguez, I.M.; Piris, A.; Canete, S.; Lezcano, C.; Velazquez, E.F.; Fernandez-Nestosa, M.J.; Mendez-Pena, J.E.; Hoang, M.P.; Cubilla, A.L. Clear Cell Carcinoma of the Penis: An HPV-related Variant of Squamous Cell Carcinoma: A Report of 3 Cases. Am. J. Surg. Pathol. 2016, 40, 917–922.
- Chaux, A.; Reuter, V.; Lezcano, C.; Velazquez, E.; Codas, R.; Cubilla, A.L. Autopsy findings in 14 patients with penile squamous cell carcinoma. Int. J. Surg. Pathol. 2011, 19, 164–169.
- Knight, C.G.; Munday, J.S.; Peters, J.; Dunowska, M. Equine penile squamous cell carcinomas are associated with the presence of equine papillomavirus type 2 DNA sequences. Vet. Pathol. 2011, 48, 1190–1194.
- Arthurs, C.; Suarez-Bonnet, A.; Willis, C.; Xie, B.; Machulla, N.; Mair, T.S.; Cao, K.; Millar, M.; Thrasivoulou, C.; Priestnall, S.L.; et al. Equine penile squamous cell carcinoma: Expression of biomarker proteins and EcPV2. Sci. Rep. 2020, 10, 7863.
- Sykora, S.; Jindra, C.; Hofer, M.; Steinborn, R.; Brandt, S. Equine papillomavirus type 2: An equine equivalent to human papillomavirus 16? Vet. J. 2017, 225, 3–8.
- Ramsauer, A.S.; Kubacki, J.; Favrot, C.; Ackermann, M.; Fraefel, C.; Tobler, K. RNA-seq analysis in equine papillomavirus type 2-positive carcinomas identifies affected pathways and potential cancer markers as well as viral gene expression and splicing events. J. Gen. Virol. 2019, 100, 985–998.
- Suarez-Bonnet, A.; Willis, C.; Pittaway, R.; Smith, K.; Mair, T.; Priestnall, S.L. Molecular carcinogenesis in equine penile cancer: A potential animal model for human penile cancer. Urol. Oncol. 2018, 36, 532.e9–532.e18.

