 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Tina Tootle | + 3940 word(s) | 3940 | 2021-01-14 07:19:19 | | | |
| 2 | Camila Xu | Meta information modification | 3940 | 2021-01-21 05:14:56 | | |
Video Upload Options
Fascin, an actin-binding protein, regulates many developmental migrations and contributes to cancer metastasis.
1. Introduction
Fascin is an actin-binding protein that promotes different modes of cell migration, including during embryonic development and cancer metastasis [1][2]. One means by which Fascin promotes migration is by crosslinking or bundling actin filaments together [3][4]. Through this function, Fascin aids in the formation of many actin-rich migratory structures such as filopodia [5][6][7], lamellipodia [5], stress fibers [8], and microspikes [9][10]. Fascin also localizes to actin-rich adhesive structures such as focal adhesions [8], invadopodia [11], and podocytes [12][13]. Due to its roles in cell migration, Fascin has frequently been studied in the context of cancer. Fascin is absent in adult epithelial tissue [14] but is highly expressed in many types of carcinomas. High Fascin expression is correlated with poor prognosis and increased metastasis (reviewed in [4]). For this reason, Fascin is a prognostic biomarker and a potential therapeutic target of metastatic carcinomas [2][4]. While studies on Fascin have centered around Fascin’s canonical actin bundling function, Fascin also has non-canonical, non-bundling functions in the cell. In this review, we discuss Fascin’s structure and expression, explore how both the canonical and non-canonical functions of Fascin promote migration, examine the various mechanisms regulating Fascin, and consider how the different functions of Fascin may contribute to diseases.
2. Fascin Structure, Expression, and Functions
Fascin promotes cell migration not only through its conserved actin bundling activity, but also by its non-canonical functions. In this section, we review the structure of Fascin, how it bundles actin, and Fascin’s expression pattern during development and in adult tissues. Then, we discuss how Fascin promotes cell migration through both its canonical actin bundling function and non-canonical functions: modulating the activity of other actin-binding proteins [8][15][16], binding to microtubules [17], interacting with the Linker of the Nucleoskeleton and Cytoskeleton (LINC) Complex [18], and having nuclear roles [19].
2.1. Structure of Fascin
Fascin was initially discovered in sea urchins [20] and later found in Drosophila [21][22], Xenopus [23], mouse [24], and humans [25][26]. Fascin is a globular protein of approximately 55kDa and is comprised of four β-trefoil domains (Figure 1) [26]. Each β-trefoil domain contains six two-stranded β-hairpins oriented in a trifold symmetry [27][28]. The four β-trefoils of Fascin are arranged into two lobes (Figure 1B) [26]. This structure allows a monomer of Fascin to bundle actin, while most actin bundlers function as dimers (Figure 1A). Fascin has three distinct surface areas able to bind actin; two larger actin-binding areas are in the clefts of β-trefoils 1 and 2, and β-trefoils 1 and 4, while a third smaller area is in β-trefoil 3 (Figure 1A,B) [29][30]. Based on cryoEM analysis of actin bundled by Fascin in filopodia, the two larger actin-binding areas (referred to as actin-binding site 1) bind to a single actin filament, while the actin-binding area in β-trefoil 3 (referred to as actin-binding site 2) binds to a second actin filament. The two sites of actin bundling are approximately 5 nm apart, which along with Fascin being monomeric, causes Fascin to be the tightest actin bundler with approximately 8.1 nm between actin filaments [29][31]. Recent X-ray crystallography, along with the systematic mutagenesis of Fascin, reveals that mutation in one of the actin-binding sites impairs the other site’s actin-binding activity, suggesting there is coordinated regulation between Fascin’s actin-binding sites [29].
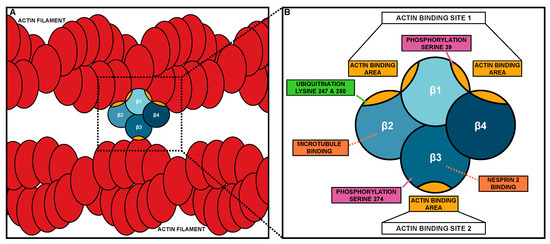
Figure 1. Fascin bundling mechanism and actin-binding domains. (A) Schematic of Fascin bundling two actin filaments (red). (B) Schematic of the domains and binding site of Fascin. The four β-trefoil domains of Fascin are in different shades of blue. Actin-binding areas of Fascin are in gold. In B, the different protein interaction sites and post-translational modifications are labeled.
2.2. Expression and Bundling Function of Fascin
Humans and other vertebrates have three Fascin isoforms (Fascin-1, Fascin-2, and Fascin-3) that each have distinct expression patterns and functions. Fascin-1 (FSCN1) is the most widely studied and is expressed in mesenchymal and nervous tissues during development [1]. Fascin-2 has 56% homology to Fascin-1 and is expressed in the inner and outer segments of photoreceptors and in the stereocilia of the hair cells [32][33][34], where it bundles actin to stabilize the protrusive structures these cells form [32][33]. Moreover, mutations in Fascin-2 are associated with retinitis pigmentosa and hearing loss [33][35][36]. Fascin-3 has 29% homology to Fascin-1 and is expressed in the testis and developing spermatozoa, where it plays a role in the terminal elongation of the spermatid head [37]. Interestingly, the area surrounding actin-binding site 1 is highly conserved across all three Fascins [37]. Thus, the different isoforms of Fascin bundle actin in different tissues and structures.
During mouse embryonic development, Fascin-1 (subsequently referred to as Fascin) is widely expressed [1][14][38]. Indeed, Fascin is expressed in the central and peripheral nervous system, the neuroepithelium, developing somites, and the mesenchyme of limb buds [1]. While Fascin is expressed in a wide variety of tissues, it is largely dispensable for mouse development. Fascin knockout mice are not embryonic lethal but have increased neonatal lethality [38]. These mice display defects in the central nervous system, including a notable enlargement of the lateral ventricles [38]. The neonatal lethality is attributed to poor feeding and breathing after birth, but the consequences of the other abnormalities are not well understood [38]. Loss of Fascin in these mice is potentially compensated for by the other two Fascins (Fascin-2 and Fascin-3); however, a triple knockout mouse model has not been created. While Fascin is largely dispensable for mouse embryonic development, its expression patterns are largely conserved in human tissues.
In humans, Fascin is more widely expressed during development than in the adult. While Fascin is largely absent from adult epithelial tissues, Fascin is expressed in neurons, the glomerulus of the kidney, the adrenal gland, the basal layer of the skin, and immune cells of adults [14]. In particular, Fascin is highly expressed during dendritic cell maturation [39]. Dendritic cells are highly motile cells and play an important role in innate and adaptive immunity [39]. Loss of Fascin impairs dendritic cell maturation and therefore motility [39]. During embryonic and fetal development, Fascin is widely expressed in the nervous system, including neuroblasts, neural crest cells, melanoblasts, mesenchymal tissues, microcapillary endothelial cells, gastrointestinal tract, and antigen-presenting dendritic cells [14][40][41][42][43][44]. A common feature of many of these cell types is that they undergo significant cell migrations, highlighting the importance of Fascin in promoting cell migration. For example, Fascin-expressing neuroblasts of the subventricular zone in the postnatal brain migrate along the rostral migratory stream to the olfactory bulb [44].
The role of Fascin in migration is conserved across organisms. Indeed, even in lower eukaryotes, Fascin-expressing cells undergo significant migrations. For example, in sea urchins, Fascin is expressed in coelomocytes [45]. Coelomocytes are migratory phagocytic leukocytes that perform essential immune functions [45]. In these cells, Fascin bundles actin to promote filopodia formation and migration [45]. In zebrafish, Fascin promotes migration by forming filopodia in a subset of neural crest cells [40]. In Drosophila, the macrophage-like hemocytes that migrate throughout the developing embryo highly express Fascin, which promotes their migration through filopodia formation [46]. Fascin is also highly expressed in the migratory border cells of the Drosophila ovary [22] and is essential for their on-time collective migration [47]. Specifically, Fascin promotes both delamination of the border cell cluster from the epithelium and the proper formation of protrusions [47]. These examples highlight the conservation of Fascin in promoting cell migration across organisms.
2.3. Non-Canonical Roles of Fascin
While Fascin’s role in promoting migration, especially in the context of cancer metastasis, is primarily attributed to its function as an actin bundler, research has uncovered many other functions of Fascin. These new functions not only demonstrate that Fascin promotes cell migration through multiple mechanisms and pathways (Figure 2) but are new potential therapeutic targets to prevent cancer metastasis.
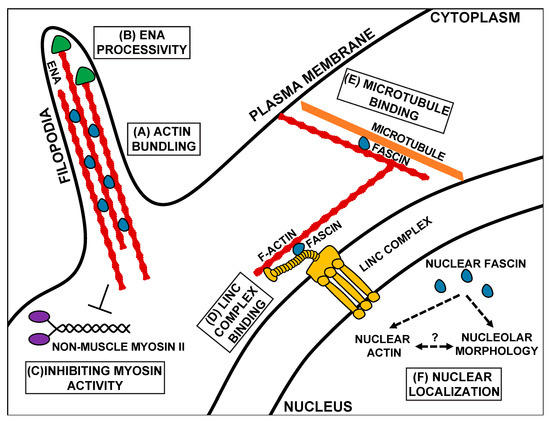
Figure 2. Schematic of the canonical and non-canonical functions of Fascin. Canonically, Fascin (blue) bundles actin filaments (red) to form structures such as filopodia (A). Fascin also regulates other actin-binding proteins, such as increasing the processivity of the actin elongating factor, Ena (green, B), and inhibiting Non-muscle Myosin II (purple) activity (C). At the nuclear periphery, Fascin mediates mechanotransduction by interacting with the Linker of the Nucleoskeleton and Cytoskeleton (LINC) complex (gold, D). In addition to binding actin, Fascin also binds to microtubules (orange, E). Fascin also localizes to the nucleus, where it likely regulates nuclear actin and nucleolar morphology (F). Schematic is not drawn to scale.
In addition to forming cellular protrusions and extensions, the actin bundling function of Fascin regulates other actin-binding proteins, including Enabled (Ena) and Non-muscle Myosin II (Figure 2B,C). Ena is an actin elongation factor that binds to the plus end of actin filaments and promotes polymerization [48]. When actin filaments are bundled in vitro by either human or Drosophila Fascin (Singed), Ena has increased processivity, meaning it stays associated with and polymerizes actin filaments for longer periods of time (Figure 2B) [15][16]. This increased processivity is not observed with other actin bundlers [15][16]. Similarly, during Drosophila border cell migration, Fascin and Ena genetically interact to promote on-time migration [47]. This interaction suggests that Fascin increases Ena processivity in the border cells to mediate on-time migration, potentially by promoting protrusion formation in the cluster [47]. In addition to regulating Ena, Fascin also regulates Non-muscle Myosin II activity in vitro [8] (subsequently referred to as Myosin II). Specifically, Fascin limits Myosin II’s ATP consumption and decreases Myosin II’s motor activity (Figure 2C) [8]. This regulation may occur by Fascin bundling actin filaments and precluding Myosin II’s ability to bind to the filaments [8]. Since Myosin II can alter both cell contractility and cell stiffness, it is an essential component in both migrating cells and their substrate [49]. Thus, Fascin’s actin bundling activity promotes migration by not only by mediating the formation of cell migratory structures but also by modulating the activity of actin-binding proteins.
Fascin’s bundling activity also controls actin filaments outside of cell migratory structures. For example, Fascin mediates the remodeling of mitochondrial actin filaments to control mitochondrial oxidative phosphorylation. In lung adenocarcinomas, Fascin stabilizes filamentous actin around mitochondria, which regulates mitochondrial fission and fusion as well as promotes metastasis [50].
Another means by which Fascin contributes to migration is by binding to and regulating microtubules [17]. Microtubules directly interact with Fascin via β-trefoil 2, which likely blocks Fascin’s actin-binding site 1 (Figure 1B or Figure 2E) [17]. Fascin’s interaction with microtubules promotes cell migration by regulating microtubule dynamics [17]. Blocking the microtubule–Fascin interaction impairs in vitro cell migration by increasing the size and reducing the turnover of focal adhesions [17]. Fascin promotes microtubule-dependent focal adhesion disassembly, and therefore cell migration, by activating the FAK-Src pathway [17]. These data reveal that Fascin interacts with microtubules to regulate focal adhesions to promote cell migration.
Fascin also localizes to the outside of the nucleus in Drosophila nurse cells and mammalian cells [19]. This perinuclear localization is reminiscent of the LINC Complex, which is a key mediator of mechanotransduction [51][52]. The LINC Complex is composed of KASH (Klarischt, ANC-1, Syne Homology) and SUN (Sad1p, UNC-84) proteins. KASH proteins interact with cytoskeletal filaments in the cytoplasm and extend through the outer nuclear envelope to connect to SUN proteins. SUN proteins extend from the inner nuclear envelope into the nucleus, where they bind to the nuclear lamina [51][52]. This structure allows the transmission force from the outside of the cell, through the cytoskeleton, to the nucleus and regulates the nuclear shape and position during cell migration [51][52][53][54]. Fascin binds directly to the KASH protein, Nesprin 2, through β-trefoil 3 (Figure 1B or Figure 2D). This interaction allows Fascin to bind to the LINC Complex and to actin through its actin-binding site 1, thus connecting the LINC Complex to the actin cytoskeleton [18]. While Nesprins can directly bind to actin filaments, interaction with Fascin may aid in strengthening this connection, particularly for invasive migrations [18]. Disruption in this interaction leads to significant impairments in nuclear deformation during single-cell invasive migrations [18]. This work suggests that in addition to bundling actin, Fascin plays a key role in regulating mechanotransduction, which is critical for cell migration.
Fascin may also contribute to cell migration independent of its interaction with cytoskeletal filaments. Indeed, while Fascin primarily localizes in the cytoplasm, it also localizes to the nucleus (Figure 2F) [19]. This nuclear localization is likely conserved across organisms, as it is observed in both Drosophila nurse cells and mammalian cell lines [19]. While the functions of nuclear Fascin remain poorly understood, it may regulate nuclear actin. Indeed, complete loss of Fascin reduces, while an overexpression of Fascin increases the frequency of Drosophila nurse cells with high levels of one form of nuclear actin [55]. Changes in nuclear actin regulation can alter gene expression (reviewed in [56]), which can impinge upon processes such as cell migration. Thus, it is tempting to speculate that nuclear Fascin could promote cell migration by regulating nuclear actin and, thereby, transcription factor activity and gene expression. For example, nuclear actin regulates transcription by modulating several transcription factors, such as myocardin-related transcription factor A (MRTF, also referred to as megakaryocytic acute leukemia (MAL)). Specifically, nuclear actin regulates MRTF’s nuclear localization and activity [57]. MRTF promotes many types of cell migration [58]. Another potential role of nuclear Fascin is to regulate the functions of the nucleolus [19]. The nucleolus forms by phase separation, meaning that its function is tightly associated with its structure. Indeed, disruption of its functions results in severe alteration in its morphology and vice versa [59]. Fascin promotes proper nucleolar morphology in both Drosophila nurse cells and mammalian cell lines [19]. In addition to producing ribosomes, the nucleolus is an essential metabolic regulator and responder to cellular stress, both of which can impinge upon cell migration. Interestingly, nuclear actin localizes to the nucleolus and regulates nucleolar functions [60]; these data suggest that Fascin may regulate nuclear actin to influence the nucleolar structure and function. Supporting a nucleolar role for Fascin, a proteomic analysis of Fascin-interacting proteins in laryngeal squamous cell carcinomas identified several nucleolar interacting partners involved in ribosomal RNA processing [61]. Lastly, nuclear Fascin interacts with a histone methyltransferase subunit, RbBP5, to promote histone3 Lys4 trimethylation at target gene promoter regions [62]. Thus, one nuclear function of Fascin is to regulate histone modifications and transcription [62]. Together, these findings highlight how Fascin has diverse and poorly understood nuclear functions and that further studies are needed to determine how nuclear Fascin functions during cell migration.
Overall, Fascin is a highly conserved regulator of cell migration. Fascin is the tightest actin bundler in the cell and mediates the formation of cell migratory structures, such as filopodia. In addition to bundling actin, Fascin has many non-canonical functions, including modulating the activity of actin-binding proteins [8][15][16], regulating microtubule dynamics [17], mediating mechanotransduction via the LINC Complex [18], and acting within the nucleus [19]. Therefore, Fascin influences cell migration through its canonical actin bundling activity, but it likely also promotes cell migration through one or more of these other functions.
3. Regulation of Fascin
Since Fascin has multiple functions, tight regulation of these functions is necessary to ensure cell migration. Fascin is regulated by post-translational modifications, upstream pathways, protein–protein interactions, and transcriptional means. However, very little is known about how these different regulatory mechanisms coordinate the different functions of Fascin. This section discusses the mechanisms regulating Fascin and how they influence both Fascin’s actin bundling and non-canonical functions.
3.1. Post-Translational Modifications
The most well-known mechanism of regulating Fascin is through phosphorylation. Fascin phosphorylation has been investigated both in human cell lines and Drosophila. Fascin has two known phosphorylation sites. One site is serine 39 (S39, Drosophila S52), which is in actin-binding site 1 on β-trefoil 1 (Figure 1B). Thus, it is not surprising that phosphorylation at S39 inhibits Fascin’s actin bundling function [63][64]. Protein Kinase C (PKC) phosphorylates Fascin at S39, and this phosphorylation promotes Fascin binding to PKC; this is observed both in vitro and in vivo [65]. The other phosphorylation site was first identified in Drosophila (S289) but is conserved in humans (S274) [66]. This site is on β-trefoil 3 near actin-binding site 2 (Figure 1B). Both the phospho-mutant and phospho-mimetic of S289 in Drosophila impair Fascin’s actin bundling activity in vitro and alter Fascin’s localization along actin bundles in Drosophila hemocytes and mammalian cell lines. However, the phospho-mutant can rescue filopodia formation and migration in Drosophila hemocytes and actin bundle formation in Drosophila nurse cells [66]. These data suggest that Fascin requires dynamic cycles of phosphorylation and dephosphorylation to properly form filopodia and promote migration. It remains unknown what kinase mediates phosphorylation at S274 (Drosophila S289) and what phosphatases regulate both phosphorylation sites; however, calcineurin is able to dephosphorylate Fascin in vitro [64]. While the phosphorylation of Fascin has largely been investigated in the context of Fascin’s actin bundling function, phosphorylation also regulates Fascin’s non-canonical roles.
The phosphorylation of Fascin controls its microtubule binding, nuclear localization, and LINC Complex interaction. Specifically, the phosphorylation of S274 promotes Fascin’s interaction with microtubules in cultured cells [17]. In Drosophila, phosphorylation at S289 may promote, while dephosphorylation at S52 (mammalian S39) may inhibit Fascin’s nuclear localization (Groen and Tootle, unpublished observations). Finally, the phosphorylation of S39 (Drosophila S52) promotes Fascin’s perinuclear localization and binding to Nesprin-2, the cytoplasmic portion of the LINC Complex, in both mammalian cells and Drosophila nurse cells [18]. Interestingly, binding to Nesprin 2 disrupts Fascin’s actin-binding site 2 and phosphorylation at S39 impairs actin-binding site 1 (Figure 1B); therefore, how Fascin connects the LINC Complex to the actin cytoskeleton is unclear. Potentially, Fascin is able to bind to an actin filament using part of actin-binding site 1, such as the actin-binding area in the cleft between β-trefoil 1 and 2, while still interacting with the LINC Complex via β-trefoil 3. Together, these data suggest that phosphorylation coordinately regulates multiple functions of Fascin.
In addition to phosphorylation, Fascin has other post-translational modifications. In vitro, Fascin is monoubiquitinated at two different lysine residues, lysine 247 (K247) and lysine 250 (K250). These residues reside in Fascin’s actin-binding site 1 on β-trefoil 2 (Figure 1B) [67]. Ubiquitination at these sites impairs bundle formation and accelerates bundle disassembly [67]. In vitro, this ubiquitination is mediated by the E3 ligase, Smurf1 [67]. However, how ubiquitination regulates both the canonical and non-canonical functions of Fascin is not fully understood.
3.2. Regulation by Prostaglandin Signaling
An upstream regulator of Fascin is prostaglandin (PG) signaling. PGs are short-range lipid signaling molecules that regulate various physiological processes through numerous downstream targets [68][69]. One group of downstream targets of PG signaling is the actin cytoskeleton [70][71][72][73][74][75][76]. Studies using Drosophila oogenesis uncovered that PG signaling regulates numerous actin-binding proteins, including Fascin, Ena, and Myosin II [71][77][78]. In Drosophila, PG signaling positively regulates Fascin to mediate actin bundle formation and cortical actin integrity in the nurse cells [71] and promotes the collective migration of the border cells [79]. In these contexts, PG signaling likely regulates both the actin-bundling activity of Fascin, and its roles in regulating Ena and Myosin II. Indeed, PGs act upstream of Ena to promote actin filament formation [77][78] and control Myosin II-dependent cellular contraction [78]. During Drosophila oogenesis, PG signaling is required for Fascin’s perinuclear localization [19]. As this perinuclear localization also requires a functional LINC Complex [18], PG signaling likely regulates Fascin’s interaction with the LINC Complex. PG signaling also regulates the nuclear localization of Fascin [19]. Specifically, PG signaling temporally regulates the levels of nuclear Fascin, and through Fascin, it regulates nucleolar morphology [19]. These data support the model that PG signaling coordinately regulates the multiple functions of Fascin to mediate developmental processes. While the mechanisms by which PG signaling regulates Fascin remain unknown, PGs do not regulate Fascin expression [71][80]. Furthermore, unpublished observations suggest that PGs modulate Fascin’s post-translational modifications. Notably, the kinase known to phosphorylate Fascin, PKC, and the putative phosphatase, calcineurin, are both downstream targets of PGs [68][81]. Further studies are needed to fully elucidate how PG signaling regulates the different functions of Fascin. This regulation of Fascin is likely important in other contexts, such as cancer metastasis.
3.3. Protein–Protein Interactions
Protein–protein interactions are another means of regulating Fascin. As mentioned above, PKC phosphorylates and binds to Fascin, which inhibits Fascin’s actin bundling function and mediates the localization of the complex to cell margins [65]. Intriguingly, the Fascin–PKC interaction contributes to myoblast migration on fibronectin [65], and both Fascin’s actin bundling and PKC interaction promote migration in human colon carcinoma cells [6]. The Fascin–PKC interaction is positively regulated upstream by the Rho GTPase, Rac, through its effector Pak1 in human colon carcinoma cells [82]. Thus, PKC not only regulates Fascin by phosphorylation but also by directly binding to Fascin.
Filopodia formation and cell migration are also mediated by Fascin’s direct interaction with other partners. For example, Fascin binds to p-Lin-11/Isl-1/Mec-3 kinases (LIMK) to promote filopodia formation downstream of Rho [83]. Fascin also promotes filopodia formation in vitro through its interaction with Daam1, an actin polymerization protein [84]. Additionally, by directly binding to the p75 neurotrophin receptor, Fascin promotes migration of melanoma cells [85]. This interaction is inhibited by the phosphorylation of S39 on Fascin [85]. Fascin also interacts with the vesicle trafficking protein, Rab35 [86]. Rab35 may recruit Fascin to actin bundling sites, as disruption of the interaction impairs filopodia formation in cultured cells [86]. These data indicate numerous proteins bind to and regulate Fascin and suggest that these means of regulation may occur in a cell context-specific manner.
3.4. Transcriptional Regulation
In many instances, Fascin expression is upregulated to induce or promote the migration of cells. For example, Fascin expression is upregulated during dendritic cell maturation through a core promoter region containing a cyclic adenosine monophosphate (cAMP) response element, an enhancer, and a distal repressor [87][88]. These elements regulate Fascin expression, which promotes dendritic cell migration and T-cell activation [89]. In Drosophila, Fascin expression is also transcriptionally upregulated in the migrating border cells [22][90]. The border cells are specified through Janus kinase (JAK)-signal transducer and activator of transcription (STAT) signaling [91]. JAK/STAT signaling induces the expression of the CCAAT-enhancer-binding proteins (C/EBP) transcription factor, Slbo, which then promotes the expression of hundreds of migratory genes in the border cells [90][91]. One of Slbo’s targets is Fascin [90]. Fascin is dramatically upregulated in the border cells during their specification, remains highly expressed throughout the migration, and is critical for on-time migration [22][47][90]. These studies demonstrate that transcriptional expression is one of the mechanisms by which Fascin is regulated to control cellular migrations.
The transcriptional regulation of Fascin has also been extensively studied in the context of cancer. Understanding how Fascin, which is not typically expressed in adult epithelial tissues, becomes highly expressed in malignant carcinomas is of particular interest [3][4]. A study of Fascin’s promoter identified region −219/+114 as having strong transcriptional activity [92]. The cAMP response element-binding protein (CREB) and aryl hydrocarbon receptor binding motifs in this region are potent regulators of fascin transcription in colon, breast, and lung carcinomas [92][93][94]. Specifically, mutations in the binding sites significantly decreased Fascin expression in cancer cell lines [92]. There is also evidence that Fascin is a putative target of β-catenin–T-cell factor (TCF) signaling in colon and gastric carcinomas [95][96], Transforming growth factor beta (TGFβ) signaling in breast carcinomas [97][98] and STAT3/ Nuclear factor kappa B (NF-kB) signaling in breast and gastric carcinomas [99][100]. The specific signaling mechanisms that induce high Fascin expression in malignant carcinomas are likely dependent on the cellular context. However, understanding the mechanisms that increase Fascin expression in different cancers is crucial for understanding Fascin’s roles in promoting cancer progression.
Another mechanism controlling the expression of Fascin is microRNAs (miRs). miRs are small, non-coding RNA molecules that post-transcriptionally regulate gene expression. Two microRNAs, miR133 and miR145, inhibit Fascin expression, and they are downregulated in many types of cancer, including breast, bladder, esophageal squamous cell, prostate, and colorectal [101][102][103][104][105][106][107][108][109][110]. In these cancers, the downregulation of these microRNAs leads to increased Fascin expression, which is linked to increased cell migration and invasion [101][102][103][104][105][106][107][108][109][110]. Thus, post-transcriptional regulation is a critical mechanism of controlling the expression of Fascin.
These studies indicate that the functions and expression of Fascin are regulated through various mechanisms, including post-translation modifications, upstream regulation, and expression. Given its canonical and non-canonical functions, Fascin likely requires many layers of regulation to ensure its proper function. Further investigation into how Fascin’s many functions are regulated and coordinated in a cell is warranted.
References
- De Arcangelis, A.; Georges-Labouesse, E.; Adams, J.C. Expression of fascin-1, the gene encoding the actin-bundling protein fascin-1, during mouse embryogenesis. Gene Expr. Patterns 2004, 4, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Machesky, L.M. Fascin1 in carcinomas: Its regulation and prognostic value. Int. J. Cancer 2015, 137, 2534–2544. [Google Scholar] [CrossRef] [PubMed]
- Jayo, A.; Parsons, M. Fascin: A key regulator of cytoskeletal dynamics. Int. J. Biochem. Cell Biol. 2010, 42, 1614–1617. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Kim, D.J.; Adams, J.C. The roles of fascins in health and disease. J. Pathol. 2011, 224, 289–300. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Parsons, M.; Adams, J.C. Dual Actin-bundling and Protein Kinase C-binding Activities of Fascin Regulate Carcinoma Cell Migration Downstream of Rac and Contribute to Metastasis. Mol. Biol. Cell 2007, 18, 4591–4602. [Google Scholar] [CrossRef]
- Van Audenhove, I.; Denert, M.; Boucherie, C.; Pieters, L.; Cornelissen, M.; Gettemans, J. Fascin Rigidity and L-plastin Flexibility Cooperate in Cancer Cell Invadopodia and Filopodia. J. Biol. Chem. 2016, 291, 9148–9160. [Google Scholar] [CrossRef]
- Vignjevic, D.M.; Kojima, S.-I.; Aratyn, Y.; Danciu, O.; Svitkina, T.; Borisy, G.G. Role of fascin in filopodial protrusion. J. Cell Biol. 2006, 174, 863–875. [Google Scholar] [CrossRef]
- Elkhatib, N.; Neu, M.B.; Zensen, C.; Schmoller, K.M.; Louvard, D.; Bausch, A.R.; Betz, T.; Vignjevic, D.M. Fascin Plays a Role in Stress Fiber Organization and Focal Adhesion Disassembly. Curr. Biol. 2014, 24, 1492–1499. [Google Scholar] [CrossRef]
- Adams, J.C. Characterization of Cell–Matrix Adhesion Requirements for the Formation of Fascin Microspikes. Mol. Biol. Cell 1997, 8, 2345–2363. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.C.; Clelland, J.D.; Collett, G.D.; Matsumura, F.; Yamashiro, S.; Zhang, L. Cell-Matrix Adhesions Differentially Regulate Fascin Phosphorylation. Mol. Biol. Cell 1999, 10, 4177–4190. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Dawson, J.C.; Forerovargas, M.G.; Spence, H.J.; Yu, X.; König, I.; Anderson, K.I.; Machesky, L.M. The Actin-Bundling Protein Fascin Stabilizes Actin in Invadopodia and Potentiates Protrusive Invasion. Curr. Biol. 2010, 20, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Kliewe, F.; Scharf, C.; Rogge, H.; Darm, K.; Lindenmeyer, M.T.; Amann, K.; Cohen, C.D.; Endlich, K.; Endlich, N. Studying the role of fascin-1 in mechanically stressed podocytes. Sci. Rep. 2017, 7, 9916. [Google Scholar] [CrossRef] [PubMed]
- Quintavalle, M.; Elia, L.; Condorelli, G.; Courtneidge, S.A. MicroRNA control of podosome formation in vascular smooth muscle cells in vivo and in vitro. J. Cell Biol. 2010, 189, 13–22. [Google Scholar] [CrossRef]
- Zhang, F.-R.; Tao, L.-H.; Shen, Z.-Y.; Lv, Z.; Xu, L.; Li, E.-M. Fascin Expression in Human Embryonic, Fetal, and Normal Adult Tissue. J. Histochem. Cytochem. 2008, 56, 193–199. [Google Scholar] [CrossRef]
- Harker, A.J.; Katkar, H.H.; Bidone, T.C.; Aydin, F.; Voth, G.A.; Applewhite, D.A.; Kovar, D.R. Ena/VASP processive elongation is modulated by avidity on actin filaments bundled by the filopodia cross-linker fascin. Mol. Biol. Cell 2019, 30, 851–862. [Google Scholar] [CrossRef]
- Winkelman, J.D.; Bilancia, C.G.; Peifer, M.; Kovar, D.R. Ena/VASP Enabled is a highly processive actin polymerase tailored to self-assemble parallel-bundled F-actin networks with Fascin. Proc. Natl. Acad. Sci. USA 2014, 111, 4121–4126. [Google Scholar] [CrossRef] [PubMed]
- Villari, G.; Jayo, A.; Zanet, J.; Fitch, B.; Serrels, B.; Frame, M.C.; Stramer, B.M.; Goult, B.T.; Parsons, M. A direct interaction between fascin and microtubules contributes to adhesion dynamics and cell migration. J. Cell Sci. 2015, 128, 4601–4614. [Google Scholar] [CrossRef] [PubMed]
- Jayo, A.; Malboubi, M.; Antoku, S.; Chang, W.; Ortiz-Zapater, E.; Groen, C.; Pfisterer, K.; Tootle, T.; Charras, G.; Gundersen, G.G.; et al. Fascin Regulates Nuclear Movement and Deformation in Migrating Cells. Dev. Cell 2016, 38, 371–383. [Google Scholar] [CrossRef] [PubMed]
- Groen, C.M.; Jayo, A.; Parsons, M.; Tootle, T.L. Prostaglandins regulate nuclear localization of Fascin and its function in nucleolar architecture. Mol. Biol. Cell 2015, 26, 1901–1917. [Google Scholar] [CrossRef]
- Kane, R.E. Preparation and purification of polymerized actin from sea urchin egg extracts. J. Cell Biol. 1975, 66, 305–315. [Google Scholar] [CrossRef]
- Cant, K.; Cooley, L. Single Amino Acid Mutations in Drosophila Fascin Disrupt Actin Bundling Function in Vivo. Genetics 1996, 143, 249–258. [Google Scholar] [PubMed]
- Cant, K.; Knowles, B.A.; Mooseker, M.S.; Cooley, L. Drosophila singed, a fascin homolog, is required for actin bundle formation during oogenesis and bristle extension. J. Cell Biol. 1994, 125, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Holthuis, J.C.; Schoonderwoert, V.T.; Martens, G.J. A vertebrate homolog of the actin-bundling protein fascin. Biochim. Biophys. Acta 1994, 1219, 184–188. [Google Scholar] [CrossRef]
- Edwards, R.A.; Herrera-Sosa, H.; Otto, J.; Bryan, J. Cloning and Expression of a Murine Fascin Homolog from Mouse Brain. J. Biol. Chem. 1995, 270, 10764–10770. [Google Scholar] [CrossRef]
- Duh, F.-M.; Latif, F.; Weng, Y.; Geil, L.; Modi, W.; Stackhouse, T.; Matsumura, F.; Duan, D.R.; Linehan, W.M.; Lerman, M.I.; et al. cDNA Cloning and Expression of the Human Homolog of the Sea Urchin fascin and Drosophila singed Genes Which Encodes an Actin-Bundling Protein. DNA Cell Biol. 1994, 13, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Yamashiro-Matsumura, S.; Matsumura, F. Purification and characterization of an F-actin-bundling 55-kilodalton protein from HeLa cells. J. Biol. Chem. 1985, 260, 5087–5097. [Google Scholar]
- Murzin, A.G.; Lesk, A.M.; Chothia, C. Beta-Trefoil fold. Patterns of structure and sequence in the Kunitz inhibitors interleukins-1 beta and 1 alpha and fibroblast growth factors. J. Mol. Biol. 1992, 223, 531–543. [Google Scholar] [CrossRef]
- Ponting, C.P.; Russell, R.B. Identification of distant homologues of fibroblast growth factors suggests a common ancestor for all beta-trefoil proteins. J. Mol. Biol. 2000, 302, 1041–1047. [Google Scholar] [CrossRef]
- Yang, S.; Huang, F.-K.; Huang, J.; Chen, S.; Jakoncic, J.; Leo-Macias, A.; Diaz-Avalos, R.; Chen, L.; Zhang, J.J.; Huang, X.-Y. Molecular Mechanism of Fascin Function in Filopodial Formation. J. Biol. Chem. 2013, 288, 274–284. [Google Scholar] [CrossRef]
- Aramaki, S.; Mayanagi, K.; Jin, M.; Aoyama, K.; Yasunaga, T. Filopodia formation by crosslinking of F-actin with fascin in two different binding manners. Cytoskeleton (Hoboken) 2016, 73, 365–374. [Google Scholar] [CrossRef]
- Jansen, S.; Collins, A.; Yang, C.; Rebowski, G.; Svitkina, T.; Dominguez, R. Mechanism of Actin Filament Bundling by Fascin. J. Biol. Chem. 2011, 286, 30087–30096. [Google Scholar] [CrossRef] [PubMed]
- Lin-Jones, J.; Burnside, B. Retina-Specific Protein Fascin 2 Is an Actin Cross-linker Associated with Actin Bundles in Photoreceptor Inner Segments and Calycal Processes. Investig. Opthalmol. Vis. Sci. 2007, 48, 1380–1388. [Google Scholar] [CrossRef] [PubMed]
- Perrin, B.J.; Strandjord, D.M.; Narayanan, P.; Henderson, D.M.; Johnson, K.R.; Ervasti, J.M. Beta-Actin and fascin-2 cooperate to maintain stereocilia length. J. Neurosci. 2013, 33, 8114–8121. [Google Scholar] [CrossRef] [PubMed]
- Tubb, B.E.; Bardien-Kruger, S.; Kashork, C.D.; Shaffer, L.G.; Ramagli, L.S.; Xu, J.; Siciliano, M.J.; Bryan, J. Characterization of Human Retinal Fascin Gene (FSCN2) at 17q25: Close Physical Linkage of Fascin and Cytoplasmic Actin Genes. Genomics 2000, 65, 146–156. [Google Scholar] [CrossRef]
- Gamundi, M.J.; Hernan, I.; Maseras, M.; Baiget, M.; Ayuso, C.; Borrego, S.; Antiñolo, G.; Millán, J.M.; Valverde, D.; Carballo, M. Sequence variations in the retinal fascin FSCN2 gene in a Spanish population with autosomal dominant retinitis pigmentosa or macular degeneration. Mol. Vis. 2005, 11, 922–928. [Google Scholar]
- Yokokura, S.; Wada, Y.; Nakai, S.; Sato, H.; Yao, R.; Yamanaka, H.; Ito, S.; Sagara, Y.; Takahashi, M.; Nakamura, Y.; et al. Targeted Disruption of FSCN2 Gene Induces Retinopathy in Mice. Investig. Opthalmol. Vis. Sci. 2005, 46, 2905–2915. [Google Scholar] [CrossRef]
- Tubb, B.; Mulholland, D.J.; Vogl, W.; Lan, Z.-J.; Niederberger, C.; Cooney, A.; Bryan, J. Testis Fascin (FSCN3): A Novel Paralog of the Actin-Bundling Protein Fascin Expressed Specifically in the Elongate Spermatid Head. Exp. Cell Res. 2002, 275, 92–109. [Google Scholar] [CrossRef]
- Yamakita, Y.; Matsumura, F.; Yamashiro, S. Fascin1 is dispensable for mouse development but is favorable for neonatal survival. Cell Motil. Cytoskelet. 2009, 66, 524–534. [Google Scholar] [CrossRef]
- Yamakita, Y.; Matsumura, F.; Lipscomb, M.W.; Chou, P.-C.; Werlen, G.; Burkhardt, J.K.; Yamashiro, S. Fascin1 Promotes Cell Migration of Mature Dendritic Cells. J. Immunol. 2011, 186, 2850–2859. [Google Scholar] [CrossRef]
- Boer, E.; Howell, E.D.; Schilling, T.F.; Jette, C.A.; Stewart, R.A. Fascin1-Dependent Filopodia are Required for Directional Migration of a Subset of Neural Crest Cells. PLoS Genet. 2015, 11, e1004946. [Google Scholar] [CrossRef]
- Ma, Y.; Li, A.; Faller, W.J.; Libertini, S.; Fiorito, F.; Gillespie, D.A.; Sansom, O.J.; Yamashiro, S.; Machesky, L.M. Fascin 1 is transiently expressed in mouse melanoblasts during development and promotes migration and proliferation. Development 2013, 140, 2203–2211. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Reynolds, L.E.; Li, A.; Stevenson, R.P.; Hodivala-Dilke, K.M.; Yamashiro, S.; Machesky, L.M. Fascin 1 is dispensable for developmental and tumour angiogenesis. Biol. Open 2013, 2, 1187–1191. [Google Scholar] [CrossRef]
- Mosialos, G.; Birkenbach, M.; Ayehunie, S.; Matsumura, F.; Pinkus, G.S.; Kieff, E.; Langhoff, E. Circulating human dendritic cells differentially express high levels of a 55-kd actin-bundling protein. Am. J. Pathol. 1996, 148, 593–600. [Google Scholar] [PubMed]
- Sonego, M.; Gajendra, S.; Parsons, M.; Ma, Y.; Hobbs, C.; Zentar, M.P.; Williams, G.; Machesky, L.M.; Doherty, P.; Lalli, G. Fascin regulates the migration of subventricular zone-derived neuroblasts in the postnatal brain. J. Neurosci. 2013, 33, 12171–12185. [Google Scholar] [CrossRef] [PubMed]
- DeRosier, D.J.; Edds, K.T. Evidence for fascin cross-links between the actin filaments in coelomocyte filopodia. Exp. Cell Res. 1980, 126, 490–494. [Google Scholar] [CrossRef]
- Zanet, J.; Stramer, B.M.; Millard, T.; Martin, P.; Payre, F.; Plaza, S. Fascin is required for blood cell migration during Drosophila embryogenesis. Development 2009, 136, 2557–2565. [Google Scholar] [CrossRef] [PubMed]
- Lamb, M.C.; Anliker, K.K.; Tootle, T.L. Fascin regulates protrusions and delamination to mediate invasive, collective cell migration in vivo. Dev. Dyn. 2020, 249, 961–982. [Google Scholar] [CrossRef] [PubMed]
- Bear, J.E.; Gertler, F.B. Ena/VASP: Towards resolving a pointed controversy at the barbed end. J. Cell Sci. 2009, 122, 1947–1953. [Google Scholar] [CrossRef]
- Vicente-Manzanares, M.; Ma, X.; Adelstein, R.S.; Horwitz, A.R. Non-muscle myosin II takes centre stage in cell adhesion and migration. Nat. Rev. Mol. Cell Biol. 2009, 10, 778–790. [Google Scholar] [CrossRef]
- Lin, S.; Huang, C.; Gunda, V.; Sun, J.; Chellappan, S.P.; Li, Z.; Izumi, V.; Fang, B.; Koomen, J.; Singh, P.K.; et al. Fascin Controls Metastatic Colonization and Mitochondrial Oxidative Phosphorylation by Remodeling Mitochondrial Actin Filaments. Cell Rep. 2019, 28, 2824–2836.e8. [Google Scholar] [CrossRef]
- Lombardi, M.L.; Jaalouk, D.E.; Shanahan, C.M.; Burke, B.; Roux, K.J.; Lammerding, J. The Interaction between Nesprins and Sun Proteins at the Nuclear Envelope Is Critical for Force Transmission between the Nucleus and Cytoskeleton. J. Biol. Chem. 2011, 286, 26743–26753. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, M.L.; Lammerding, J. Keeping the LINC: The importance of nucleocytoskeletal coupling in intracellular force transmission and cellular function. Biochem. Soc. Trans. 2011, 39, 1729–1734. [Google Scholar] [CrossRef] [PubMed]
- Alam, H.; Bhate, A.V.; Gangadaran, P.; Sawant, S.; Salot, S.; Sehgal, L.; Dange, P.P.; Chaukar, D.A.; D’Cruz, A.; Kannan, S.; et al. Fascin overexpression promotes neoplastic progression in oral squamous cell carcinoma. BMC Cancer 2012, 12, 32. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Swift, J.; Irianto, J.; Shin, J.-W.; Spinler, K.R.; Athirasala, A.; Diegmiller, R.; Dingal, P.D.P.; Ivanovska, I.L.; Discher, D.E. Nuclear lamin stiffness is a barrier to 3D migration, but softness can limit survival. J. Cell Biol. 2014, 204, 669–682. [Google Scholar] [CrossRef]
- Kelpsch, D.J.; Groen, C.M.; Fagan, T.N.; Sudhir, S.; Tootle, T.L. Fascin regulates nuclear actin during Drosophila oogenesis. Mol. Biol. Cell 2016, 27, 2965–2979. [Google Scholar] [CrossRef]
- Kelpsch, D.J.; Tootle, T.L. Nuclear Actin: From Discovery to Function. Anat. Rec. (Hoboken) 2018, 301, 1999–2013. [Google Scholar] [CrossRef]
- Vartiainen, M.K.; Guettler, S.; Larijani, B.; Treisman, R. Nuclear Actin Regulates Dynamic Subcellular Localization and Activity of the SRF Cofactor MAL. Science 2007, 316, 1749–1752. [Google Scholar] [CrossRef]
- Gau, D.; Roy, P. SRF’ing and SAP’ing—The role of MRTF proteins in cell migration. J. Cell Sci. 2018, 131. [Google Scholar] [CrossRef]
- Feric, M.; Vaidya, N.; Harmon, T.S.; Mitrea, D.M.; Zhu, L.; Richardson, T.M.; Kriwacki, R.W.; Pappu, R.V.; Brangwynne, C.P. Coexisting Liquid Phases Underlie Nucleolar Subcompartments. Cell 2016, 165, 1686–1697. [Google Scholar] [CrossRef]
- Percipalle, P. Co-transcriptional nuclear actin dynamics. Nucleus 2013, 4, 43–52. [Google Scholar] [CrossRef]
- Liu, Q.; Cui, J.; Zhang, Y.; Niu, M.; Xue, X.; Yin, H.; Tang, Y.; Dai, L.; Dai, F.; Guo, Y.; et al. Mass spectrometry-based proteomic analysis of FSCN1-interacting proteins in laryngeal squamous cell carcinoma cells. IUBMB Life 2019, 71, 1771–1784. [Google Scholar] [CrossRef] [PubMed]
- Saad, A.; Bijian, K.; Qiu, D.; Da Silva, S.D.; Marques, M.; Chang, C.-H.; Nassour, H.; Ramotar, D.; Damaraju, S.; Mackey, J.; et al. Insights into a novel nuclear function for Fascin in the regulation of the amino-acid transporter SLC3A2. Sci. Rep. 2016, 6, 36699. [Google Scholar] [CrossRef] [PubMed]
- Ono, S.; Yamakita, Y.; Yamashiro, S.; Matsudaira, P.T.; Gnarra, J.R.; Obinata, T.; Matsumura, F. Identification of an Actin Binding Region and a Protein Kinase C Phosphorylation Site on Human Fascin. J. Biol. Chem. 1997, 272, 2527–2533. [Google Scholar] [CrossRef] [PubMed]
- Yamakita, Y.; Ono, S.; Matsumura, F.; Yamashiro, S. Phosphorylation of Human Fascin Inhibits Its Actin Binding and Bundling Activities. J. Biol. Chem. 1996, 271, 12632–12638. [Google Scholar] [CrossRef] [PubMed]
- Anilkumar, N.; Parsons, M.; Monk, R.; Ng, T.; Adams, J.C. Interaction of fascin and protein kinase Calpha: A novel intersection in cell adhesion and motility. EMBO J. 2003, 22, 5390–5402. [Google Scholar] [CrossRef]
- Zanet, J.; Jayo, A.; Plaza, S.; Millard, T.; Parsons, M.; Stramer, B. Fascin promotes filopodia formation independent of its role in actin bundling. J. Cell Biol. 2012, 197, 477–486. [Google Scholar] [CrossRef]
- Lin, S.; Lu, S.; Mulaj, M.; Fang, B.; Keeley, T.; Wan, L.; Hao, J.; Muschol, M.; Sun, J.; Yang, S. Monoubiquitination Inhibits the Actin Bundling Activity of Fascin. J. Biol. Chem. 2016, 291, 27323–27333. [Google Scholar] [CrossRef]
- Bos, C.L.; Richel, D.J.; Ritsema, T.; Peppelenbosch, M.P.; Versteeg, H.H. Prostanoids and prostanoid receptors in signal transduction. Int. J. Biochem. Cell Biol. 2004, 36, 1187–1205. [Google Scholar] [CrossRef]
- Tootle, T.L. Genetic insights into the in vivo functions of prostaglandin signaling. Int. J. Biochem. Cell Biol. 2013, 45, 1629–1632. [Google Scholar] [CrossRef]
- Peppelenbosch, M.P.; Tertoolen, L.G.; Hage, W.J.; De Laat, S.W. Epidermal growth factor—Induced actin remodeling is regulated by 5-lipoxygenase and cyclooxygenase products. Cell 1993, 74, 565–575. [Google Scholar] [CrossRef]
- Groen, C.M.; Spracklen, A.J.; Fagan, T.N.; Tootle, T.L. DrosophilaFascin is a novel downstream target of prostaglandin signaling during actin remodeling. Mol. Biol. Cell 2012, 23, 4567–4578. [Google Scholar] [CrossRef] [PubMed]
- Tootle, T.L.; Spradling, A.C. Drosophila Pxt: A cyclooxygenase-like facilitator of follicle maturation. Development 2008, 135, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Pierce, K.L.; Fujino, H.; Srinivasan, D.; Regan, J.W. Activation of FP Prostanoid Receptor Isoforms Leads to Rho-mediated Changes in Cell Morphology and in the Cell Cytoskeleton. J. Biol. Chem. 1999, 274, 35944–35949. [Google Scholar] [CrossRef]
- Bulin, C.; Albrecht, U.; Bode, J.; Weber, A.-A.; Schrör, K.; Levkau, B.; Fischer, J.W. Differential Effects of Vasodilatory Prostaglandins on Focal Adhesions, Cytoskeletal Architecture, and Migration in Human Aortic Smooth Muscle Cells. Arter. Thromb. Vasc. Biol. 2005, 25, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Dormond, O.; Bezzi, M.; Mariotti, A.; Rüegg, C. Prostaglandin E2 promotes integrin alpha Vbeta 3-dependent endothelial cell adhesion, rac-activation, and spreading through cAMP/PKA-dependent signaling. J. Biol. Chem. 2002, 277, 45838–45846. [Google Scholar] [CrossRef]
- Birukova, A.A.; Zagranichnaya, T.; Fu, P.; Alekseeva, E.; Chen, W.; Jacobson, J.R.; Birukov, K.G. Prostaglandins PGE(2) and PGI(2) promote endothelial barrier enhancement via PKA—And Epac1/Rap1-dependent Rac activation. Exp. Cell Res. 2007, 313, 2504–2520. [Google Scholar] [CrossRef]
- Spracklen, A.J.; Kelpsch, D.J.; Chen, X.; Spracklen, C.N.; Tootle, T.L. Prostaglandins temporally regulate cytoplasmic actin bundle formation during Drosophila oogenesis. Mol. Biol. Cell 2014, 25, 397–411. [Google Scholar] [CrossRef]
- Spracklen, A.J.; Lamb, M.C.; Groen, C.M.; Tootle, T.L. Pharmaco-Genetic Screen to Uncover Actin Regulators Targeted by Prostaglandins During Drosophila Oogenesis. G3 (Bethesda) 2019, 9, 3555–3565. [Google Scholar] [CrossRef]
- Tootle, T.L.; Williams, D.; Hubb, A.; Frederick, R.; Spradling, A. Drosophila Eggshell Production: Identification of New Genes and Coordination by Pxt. PLoS ONE 2011, 6, e19943. [Google Scholar] [CrossRef]
- Liu, L.; Clipstone, N.A. Prostaglandin F2alpha inhibits adipocyte differentiation via a G alpha q-calcium-calcineurin-dependent signaling pathway. J. Cell Biochem. 2007, 100, 161–173. [Google Scholar] [CrossRef]
- Parsons, M.; Adams, J.C. Rac regulates the interaction of fascin with protein kinase C in cell migration. J. Cell Sci. 2008, 121, 2805–2813. [Google Scholar] [CrossRef]
- Jayo, A.; Parsons, M.; Adams, J.C. A novel Rho-dependent pathway that drives interaction of fascin-1 with p-Lin-11/Isl-1/Mec-3 kinase (LIMK) 1/2 to promote fascin-1/actin binding and filopodia stability. BMC Biol. 2012, 10, 72. [Google Scholar] [CrossRef]
- Jaiswal, R.; Breitsprecher, D.; Collins, A.; Corrêa, I.R.; Xu, M.-Q.; Goode, B.L. The Formin Daam1 and Fascin Directly Collaborate to Promote Filopodia Formation. Curr. Biol. 2013, 23, 1373–1379. [Google Scholar] [CrossRef] [PubMed]
- Shonukan, T.; Bagayogo, I.; McCrea, P.; Chao, M.V.; Hempstead, B.; Shonukan, O.; McCrea, P. Neurotrophin-induced melanoma cell migration is mediated through the actin-bundling protein fascin. Oncogene 2003, 22, 3616–3623. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Fonovic, M.; Suyama, K.; Bogyo, M.; Scott, M.P. Rab35 Controls Actin Bundling by Recruiting Fascin as an Effector Protein. Science 2009, 325, 1250–1254. [Google Scholar] [CrossRef] [PubMed]
- Bros, M.; Ross, X.-L.; Pautz, A.; Reske-Kunz, A.B.; Ross, R. The Human Fascin Gene Promoter Is Highly Active in Mature Dendritic Cells Due to a Stage-Specific Enhancer. J. Immunol. 2003, 171, 1825–1834. [Google Scholar] [CrossRef] [PubMed]
- Ross, R.; Jonuleit, H.; Bros, M.; Ross, X.-L.; Enk, A.H.; Knop, J.; Reske-Kunz, A.B.; Yamashiro, S.; Matsumura, F. Expression of the Actin-Bundling Protein Fascin in Cultured Human Dendritic Cells Correlates with Dendritic Morphology and Cell Differentiation. J. Investig. Dermatol. 2000, 115, 658–663. [Google Scholar] [CrossRef]
- Geyeregger, R.; Zeyda, M.; Bauer, W.M.; Kriehuber, E.; Säemann, M.D.; Zlabinger, G.J.; Maurer, D.; Stulnig, T. Liver X receptors regulate dendritic cell phenotype and function through blocked induction of the actin-bundling protein fascin. Blood 2007, 109, 4288–4295. [Google Scholar] [CrossRef]
- Borghese, L.; Fletcher, G.; Mathieu, J.; Atzberger, A.; Eades, W.C.; Cagan, R.L.; Rørth, P. Systematic Analysis of the Transcriptional Switch Inducing Migration of Border Cells. Dev. Cell 2006, 10, 497–508. [Google Scholar] [CrossRef]
- Silver, D.L.; Geisbrecht, E.R.; Montell, D.J. Requirement for JAK/STAT signaling throughout border cell migration in Drosophila. Development 2005, 132, 3483–3492. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Loftis, D.W.; Adams, J.C. Fascin-1 Promoter Activity Is Regulated by CREB and the Aryl Hydrocarbon Receptor in Human Carcinoma Cells. PLoS ONE 2009, 4, e5130. [Google Scholar] [CrossRef] [PubMed]
- Grothey, A.; Hashizume, R.; Ji, H.; Tubb, B.E.; Patrick, C.W.; Yu, D.; Money, E.E.; McCrea, P.D. C-erbB-2/ HER-2 upregulates fascin, an actin-bundling protein associated with cell motility, in human breast cancer cell lines. Oncogene 2000, 19, 4864–4875. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Jin, L.; Alesi, G.N.; Kim, Y.-M.; Fan, J.; Seo, J.H.; Wang, N.; Tucker, M.; Gu, T.-L.; Lee, B.H.; et al. The Prometastatic Ribosomal S6 Kinase 2-cAMP Response Element-binding Protein (RSK2-CREB) Signaling Pathway Up-regulates the Actin-binding Protein Fascin-1 to Promote Tumor Metastasis. J. Biol. Chem. 2013, 288, 32528–32538. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Choi, I.; Cheong, T.; Lee, S.; Lotan, R.; Park, S.H.; Chun, K.-H. Galectin-3 Increases Gastric Cancer Cell Motility by Up-regulating Fascin-1 Expression. Gastroenterology 2010, 138, 1035–1045.e2. [Google Scholar] [CrossRef] [PubMed]
- Vignjevic, D.M.; Schoumacher, M.; Gavert, N.; Janssen, K.-P.; Jih, G.; Laé, M.; Louvard, D.; Ben-Ze’Ev, A.; Robine, S. Fascin, a Novel Target of beta--Catenin-TCF Signaling, Is Expressed at the Invasive Front of Human Colon Cancer. Cancer Res. 2007, 67, 6844–6853. [Google Scholar] [CrossRef]
- Sun, J.; He, H.; Pillai, S.; Xiong, Y.; Challa, S.; Xu, L.; Chellappan, S.; Yang, S. GATA3 Transcription Factor Abrogates Smad4 Transcription Factor-mediated Fascin Overexpression, Invadopodium Formation, and Breast Cancer Cell Invasion. J. Biol. Chem. 2013, 288, 36971–36982. [Google Scholar] [CrossRef]
- Sun, J.; He, H.; Xiong, Y.; Lu, S.; Shen, J.; Cheng, A.; Chang, W.-C.; Hou, M.-F.; Lancaster, J.M.; Kim, M.; et al. Fascin protein is critical for transforming growth factor beta protein-induced invasion and filopodia formation in spindle-shaped tumor cells. J. Biol. Chem. 2011, 286, 38865–38875. [Google Scholar] [CrossRef]
- Snyder, M.; Huang, X.-Y.; Zhang, J.J. Signal Transducers and Activators of Transcription 3 (STAT3) Directly Regulates Cytokine-induced Fascin Expression and Is Required for Breast Cancer Cell Migration. J. Biol. Chem. 2011, 286, 38886–38893. [Google Scholar] [CrossRef]
- Yao, J.; Qian, C.J.; Ye, B.; Zhao, Z.Q.; Wei, J.; Liang, Y.; Zhang, X. Signal. transducer and activator of transcription 3 signaling upregulates fascin via nuclear factor-kappaB in gastric cancer: Implications in cell invasion and migration. Oncol. Lett. 2014, 7, 902–908. [Google Scholar] [CrossRef]
- Chivomaru, T.; Enokida, H.; Tatarano, S.; Kawahara, K.; Nishiyama, K.; Seki, N.; Nakagawa, M. miR-145 and miR-133a function as tumour suppressors and directly regulate FSCN1 expression in bladder cancer. Br. J. Cancer 2010, 102, 883–891. [Google Scholar] [CrossRef]
- Feng, Y.; Zhu, J.; Ou, C.; Deng, Z.; Chen, M.; Huang, W.; Liu, L.-Z. MicroRNA-145 inhibits tumour growth and metastasis in colorectal cancer by targeting fascin-1. Br. J. Cancer 2014, 110, 2300–2309. [Google Scholar] [CrossRef] [PubMed]
- Fuse, M.; Seki, N.; Nohata, N.; Kojima, S.; Sakamoto, S.; Chiyomaru, T.; Kawakami, K.; Enokida, H.; Nakagawa, M.; Naya, Y.; et al. Restoration of miR-145 expression suppresses cell proliferation, migration and invasion in prostate cancer by targeting FSCN1. Int. J. Oncol. 2011, 38, 1093–1101. [Google Scholar] [PubMed]
- Gao, R.; Zhang, N.; Yang, J.; Zhu, Y.; Zhang, Z.; Wang, J.; Xu, X.; Li, Z.; Liu, X.; Li, Z.; et al. Long non-coding RNA ZEB1-AS1 regulates miR-200b/FSCN1 signaling and enhances migration and invasion induced by TGF-beta1 in bladder cancer cells. J. Exp. Clin. Cancer Res. 2019, 38, 111. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Zhang, C.; Li, W.; Li, H.; Sang, J.; Zhao, Q.; Bo, Y.; Luo, H.; Zheng, X.; Lu, Y.; et al. Promoter Methylation-Regulated miR-145-5p Inhibits Laryngeal Squamous Cell Carcinoma Progression by Targeting FSCN1. Mol. Ther. 2019, 27, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Götte, M.; Mohr, C.; Koo, C.Y.; Stock, C.; Vaske, A.-K.; Viola, M.; Ibrahim, S.A.; Peddibhotla, S.; Teng, Y.H.-F.; Low, J.-Y.; et al. miR-145-dependent targeting of Junctional Adhesion Molecule A and modulation of fascin expression are associated with reduced breast cancer cell motility and invasiveness. Oncogene 2010, 29, 6569–6580. [Google Scholar] [CrossRef]
- Kano, M.; Seki, N.; Kikkawa, N.; Fujimura, L.; Hoshino, I.; Akutsu, Y.; Chiyomaru, T.; Enokida, H.; Nakagawa, M.; Matsubara, H. miR-145, miR-133a and miR-133b: Tumor-suppressive miRNAs target FSCN1 in esophageal squamous cell carcinoma. Int. J. Cancer 2010, 127, 2804–2814. [Google Scholar] [CrossRef]
- Liu, R.; Liao, J.; Yang, M.; Sheng, J.; Yang, H.; Wang, Y.; Pan, E.; Guo, W.; Pu, Y.; Kim, S.J.; et al. The Cluster of miR-143 and miR-145 Affects the Risk for Esophageal Squamous Cell Carcinoma through Co-Regulating Fascin Homolog 1. PLoS ONE 2012, 7, e33987. [Google Scholar] [CrossRef]
- Wu, Z.; Wang, C.-Q.; Xiang, R.; Liu, X.; Ye, S.; Yang, X.-Q.; Zhang, G.-H.; Xu, X.; Zhu, T.; Wu, Q. Loss of miR-133a expression associated with poor survival of breast cancer and restoration of miR-133a expression inhibited breast cancer cell growth and invasion. BMC Cancer 2012, 12, 51. [Google Scholar] [CrossRef]
- Zhao, H.; Kang, X.; Xia, X.; Wo, L.; Gu, X.; Hu, Y.; Xie, X.; Chang, H.; Lou, L.; Shen, X. miR-145 suppresses breast cancer cell migration by targeting FSCN-1 and inhibiting epithelial-mesenchymal transition. Am. J. Transl. Res. 2016, 8, 3106–3114. [Google Scholar]
- Arlt, M.J.E.; Kuzmanov, A.; Snedeker, J.G.; Fuchs, B.; Silvan, U.; Sabile, A.A. Fascin-1 enhances experimental osteosarcoma tumor formation and metastasis and is related to poor patient outcome. BMC Cancer 2019, 19, 83. [Google Scholar] [CrossRef]

