1. Introduction
The genus
Yersinia includes several pathogenic bacterial species for humans, which include the food-borne enteropathogens
Y. enterocolitica and
Y. pseudotuberculosis [
1], and the vector-borne pathogen
Y. pestis, the etiologic agent of plague [
2]. During
Y. enterocolitica and
Y. pseudotuberculosis host infection, interaction between the bacterial surface molecule invasin and host β1-integrins promotes bacterial internalization into intestinal tract cells and colonization of Peyer’s patches and the cecum [
3,
4,
5]. Traversal of the intestinal barrier leads to bacterial draining by local mesenteric lymph nodes, where bacterial extracellular proliferation takes place [
6]. Invasin is inactivated in
Y. pestis [
7] and inoculation through flea bites favors pathogen draining by inguinal, axillary or cervical lymph nodes, where bacterial proliferation takes also place extracellularly [
8]. Inhibition of phagocytosis and bacterial extracellular life are promoted by
Yersinia outer proteins (Yops), translocated into host cells by a type 3 secretion system (T3SS) encoded in the
Yersinia virulence plasmid (pYV in
Y. enterocolitica and
Y. pseudotuberculosis, pCD1 in
Y. pestis) [
9,
10].
The early signaling cascades that trigger
Y. enterocolitica and
Y. pseudotuberculosis entry within epithelial cells upon invasin/β1-integrins interaction have been well studied [
11,
12,
13]. The extracellular stages that characterize lymph node infection by all pathogenic
Yersinia have been also well described in the literature [
14]. However, the bacterial intracellular stages have been less investigated, despite their importance for early infection or bacterial persistence in the intestinal barrier during enteric yersiniosis [
3,
4,
5]. Replication within the intracellular environment of phagocytic cells seems also to play a major role for pathogenic
Yersinia virulence in vivo [
15,
16]. Nevertheless, the precise nature of the intracellular compartments supporting bacterial replication remained unknown.
2. Autophagy
Autophagy is an essential cellular homeostatic process highly conserved in eukaryotes. It directs cytoplasmic cellular components (damaged organelles, misfolded or aggregated proteins) to endo-lysosomal compartments for degradation and recycling. The pan-eukaryotic distribution of core components of the autophagic machinery argues for the presence of autophagy in the last common eukaryotic ancestor [18]. Autophagy probably played a major role during early eukaryotic evolution by allowing cells to survive under starvation conditions and to localize new foraging locations while consuming intracellular pools of amino acids. Currently, autophagy plays major roles in embryogenesis, placentation, metabolism, cardiovascular health, and neural development. Consequently, autophagy dysfunctions contribute to various pathological processes such as tumor progression, neurodegeneration,or heart failure [19,20]. Autophagy probably played also a major role in the evolution of the immune system, and it is now important for inflammasome activation, type I interferon production, antigen presentation, and pathogen degradation [21].
Three major forms of autophagy are described based on their mode of cargo delivery to lysosomes: (a) chaperon-mediated autophagy (CMA) involves chaperon recognition of cytosolic proteins that are directly translocated across the lysosomal membrane for degradation; (b) microautophagy consists of direct invagination of cytoplasm within a lysosome, and (c) macroautophagy involves isolation ofcytoplasmic particles or organelles within a double-membraned compartment that will subsequently fuse with lysosomes [22]. Pathogens can be selectively targeted for degradation by macroautophagy (called in this specific case xenophagy). LC3-associated phagocytosis (LAP) is a non-canonical autophagic pathwaythat also contributes to pathogen elimination. Macroautophagy/xenophagy and LAP share molecularactors such as evolutionary conserved autophagy-related (ATG) genes and phosphatidylinositol 3-kinaseclass III (PIK3C3), regulating the different autophagic steps [23].
Macroautophagy/xenophagy is initiated through the kinase AMP-dependent activation of the ULK1 serine threonine kinase complex (ULK1, FIP200, ATG13, ATG101). The activated ULK1 complex translocates to an isolated cellular membrane and becomes the phagophore nucleation site, where it phosphorylates multiple effectors (including itself) [24]. Within the targeted membrane, the autophagy-specific PIK3C3 synthesizes phosphatidylinositol-3-phosphate (PI3P) allowing the recruitment of specific effectors from the WIPI protein family, which participates in phagophore biosynthesis by recruiting ATG effectors [25]. The phagophore elongates thanks to two main ubiquitin-like systems: the Atg5-Atg12 conjugation system, and the microtubule-associated protein light chain 3 system (LC3, themammalian ortholog of the yeast protein Atg8) [26]. The elongated phagophore curves and fuses between its two ends, forming a double membrane compartment named autophagosome which encapsulates cytoplasmic organelles (or pathogens in the case of xenophagy). The autophagosome matures by fusing directly with lysosomes, building an acidic and degradative autophagolysosome or autolysosome. Before fusing with lysosomes, the autophagosome can fuse with endocytic compartments such as early and late endosomes, as well as multivesicular bodies, forming an intermediary compartment named amphisome [27].
LAP is directly activated by extracellular particles (pathogens or dead cells) binding to pathogen recognition receptors [28], receptors that detect phosphatidylserine (TIM4) [29], or antibodies (FcγR2a) [30]. As it happens in xenophagy, synthesis of PI3P by the PIK3C3 occurs but directly at the phagosomesite. Both ubiquitin-like conjugation systems Atg5-Atg12 and Atg8/LC3 are also implicated in the LAP pathway, forming a LC3-positive phagosome called LAPosome, which matures by fusing with lysosomesfor degradation [31].
LC3 is used as a hallmark of autophagy in many studies [32]. Indeed, LC3 is involved in different autophagic steps such as: phagophore elongation [33], tethering and membrane fusion [34], interaction with selective autophagy receptors, and autolysosome inner-membrane degradation [35]. The cytosolic form of LC3 is cleaved on its C-terminal part by the cysteine protease Atg4, exposing a glycine residue (LC3-I). The E1-like enzyme Atg7 transfers LC3-I to the E2-like enzyme Atg3. Then, Atg5-Atg12 facilitates the conjugation of the phosphatidylethanolamine to the C-terminal glycine. This allows LC3-II insertion in the inner and outer membrane of the autophagosome, or directly within the single membrane of the LAPosome [22]. LC3-II is then degraded within the autophagolysosome [36].
3. Yersinia pseudotuberculosis
In vitro analyses have demonstrated that
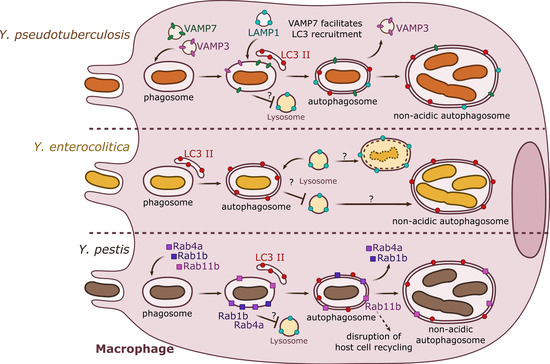
Y. pseudotuberculosis can establish an intracellular stage within macrophages by inducing autophagy (). Indeed, in murine bone marrow-derived macrophages (BMDMs), Moreau and colleagues (2010) [
37] showed that intracellular and metabolically active
Y. pseudotuberculosis trigger the autophagic machinery in a process independent of the pYV-encoded T3SS. Ultrastructural electron microscopy analyses showed that
Y. pseudotuberculosis survives and replicates within BMDMs inside autophagosomal compartments formed by double or multiple membranes. Fluorescence microscopy further demonstrated that during the course of infection,
Y. pseudotuberculosis-containing vacuoles (YCVs) enlarge progressively overtime by fusing with LC3-positive membranes, and harbor the lysosomal associated membrane protein 1 (LAMP1). Upon measuring of the autophagic flux (assessing the conversion of cytosolic LC3 I towards lipidated/membrane-associated LC3 II), as well as quantifying the levels of vacuolar LC3-GFP in
Y. pseudotuberculosis infected cells, the authors confirm that bacterial infection induces autophagy. Bacterial replication can be detected in
Atg5−/− (autophagic pathway impaired) mouse embryonic fibroblasts (MEFs), but is significantly lower to that observed in wild type MEFs, indicating that autophagy is required for optimal intracellular
Y. pseudotuberculosis growth. In experiments using BMDMs transfected with the inactive Atg4B C74A mutant of Atg4B (a processing protease for LC3), bacteria are found in acidic compartments labeled with LysoTracker. In wild type BMDMs, conversely,
Y. pseudotuberculosis is found in non-acidic compartments, indicating that bacteria are able to impair the maturation of YCVs into degradative compartments. This result confirms previous observations indicating that
Y. pseudotuberculosis inhibits the vacuolar proton ATPase (V-ATPase) [
38]. Interestingly, bacteria-free autophagosomes fuse with lysosomes, indicating that the basal autophagic flux is not impaired in infected macrophages [
37].
Figure 1. Summary results from independent studies of autophagy induction by Y. pseudotuberculosis, Y. enterocolitica, and Y. pestis in macrophages. Y. pseudotuberculosis (top) recruits VAMP3 and VAMP7 to the YCV early in the invasion process. VAMP7 facilitates the recruitment of LC3-autophagic membranes, forming an autophagosome with double or multiple membranes. Rab1b (not represented) is important for bacterial survival. Y. pseudotuberculosis survives and replicates in a non-acidic autophagosome. Y. enterocolitica (center) is present in a double- or multiple-membrane autophagosomal compartment positive for LC3. Depending on the strain and its pathogenicity, Y. enterocolitica seems to survive in autophagosomes. Y. pestis (bottom) targets Rab GTPases (1b, 4a, 11b) to the phagosome. Rab1b and Rab4a participate in the inhibition of acidification and thus are involved in bacterial survival. Rab11b is sequestered to the autophagosome over the course of infection, which leads to a global inhibition of host endosomal recycling. Y. pestis proliferates in a non-acidic autophagosome.
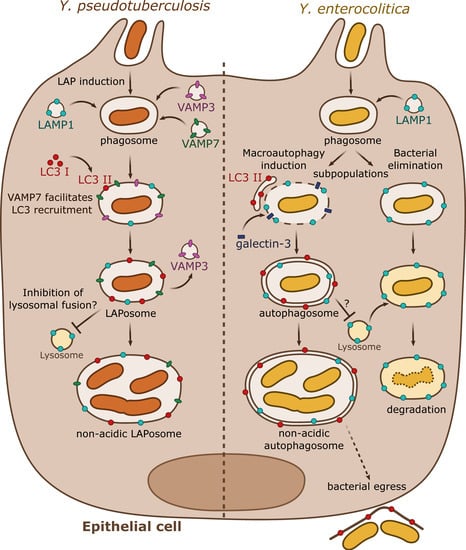
As demonstrated in macrophages, a subsequent study by the same group described that in epithelial HeLa cells,
Y. pseudotuberculosis also activates autophagy () [
39]. In these epithelial cells,
Y. pseudotuberculosis inhibits the maturation of YCVs, favoring bacterial proliferation in non-acidic compartments. However, and by contrast to what occurs in BMDMs, in HeLa cells, the non-conventional autophagic pathway LAP is engaged with direct recruitment of LC3 proteins to the YCVs. This process triggers the formation of a single-membrane niche for bacterial replication called LAPosome. Fluorescent microscopy experiments then revealed that VAMP3 and VAMP7 are sequentially recruited to the YCVs in both BMDMs and HeLa cells. siRNA inactivation assays demonstrated that VAMP7 facilitates the recruitment of LC3 to YCVs. Interestingly, VAMP3 is found in a higher proportion in YCVs in HeLa cells than in BMDMs. A combination of overexpression and siRNA experiments demonstrated that VAMP3 controls a checkpoint at which bacteria are committed to single LC3-positive compartments [
39].
Figure 2. Summary results from independent studies of autophagy subversion by enteropathogenic Yersinia in epithelial cells. Y. pseudotuberculosis (left) activates the LAP autophagic pathway during epithelial infection. VAMP3 and VAMP7 are sequentially targeted to the phagosome. VAMP3 favors the commitment towards a single-membrane compartment. VAMP7 participates to the recruitment of LC3-II directly to the phagosome forming the LAPosome. Y. pseudotuberculosis survives and multiplies within a non-acidic single membrane LAPosome. Y. enterocolitica (right) can survive or be degraded in epithelial cells. A subpopulation follows the lysosomal degradative pathway whereas the rest activates the macroautophagy pathway by recruiting LC3-positive autophagic membranes. Galectin-3, a marker of damaged endomembrane, is also recruited to some YCVs, suggesting their potential disruption. Y. enterocolitica survives and replicates in a non-acidic double or multiple membrane autophagosome blocked in its maturation process. The autophagosome seems to support bacterial egress without cell lysis.
4. Yersinia enterocolitica
As observed for
Y. pseudotuberculosis, Y. enterocolitica has been also shown to activate autophagy in macrophages and epithelial cells (; ). Deuretzbacher and colleagues (2009) [
40] demonstrated that in murine J774A.1 macrophages,
Y. enterocolitica WA cured from the pYV plasmid triggers the conversion of cytoplasmic LC3 I towards LC3 II. These authors observed intracellular bacteria in multimembrane compartments using electron microscopy as well as bacteria associated to LC3-GFP-positive vacuoles, observed using fluorescence microscopy. The activity of the T3SS, the absence of invasin, or chemical interference with invasin/β1 integrin-mediated signaling, which all lead to reduced bacterial intracellular numbers, reduce autophagy activation and bacterial association with autophagosomes. Interestingly, and opposed to what had been proposed for
Y. pseudotuberculosis in BMDMs, the authors suggest that autophagy does not favor the creation of an intracellular replication niche for
Y. enterocolitica in J774A.1 cells, and instead its activation promotes killing of intracellular bacterial. In the same line, chemical inhibition of autophagy or of vacuolar acidification leads to
Y. enterocolitica WA pYV-cured survival. Inhibition of autophagy by wild type
Y. enterocolitica involves the T3SS effector YopE, and targeting of Rho-GTPases. Of note, Connor et al. (2015) propose that
Y. enterocolitica 8081 pYV-cured is able to proliferate intracellularly within Raw264.7 macrophages, but the bacterial replication niche was not explored [
41].
More recently, Valencia-Lopez and colleagues (2019) [
42] explored infection of HeLa cells by the
Y. enterocolitica WA pYV-cured strain previously used to investigate macrophage infection. In this work, upon correlative-light electron microscopy studies, the authors indicate that bacteria are found in YCVs characterized by autophagy-related, ultrastructural features, including the presence of double or multiple membranes. By exploring infection of wild type HeLa cells or with FIP200 deficiency (which affects classical canonical autophagy, but not LAP) through correlative light/electron microscopy (CLEM), the authors conclude that canonical autophagy is the main cellular process subverted by
Y. enterocolitica (and not preferentially LAP, as proposed by Moreau et al. for
Y. pseudotuberculosis). Fluorescence microscopy indicates that YCVs are decorated by LC3-GFP as well as by the LC3 adapter p62 and ubiquitin. Interestingly, and as previously observed for
Y. pseudotuberculosis, Valencia-Lopez and co-authors demonstrate that
Y. enterocolitica also escapes autolysosomal maturation by inhibiting fusion with lysosomes, resulting in bacterial-containing, non-acidic compartments devoid of proteolytic activity, in which bacteria replicate. Dead bacteria are not able to trigger LC3-GFP recruitment to the YCV, and they are found in acidic compartments suggesting that the induction of autophagy and acidification inhibition are active processes requiring metabolically active bacteria. As opposed to observations performed by Deuretzbacher et al. in macrophages, invasin signaling is not sufficient for autophagy induction or inhibition of lysosomal fusion. Interestingly, the authors show that half of the internalized
Y. enterocolitica are present in phagosomes which do not engage autophagy and are LAMP-1 positive but devoid of LC3, indicating that this bacterial population is eliminated by a classical lysosomal pathway characterized by phagosomal acidification and proteolytic degradation [
40]. The authors also suggest that autophagy could be subverted by
Y. enterocolitica to promote a non-lytic egress from epithelial cells [
42].
5. Yersinia pestis
Replication of wild type
Y. pestis, or strains lacking pCD1, had been observed in mouse macrophages [
15,
43], and ultrastructural studies suggested that bacterial replication takes place within a phagolysosomal compartment [
44]. Pujol and colleagues (2009) [
45] determined that wild type
Y. pestis KIM5+ multiplies efficiently in BMDMs, and that a subpopulation of bacteria could be found in double membrane compartments. Autophagic flux analyses demonstrated that conversion of LC3 I towards LC3 II is augmented upon BMDM infection by
Y. pestis, and fluorescent microscopy identified recruitment of LC3-GFP to YCVs. As previously observed for enteropathogenic
Yersinia,
Y. pestis-containing vacuoles start as tight-fitting compartments, but progressively enlarge and become spacious vacuoles at late infection time points, which sustained bacterial replication. In addition, as previously observed for
Y. pseudotuberculosis, Y. pestis avoids acidification of its compartment. However, differently from what had been reported for
Y. pseudotuberculosis, Y. pestis survives equally well in BMDMs proficient (wild type) or deficient (ATG5 mutant) for autophagy, suggesting that
Y. pestis may not require autophagy for its survival in macrophages [
45].
Two recent studies have also implicated the small GTPases of the Rab family as key effectors allowing
Y. pestis to avoid killing by macrophages. Rab1 is normally associated to ER-to-Golgi trafficking, but the Rab1b isoform has been specifically involved in autophagosome formation [
46]. Connor and colleagues (2015) [
41] first showed that siRNA inactivation of Rab1b significantly reduced the survival of
Y. pestis CO92 pCD1-cured in RAW264.7 macrophages, while it does not affect bacterial entry. The authors then showed that Rab1b was required for
Y. pestis to block YCVs acidification as well as fusion with LAMP-1 positive compartments. Fluorescent microscopy experiments showed that Rab1b was directly recruited to YCVs, but surprisingly Rab1b siRNA inactivation did not affect
Y. pestis recruitment of LC3, suggesting that Rab1b controls a signaling cascade that is not directly related to autophagosome formation. A subsequent genome-wide siRNA screen performed by the same team (Connor et al. 2018) [
47] showed that the small GTPases Rab4 and Rab11b are also recruited to the YCVs and contribute to
Y. pestis survival in macrophages. Rab4a would cooperate together with Rab1b in the early steps of infection by inhibiting YCVs fusion with lysosomes, avoiding acidification. On the other hand, Rab11b, which remains sequestered by the YCVs, would allow disruption of the host cell endosomal recycling pathway, favoring bacterial replication at later infection time points () [
47].
6. Yersinia ruckeri
Another species from the Yersinia genus,Y. ruckeri, is a fish pathogen responsible for entericredmouth disease [48]. Experiments in fathead minnow-derived epithelial cells (FHM), in a liver cell line from the rainbow trout (R1), as well as in salmon kidney (ASK, SHK) and embryonic celllines (CHSE-214) indicate that Y. ruckeri has the capacity to invade host cells [49–51]. However, the intracellular stage appears to be transient, leading to host cell death [51] or bacterial degradation [50],suggesting that Y. ruckeri is not able to multiply or survive inside epithelial cultured cells. Interestingly, Ryckaert and colleagues (2010) demonstrated by gentamicin protection assays that Y. ruckeri can invadeand survive within rainbow trout macrophages [52]. Using transmission electron microscopy, Y. ruckeri is detected in infected macrophages within double membrane vacuoles, which display autophagosomalfeatures. Intracellular bacteria replicate and survive within these compartments for at least 24 h afterinfection [52]. Nevertheless, potential autophagic markers of this YCV, its formation, and maturationprocess remain to be deciphered.
 +1 credit
+1 credit