 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | C.-Thomas Bock | + 2116 word(s) | 2116 | 2020-11-17 06:42:38 |
Video Upload Options
Infection with hepatitis E virus (HEV) represents the most common source of viral hepatitis globally. Although infecting over 20 million people annually in endemic regions, with major outbreaks described since the 1950s, hepatitis E remains an underestimated disease.
1. Introduction
Hepatitis E virus (HEV) is a quasi-enveloped, positive strand RNA virus belonging to the family Hepeviridae [1]. HEV is the causative agent of Hepatitis E, the most common cause of acute viral hepatitis both in resource poor and developed countries. Hepatitis E presents as a mostly asymptomatic or acute self-limiting disease with a mortality rate up to 3% in young adults [2]. However the mortality rate may reach 30% in pregnant women [3]. Furthermore, chronic hepatitis E infections may occur in high-risk groups such as immunocompromised individuals (e.g., transplant recipients), those with pre-existing liver disease, HIV-positive persons, and cancer patients [4][5][6][7]. A recent study estimates that 939 million people worldwide have been infected with HEV in the past and that 15–110 million people have recent or ongoing infections [8]. According to the WHO, an estimated 3.3 million symptomatic hepatitis E cases occur each year in endemic areas with 44,000 related deaths [9].
HEV belongs to the genus Orthohepevirus, containing four species, namely Orthohepevirus A, B, C, and D with A containing the genotypes HEV-1 to HEV-8 [10]. The genotypes that infect humans include HEV-1 to -4, -7, and Orthohepevirus C, casually termed rat HEV [10][11][12][13]. HEV-1 and -2 infect only humans and cause large waterborne outbreaks due to contaminated drinking water in endemic regions of South and Southeast Asia, Africa, and Mexico [14]. HEV-3 and -4 infect both humans and animals, cause sporadic cases in developed countries, and are mainly spread through consumption or close contact with contaminated animal products [14][15]. HEV-7 and rat HEV infections are rarely reported. HEV-1 is mainly transmitted via the fecal–oral route, but also by vertical transmission from mother to child, from person to person, and by blood transfusions [14][16]. HEV-2 is transmitted via the fecal–oral route and human-to-human [14]. In contrast, HEV-3 and -4 are transmitted by transfusion of contaminated blood products, consuming contaminated shellfish, contact with infected animals, environmental contamination by animal manure run-off, and consumption of raw or undercooked meat [14][16]. Hepatitis E is not a sole health burden of the developing world, however, with numbers of reported sporadic cases increasing in industrialized nations, where the virus is spread primarily through zoonotic transmission.
Although usually a self-limiting disease in immunocompetent persons, hepatitis E can cause serious complications in at-risk populations such as pregnant women [17] and organ transplant recipients [5][18][19]. Treatment options remain limited, and only one vaccine has been developed so far with its use limited to China [20].
2. Discovery and History
An epidemic of HEV was reported in 1955 in Delhi, India, with about 29,000 cases of icteric hepatitis [21]. After this epidemic, several waterborne outbreaks were reported throughout India, and most of these cases were non-A and non-B, leading the disease to be described as enteric non-A non-B hepatitis (ENANBH) [21]. In addition, a major water-related epidemic outbreak in the Kashmir Valley was reported at the end of 1978, with 52,000 cases and 1700 deaths [22][23]. The symptoms of these cases were similar to hepatitis A but were negative for both hepatitis A and hepatitis B and were therefore confirmed as ENANBH [22]. In 1981, hepatitis occurred in a Soviet military camp in Afghanistan. To investigate this situation, a doctor in the Russian army, Mikhail Balayan, voluntarily ingested a pooled filtrate of stool samples from the infected soldiers, and he subsequently developed acute hepatitis [24]. The serum of Dr. Balayan was negative for hepatitis A virus (HAV) and hepatitis B virus (HBV), suggesting that a new pathogen was responsible for this infection. Immunoelectron microscopy identified 20–30 nm non-enveloped virus-like particles in stool, confirming a novel ENANBH virus [24]. In 1990, this novel ENANBH was partially cloned and sequenced and was henceforth called the hepatitis E virus (HEV) [25][26]. It was initially indicated that hepatitis E infection spreads via contaminated water and is limited to resource poor countries. Later, however, increasing reports of sporadic cases emerged in non-endemic industrialized countries with high seroprevalence in a few areas of the United States [27]. The reason for this high prevalence was not understood, and it was speculated that undetected non-pathogenic or less pathogenic HEV strains were circulating. In 1998, genome sequences confirmed that human HEV were similar to that of HEV in pigs, suggesting zoonotic transmission pathways [28][29]. Overall, these studies led to the identification of a broad spectrum of HEV strains that are either restricted to humans, animals, or infect both.
3. Virology
HEV particles have an icosahedral shape, are non-enveloped, and form virions with a diameter of about 27–34 nm [30]. The HEV genome is about 7.2 kb in size and consists of a 5′ UTR capped with 7-methylguanosine (M7G), followed by three open reading frames (ORF1, ORF2, and ORF3). The 3′ UTR ends with a poly(A) tail (An) like the eukaryotic mRNA structure. Viral replication starts with the translation of the ORF1-encoded non-structural polyprotein. An RNA-dependent RNA polymerase (RdRp) subsequently transcribes the full-length negative-sense RNA. This RNA serves as a template for the synthesis of two viral positive-sense RNAs in infected cells, a full-length genomic RNA, and a subgenomic RNA containing the capsid protein-encoding ORF2 and polyfunctional protein (PP)-encoding ORF3 [31]. The genomic arrangement and the stages of viral genome replication and protein synthesis are visualized in Figure 1.
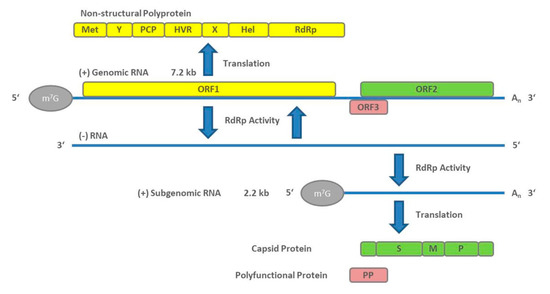
Figure 1. Genome arrangement of hepatitis E virus (HEV) and steps of viral genome replication.
ORF1 encodes a non-structural polyprotein of varying length that consists of seven domains comprising a methyl transferase (Met), X and Y domains, a papain-like cysteine protease (PCP), a proline-rich hypervariable region (HVR), RNA helicase (Hel), and RNA-dependent RNA polymerase (RdRp) [32]. Of these seven, only the Met, Hel, and RdRp have been functionally well characterized [33][34][35][36]. The complete function of all ORF1 domains is still not fully understood. In addition, whether it functions as a multi-domain polyprotein like a "Swiss army knife" or whether it needs to be cleaved for functional activity [37] is under debate and there is still no conclusive evidence that PCP has a protease function [38]. Although ORF1 is essential for HEV replication, its HVR displays considerable sequence divergence even between isolates of the same virus genotype [39]. Size differences between different HEV genomes can be primarily attributed to the HVR region. Analysis of different patient derived HEV isolates revealed various strains that harbor insertions from other regions of the viral genome, or from human genes, within the HVR [40].
The ORF2 encodes for the capsid protein. Its N-terminal signal peptide shuttles it to the extracellular compartment. The ORF2 protein contains three potential N-glycosylation sites [41][42][43], and is the main immunogenic target of neutralizing antibodies [41][44]. The full-length ORF2 encodes 660aa; however, recent reports suggested that ORF2 protein is processed into at least two forms, including one or two forms of secreted glycoproteins that are not associated with infectious particles, and one unglycosylated form which is the structural component of infectious particles [45][46]. ORF2 has been well characterized for its usefulness in diagnostics and vaccines, including the vaccine Hecolin® p239, which is currently only approved in China.
The ORF3 protein is a polyfunctional 13-kDa protein of 113 (genotype 3) or 114aa (genotypes 1, 2, and 4). Computational homology scans did not reveal any domains comparable to other known viral proteins. It has been shown to bind to microtubules and be involved in particle assembly and egress by interaction with the tumor susceptibility gene 101 protein (TSG101), a key protein involved in the endosomal sorting complexes of the ESCRT (endosomal sorting complexes required to transport) transport pathway and involved in the budding of the viruses [47][48][49][50]. Furthermore, it may play a role in infectious particle secretion via its palmitoylation and membrane association [51]. Additionally, there are also reports of its role in intracellular transduction pathways, the potential to reduce host immune responses, and protection of virus-infected cells [52][53][54]. A recent article reports that ORF3 is a functional ion channel required for release of infectious particles [55].
HEV exists as quasi-enveloped viral particles in blood and cell culture supernatant and as non-enveloped virions in bile and feces [56]. When they are shed into the environment, non-enveloped naked virions are enterically transmitted through contaminated water or food. So far, it is not well understood how HEV virions overcome the intestinal barrier and reach the liver. However, it is assumed that the virions first infect the enterocytes, where they multiply, and are excreted as quasi-enveloped virus particles into blood circulation, thus infecting hepatocytes [57]. On their way through the bile duct, the envelope is stripped off and naked, and more infectious virions are again released via the stool [56][58].
Naked HEV particles possibly attach to target cells via heparin sulfate proteoglycans (HSGPs) [59] and heat shock cognate protein 70 (HSC70) [60]. Integrin α3 has been described recently [61] as a candidate receptor to mediate entry into the cells by dynamin-dependent, clathrin-mediated endocytosis, supported by the GTPases Ras-related proteins Rab5A (RAB5) and Rab7a (RAB7), which are necessary for quasi-enveloped particle internalization [59][62][63]. Quasi-enveloped particles attach less efficiently to cells and likely enter the cell in a manner similar to exosomes [63]. Not requiring HSGPs allows attachment in a non-cell-specific manner, possibly explaining HEV’s capacity to infect extrahepatic tissues [63]. Following this, lysosomal degradation of the lipid membrane (in the case of enveloped particles) and subsequent viral capsid uncoating take place, followed by release of the genomic HEV positive strand RNA into the cytoplasm [63]. The host cellular transcriptional machinery translates ORF1 polyprotein containing RdRp from HEV RNA. The polymerase transcribes the full-length negative-sense HEV RNA. From this negative strand, two RNAs are transcribed by RNA helicases and RdRp to form a full-length genomic RNA and a 2.2 kb bicistronic subgenomic RNA. These two capped and polyadenylated RNAs serve as templates for the translation of non-structural ORF1 polyproteins, ORF2 capsid proteins, and polyfunctional ORF3 proteins [64].
The subsequent steps are viral assembly and release. ORF2 and ORF3 and positive-sense genomic HEV RNA are known to form a complex in the ER–Golgi intermediate compartment and produce viral progeny particles [65][66]. The particles of the progeny virus bind to the TSG101 protein and are secreted in a presumably basal fashion as enveloped particles [47][48]. When leaving hepatocytes from the apical part, the envelope is stripped as described above [56][58]. There are many significant gaps in the understanding of the HEV life cycle and virus–host cell interactions [67], and further studies are urgently needed. Several in vitro systems exist to study the virus. Reverse genetics models based on infectious cDNA clones have been described for several genotypes. The most commonly used are the Sar-55-related genotype 1 clone [68], the genotype 3a and 3c Kernow-C1- [69], and 47832-related [70] clones, respectively, both of which contain insertions in the HVR, and the genotype 4 TW6196 clone [71]. A recent presentation of a novel in vitro method to produce high viral titers, allowing study of the full HEV replication cycle in cell culture, has additionally created confidence that we may overcome our limited understanding of HEV pathophysiology [72]. Although these systems are most commonly used in conventional cell culture systems, several animal infection models have been developed. Rhesus monkeys have been shown to be susceptible to HEV-1 through -4 [28][73], while cynomolgous monkeys and chimpanzees have been used as models for HEV-1 and -2 [68][74]. As natural hosts for HEV-3 and -4, pigs can be readily infected by strains of these genotypes [75][76]. Recent reports have also described successful infection of human liver chimeric mice with HEV-1 and -3 strains [77][78]. Moreover, small animal and avian models exist for the study of animal HEV. Although animal infection models provide more physiological conditions than cell culture systems, limitations are also present. Neither non-human primates nor mice represent natural hosts of HEV, while pigs can only be infected with HEV-3 and -4. Experiments with primates also raise ethical concerns.
4. Conclusions
Infection with hepatitis E virus is still an under-reported disease worldwide. However, awareness of the prevalence of this virus has grown exponentially in the last decades. Before and since its discovery, there had been numerous large-scale outbreaks in the developing world with tens of thousands of cases, demonstrating that HEV is the most common cause of viral hepatitis worldwide. While HEV-1 and HEV-2 occur mainly in waterborne epidemics in areas with insufficient hygienic conditions, sporadic cases of HEV-3 and HEV-4, which are transmitted zoonotically or via contaminated blood products in industrialized countries, have confirmed that HEV is not a health burden solely of resource poor regions. Although much effort has been put into elucidating the mechanisms of infection, replication, and pathology of the virus, many questions are still left unanswered, underlining the importance of further research on its virology. This is especially important as hepatitis E infections—depending on the genotype—can cause severe liver disease in risk groups such as pregnant women and immunocompromised persons. With these aspects in mind, it is clear that further research is vital to develop more effective treatment options and approved vaccination strategies.
References
- Smith, D.B.; Simmonds, P.; Members Of The International Committee On The Taxonomy Of Viruses Hepeviridae Study, G.; Jameel, S.; Emerson, S.U.; Harrison, T.J.; Meng, X.J.; Okamoto, H.; Van der Poel, W.H.M.; Purdy, M.A. Consensus proposals for classification of the family Hepeviridae. J Gen Virol 2014, 95, 2223-2232, doi:10.1099/vir.0.068429-0.
- Teshale, E.H.; Hu, D.J. Hepatitis E: Epidemiology and prevention. World J Hepatol 2011, 3, 285-291, doi:10.4254/wjh.v3.i12.285.
- Perez-Gracia, M.T.; Suay-Garcia, B.; Mateos-Lindemann, M.L. Hepatitis E and pregnancy: current state. Rev Med Virol 2017, 27, e1929, doi:10.1002/rmv.1929.
- Dalton, H.R.; Stableforth, W.; Thurairajah, P.; Hazeldine, S.; Remnarace, R.; Usama, W.; Farrington, L.; Hamad, N.; Sieberhagen, C.; Ellis, V., et al. Autochthonous hepatitis E in Southwest England: natural history, complications and seasonal variation, and hepatitis E virus IgG seroprevalence in blood donors, the elderly and patients with chronic liver disease. Eur J Gastroenterol Hepatol 2008, 20, 784-790, doi:10.1097/MEG.0b013e3282f5195a.
- Kamar, N.; Selves, J.; Mansuy, J.M.; Ouezzani, L.; Peron, J.M.; Guitard, J.; Cointault, O.; Esposito, L.; Abravanel, F.; Danjoux, M., et al. Hepatitis E virus and chronic hepatitis in organ-transplant recipients. N Engl J Med 2008, 358, 811-817, doi:10.1056/NEJMoa0706992.
- Bai, M.J.; Zhou, N.; Dong, W.; Li, G.X.; Cong, W.; Zhu, X.Q. Seroprevalence and risk factors of hepatitis E virus infection in cancer patients in eastern China. Int J Infect Dis 2018, 71, 42-47, doi:10.1016/j.ijid.2018.04.003.
- Rivero-Juarez, A.; Lopez-Lopez, P.; Frias, M.; Rivero, A. Hepatitis E Infection in HIV-Infected Patients. Front Microbiol 2019, 10, 1425, doi:10.3389/fmicb.2019.01425.
- Li, P.; Liu, J.; Li, Y.; Su, J.; Ma, Z.; Bramer, W.M.; Cao, W.; de Man, R.A.; Peppelenbosch, M.P.; Pan, Q. The global epidemiology of hepatitis E virus infection: A systematic review and meta-analysis. Liver Int 2020, 40, 1516-1528, doi:10.1111/liv.14468.
- WHO. Hepatitis E: fact sheet. 2020. Availabe online: https://www.who.int/news-room/fact-sheets/detail/hepatitis-e (accessed on Oct 19, 2020).
- Smith, D.B.; Izopet, J.; Nicot, F.; Simmonds, P.; Jameel, S.; Meng, X.-J.; Norder, H.; Okamoto, H.; van der Poel, W.H.M.; Reuter, G., et al. Update: proposed reference sequences for subtypes of hepatitis E virus (species Orthohepevirus A). J Gen Virol 2020, 101, 692-698, doi:10.1099/jgv.0.001435.
- Lee, G.H.; Tan, B.H.; Teo, E.C.; Lim, S.G.; Dan, Y.Y.; Wee, A.; Aw, P.P.; Zhu, Y.; Hibberd, M.L.; Tan, C.K., et al. Chronic Infection With Camelid Hepatitis E Virus in a Liver Transplant Recipient Who Regularly Consumes Camel Meat and Milk. Gastroenterology 2016, 150, 355-357 e353, doi:10.1053/j.gastro.2015.10.048.
- Sridhar, S.; Yip, C.C.; Wu, S.; Chew, N.F.; Leung, K.H.; Chan, J.F.; Zhao, P.S.; Chan, W.M.; Poon, R.W.; Tsoi, H.W., et al. Transmission of rat hepatitis E virus infection to humans in Hong Kong: a clinical and epidemiological analysis. Hepatology 2020, 10.1002/hep.31138, doi:10.1002/hep.31138.
- Andonov, A.; Robbins, M.; Borlang, J.; Cao, J.; Hatchette, T.; Stueck, A.; Deschambault, Y.; Murnaghan, K.; Varga, J.; Johnston, L. Rat Hepatitis E Virus Linked to Severe Acute Hepatitis in an Immunocompetent Patient. J Infect Dis 2019, 220, 951-955, doi:10.1093/infdis/jiz025.
- Khuroo, M.S.; Khuroo, M.S.; Khuroo, N.S. Transmission of Hepatitis E Virus in Developing Countries. Viruses 2016, 8, 253, doi:10.3390/v8090253.
- Wang, B.; Meng, X.J. Hepatitis E virus: host tropism and zoonotic infection. Curr Opin Microbiol 2020, 59, 8-15, doi:10.1016/j.mib.2020.07.004.
- Mirazo, S.; Ramos, N.; Mainardi, V.; Gerona, S.; Arbiza, J. Transmission, diagnosis, and management of hepatitis E: an update. Hepat Med 2014, 6, 45-59, doi:10.2147/HMER.S63417.
- Patra, S.; Kumar, A.; Trivedi, S.S.; Puri, M.; Sarin, S.K. Maternal and fetal outcomes in pregnant women with acute hepatitis E virus infection. Ann Intern Med 2007, 147, 28-33, doi:10.7326/0003-4819-147-1-200707030-00005.
- Gérolami, R.; Moal, V.; Colson, P. Chronic hepatitis E with cirrhosis in a kidney-transplant recipient. N Engl J Med 2008, 358, 859-860, doi:10.1056/NEJMc0708687.
- Haagsma, E.B.; van den Berg, A.P.; Porte, R.J.; Benne, C.A.; Vennema, H.; Reimerink, J.H.; Koopmans, M.P. Chronic hepatitis E virus infection in liver transplant recipients. Liver Transpl 2008, 14, 547-553, doi:10.1002/lt.21480.
- Zhu, F.C.; Zhang, J.; Zhang, X.F.; Zhou, C.; Wang, Z.Z.; Huang, S.J.; Wang, H.; Yang, C.L.; Jiang, H.M.; Cai, J.P., et al. Efficacy and safety of a recombinant hepatitis E vaccine in healthy adults: a large-scale, randomised, double-blind placebo-controlled, phase 3 trial. Lancet 2010, 376, 895-902, doi:10.1016/S0140-6736(10)61030-6.
- Arankalle, V.A.; Chadha, M.S.; Tsarev, S.A.; Emerson, S.U.; Risbud, A.R.; Banerjee, K.; Purcell, R.H. Seroepidemiology of water-borne hepatitis in India and evidence for a third enterically-transmitted hepatitis agent. Proc Natl Acad Sci U S A 1994, 91, 3428-3432, doi:10.1073/pnas.91.8.3428.
- Khuroo, M.S. Hepatitis E: the enterically transmitted non-A, non-B hepatitis. Indian J Gastroenterol 1991, 10, 96-100, doi:10.1101/cshperspect.a033449.
- Khuroo, M.S. Study of an epidemic of non-A, non-B hepatitis. Possibility of another human hepatitis virus distinct from post-transfusion non-A, non-B type. Am J Med 1980, 68, 818-824, doi:10.1016/0002-9343(80)90200-4.
- Balayan, M.S.; Andjaparidze, A.G.; Savinskaya, S.S.; Ketiladze, E.S.; Braginsky, D.M.; Savinov, A.P.; Poleschuk, V.F. Evidence for a virus in non-A, non-B hepatitis transmitted via the fecal-oral route. Intervirology 1983, 20, 23-31, doi:10.1159/000149370.
- Tam, A.W.; Smith, M.M.; Guerra, M.E.; Huang, C.C.; Bradley, D.W.; Fry, K.E.; Reyes, G.R. Hepatitis E virus (HEV): molecular cloning and sequencing of the full-length viral genome. Virology 1991, 185, 120-131, doi:10.1016/0042-6822(91)90760-9.
- Reyes, G.R.; Purdy, M.A.; Kim, J.P.; Luk, K.C.; Young, L.M.; Fry, K.E.; Bradley, D.W. Isolation of a cDNA from the virus responsible for enterically transmitted non-A, non-B hepatitis. Science 1990, 247, 1335-1339, doi:10.1126/science.2107574.
- Mast, E.E.; Kuramoto, I.K.; Favorov, M.O.; Schoening, V.R.; Burkholder, B.T.; Shapiro, C.N.; Holland, P.V. Prevalence of and risk factors for antibody to hepatitis E virus seroreactivity among blood donors in Northern California. J Infect Dis 1997, 176, 34-40, doi:10.1086/514037.
- Meng, X.J.; Halbur, P.G.; Shapiro, M.S.; Govindarajan, S.; Bruna, J.D.; Mushahwar, I.K.; Purcell, R.H.; Emerson, S.U. Genetic and experimental evidence for cross-species infection by swine hepatitis E virus. J Virol 1998, 72, 9714-9721, doi:10.1128/jvi.72.12.9714-9721.1998.
- Meng, X.J.; Purcell, R.H.; Halbur, P.G.; Lehman, J.R.; Webb, D.M.; Tsareva, T.S.; Haynes, J.S.; Thacker, B.J.; Emerson, S.U. A novel virus in swine is closely related to the human hepatitis E virus. Proc Natl Acad Sci U S A 1997, 94, 9860-9865, doi:10.1073/pnas.94.18.9860.
- Bradley, D.; Andjaparidze, A.; Cook, E.H., Jr.; McCaustland, K.; Balayan, M.; Stetler, H.; Velazquez, O.; Robertson, B.; Humphrey, C.; Kane, M., et al. Aetiological agent of enterically transmitted non-A, non-B hepatitis. J Gen Virol 1988, 69 ( Pt 3), 731-738, doi:10.1099/0022-1317-69-3-731.
- Graff, J.; Torian, U.; Nguyen, H.; Emerson, S.U. A bicistronic subgenomic mRNA encodes both the ORF2 and ORF3 proteins of hepatitis E virus. J Virol 2006, 80, 5919-5926, doi:10.1128/jvi.00046-06.
- Koonin, E.V.; Gorbalenya, A.E.; Purdy, M.A.; Rozanov, M.N.; Reyes, G.R.; Bradley, D.W. Computer-assisted assignment of functional domains in the nonstructural polyprotein of hepatitis E virus: delineation of an additional group of positive-strand RNA plant and animal viruses. Proc Natl Acad Sci U S A 1992, 89, 8259-8263, doi:10.1073/pnas.89.17.8259.
- Karpe, Y.A.; Lole, K.S. Deubiquitination activity associated with hepatitis E virus putative papain-like cysteine protease. J Gen Virol 2011, 92, 2088-2092, doi:10.1099/vir.0.033738-0.
- Magden, J.; Takeda, N.; Li, T.; Auvinen, P.; Ahola, T.; Miyamura, T.; Merits, A.; Kaariainen, L. Virus-specific mRNA capping enzyme encoded by hepatitis E virus. J Virol 2001, 75, 6249-6255, doi:10.1128/JVI.75.14.6249-6255.2001.
- Karpe, Y.A.; Lole, K.S. RNA 5'-triphosphatase activity of the hepatitis E virus helicase domain. J Virol 2010, 84, 9637-9641, doi:10.1128/JVI.00492-10.
- Agrawal, S.; Gupta, D.; Panda, S.K. The 3' end of hepatitis E virus (HEV) genome binds specifically to the viral RNA-dependent RNA polymerase (RdRp). Virology 2001, 282, 87-101, doi:10.1006/viro.2000.0819.
- Parvez, M.K. Molecular characterization of hepatitis E virus ORF1 gene supports a papain-like cysteine protease (PCP)-domain activity. Virus Res 2013, 178, 553-556, doi:10.1016/j.virusres.2013.07.020.
- Parvez, M.K.; Khan, A.A. Molecular modeling and analysis of hepatitis E virus (HEV) papain-like cysteine protease. Virus Res 2014, 179, 220-224, doi:10.1016/j.virusres.2013.11.016.
- Smith, D.B.; Vanek, J.; Ramalingam, S.; Johannessen, I.; Templeton, K.; Simmonds, P. Evolution of the hepatitis E virus hypervariable region. J Gen Virol 2012, 93, 2408-2418, doi:10.1099/vir.0.045351-0.
- Lhomme, S.; Nicot, F.; Jeanne, N.; Dimeglio, C.; Roulet, A.; Lefebvre, C.; Carcenac, R.; Manno, M.; Dubois, M.; Peron, J.M., et al. Insertions and Duplications in the Polyproline Region of the Hepatitis E Virus. Front Microbiol 2020, 11, 1, doi:10.3389/fmicb.2020.00001.
- Jameel, S.; Zafrullah, M.; Ozdener, M.H.; Panda, S.K. Expression in animal cells and characterization of the hepatitis E virus structural proteins. J Virol 1996, 70, 207-216, doi:10.1128/jvi.70.1.207-216.1996.
- Torresi, J.; Li, F.; Locarnini, S.A.; Anderson, D.A. Only the non-glycosylated fraction of hepatitis E virus capsid (open reading frame 2) protein is stable in mammalian cells. J Gen Virol 1999, 80 ( Pt 5), 1185-1188, doi:10.1099/0022-1317-80-5-1185.
- Zafrullah, M.; Ozdener, M.H.; Kumar, R.; Panda, S.K.; Jameel, S. Mutational analysis of glycosylation, membrane translocation, and cell surface expression of the hepatitis E virus ORF2 protein. J Virol 1999, 73, 4074-4082, doi:10.1128/JVI.73.5.4074-4082.1999.
- Zhou, Y.H.; Purcell, R.H.; Emerson, S.U. A truncated ORF2 protein contains the most immunogenic site on ORF2: antibody responses to non-vaccine sequences following challenge of vaccinated and non-vaccinated macaques with hepatitis E virus. Vaccine 2005, 23, 3157-3165, doi:10.1016/j.vaccine.2004.12.020.
- Yin, X.; Ying, D.; Lhomme, S.; Tang, Z.; Walker, C.M.; Xia, N.; Zheng, Z.; Feng, Z. Origin, antigenicity, and function of a secreted form of ORF2 in hepatitis E virus infection. Proc Natl Acad Sci U S A 2018, 115, 4773-4778, doi:10.1073/pnas.1721345115.
- Montpellier, C.; Wychowski, C.; Sayed, I.M.; Meunier, J.C.; Saliou, J.M.; Ankavay, M.; Bull, A.; Pillez, A.; Abravanel, F.; Helle, F., et al. Hepatitis E Virus Lifecycle and Identification of 3 Forms of the ORF2 Capsid Protein. Gastroenterology 2018, 154, 211-223 e218, doi:10.1053/j.gastro.2017.09.020.
- Nagashima, S.; Takahashi, M.; Jirintai; Tanaka, T.; Yamada, K.; Nishizawa, T.; Okamoto, H. A PSAP motif in the ORF3 protein of hepatitis E virus is necessary for virion release from infected cells. J Gen Virol 2011, 92, 269-278, doi:10.1099/vir.0.025791-0.
- Nagashima, S.; Takahashi, M.; Jirintai, S.; Tanaka, T.; Nishizawa, T.; Yasuda, J.; Okamoto, H. Tumour susceptibility gene 101 and the vacuolar protein sorting pathway are required for the release of hepatitis E virions. J Gen Virol 2011, 92, 2838-2848, doi:10.1099/vir.0.035378-0.
- Kannan, H.; Fan, S.; Patel, D.; Bossis, I.; Zhang, Y.J. The hepatitis E virus open reading frame 3 product interacts with microtubules and interferes with their dynamics. J Virol 2009, 83, 6375-6382, doi:10.1128/JVI.02571-08.
- Nagashima, S.; Jirintai, S.; Takahashi, M.; Kobayashi, T.; Tanggis; Nishizawa, T.; Kouki, T.; Yashiro, T.; Okamoto, H. Hepatitis E virus egress depends on the exosomal pathway, with secretory exosomes derived from multivesicular bodies. J Gen Virol 2014, 95, 2166-2175, doi:10.1099/vir.0.066910-0.
- Gouttenoire, J.; Pollán, A.; Abrami, L.; Oechslin, N.; Mauron, J.; Matter, M.; Oppliger, J.; Szkolnicka, D.; Dao Thi, V.L.; van der Goot, F.G., et al. Palmitoylation mediates membrane association of hepatitis E virus ORF3 protein and is required for infectious particle secretion. PLoS Pathog 2018, 14, e1007471, doi:10.1371/journal.ppat.1007471.
- Chandra, V.; Kar-Roy, A.; Kumari, S.; Mayor, S.; Jameel, S. The hepatitis E virus ORF3 protein modulates epidermal growth factor receptor trafficking, STAT3 translocation, and the acute-phase response. J Virol 2008, 82, 7100-7110, doi:10.1128/JVI.00403-08.
- He, M.; Wang, M.; Huang, Y.; Peng, W.; Zheng, Z.; Xia, N.; Xu, J.; Tian, D. The ORF3 Protein of Genotype 1 Hepatitis E Virus Suppresses TLR3-induced NF-kappaB Signaling via TRADD and RIP1. Sci Rep 2016, 6, 27597, doi:10.1038/srep27597.
- Xu, J.; Wu, F.; Tian, D.; Wang, J.; Zheng, Z.; Xia, N. Open reading frame 3 of genotype 1 hepatitis E virus inhibits nuclear factor-kappaappa B signaling induced by tumor necrosis factor-alpha in human A549 lung epithelial cells. PLoS One 2014, 9, e100787, doi:10.1371/journal.pone.0100787.
- Ding, Q.; Heller, B.; Capuccino, J.M.; Song, B.; Nimgaonkar, I.; Hrebikova, G.; Contreras, J.E.; Ploss, A. Hepatitis E virus ORF3 is a functional ion channel required for release of infectious particles. Proc Natl Acad Sci U S A 2017, 114, 1147-1152, doi:10.1073/pnas.1614955114.
- Nagashima, S.; Takahashi, M.; Kobayashi, T.; Nishizawa, T.; Nishiyama, T.; Primadharsini, P.P.; Okamoto, H. Characterization of the Quasi-Enveloped Hepatitis E Virus Particles Released by the Cellular Exosomal Pathway. J Virol 2017, 91, e00822-17, doi:10.1128/jvi.00822-17.
- Marion, O.; Lhomme, S.; Nayrac, M.; Dubois, M.; Pucelle, M.; Requena, M.; Migueres, M.; Abravanel, F.; Peron, J.M.; Carrere, N., et al. Hepatitis E virus replication in human intestinal cells. Gut 2019, 69, 901-910,10.1136/gutjnl-2019-319004, doi:10.1136/gutjnl-2019-319004.
- Smith, W. The action of bile salts on viruses. The Journal of Pathology and Bacteriology 1939, 48, 557-571, doi:10.1002/path.1700480309.
- Kalia, M.; Chandra, V.; Rahman, S.A.; Sehgal, D.; Jameel, S. Heparan sulfate proteoglycans are required for cellular binding of the hepatitis E virus ORF2 capsid protein and for viral infection. J Virol 2009, 83, 12714-12724, doi:10.1128/jvi.00717-09.
- Zhou, Y.; Emerson, S. P. 302 Heat shock cognate protein 70 may mediate the entry of hepatitis E virus into host cells. J Clin Virol 2006, 36, S155, doi:10.1016/S1386-6532(06)80480-4.
- Shiota, T.; Li, T.C.; Nishimura, Y.; Yoshizaki, S.; Sugiyama, R.; Shimojima, M.; Saijo, M.; Shimizu, H.; Suzuki, R.; Wakita, T., et al. Integrin alpha3 is involved in non-enveloped hepatitis E virus infection. Virology 2019, 536, 119-124, doi:10.1016/j.virol.2019.07.025.
- Holla, P.; Ahmad, I.; Ahmed, Z.; Jameel, S. Hepatitis E virus enters liver cells through a dynamin-2, clathrin and membrane cholesterol-dependent pathway. Traffic 2015, 16, 398-416, doi:10.1111/tra.12260.
- Yin, X.; Ambardekar, C.; Lu, Y.; Feng, Z. Distinct Entry Mechanisms for Nonenveloped and Quasi-Enveloped Hepatitis E Viruses. J Virol 2016, 90, 4232-4242, doi:10.1128/jvi.02804-15.
- Debing, Y.; Moradpour, D.; Neyts, J.; Gouttenoire, J. Update on hepatitis E virology: Implications for clinical practice. J Hepatol 2016, 65, 200-212, doi:10.1016/j.jhep.2016.02.045.
- Perttila, J.; Spuul, P.; Ahola, T. Early secretory pathway localization and lack of processing for hepatitis E virus replication protein pORF1. J Gen Virol 2013, 94, 807-816, doi:10.1099/vir.0.049577-0.
- Rehman, S.; Kapur, N.; Durgapal, H.; Panda, S.K. Subcellular localization of hepatitis E virus (HEV) replicase. Virology 2008, 370, 77-92, doi:10.1016/j.virol.2007.07.036.
- Wissing, M.H.; Bruggemann, Y.; Steinmann, E.; Todt, D. Virus-Host Cell Interplay during Hepatitis E Virus Infection. Trends Microbiol 2020, 10.1016/j.tim.2020.07.002, doi:10.1016/j.tim.2020.07.002.
- Emerson, S.U.; Zhang, M.; Meng, X.J.; Nguyen, H.; St Claire, M.; Govindarajan, S.; Huang, Y.K.; Purcell, R.H. Recombinant hepatitis E virus genomes infectious for primates: importance of capping and discovery of a cis-reactive element. Proc Natl Acad Sci U S A 2001, 98, 15270-15275, doi:10.1073/pnas.251555098.
- Shukla, P.; Nguyen, H.T.; Torian, U.; Engle, R.E.; Faulk, K.; Dalton, H.R.; Bendall, R.P.; Keane, F.E.; Purcell, R.H.; Emerson, S.U. Cross-species infections of cultured cells by hepatitis E virus and discovery of an infectious virus-host recombinant. Proc Natl Acad Sci U S A 2011, 108, 2438-2443, doi:10.1073/pnas.1018878108.
- Scholz, J.; Bachlein, C.; Gadicherla, A.K.; Falkenhagen, A.; Tausch, S.H.; Johne, R. Establishment of a Plasmid-Based Reverse Genetics System for the Cell Culture-Adapted Hepatitis E Virus Genotype 3c Strain 47832c. Pathogens 2020, 9, doi:10.3390/pathogens9030157.
- Córdoba, L.; Feagins, A.R.; Opriessnig, T.; Cossaboom, C.M.; Dryman, B.A.; Huang, Y.W.; Meng, X.J. Rescue of a genotype 4 human hepatitis E virus from cloned cDNA and characterization of intergenotypic chimeric viruses in cultured human liver cells and in pigs. J Gen Virol 2012, 93, 2183-2194, doi:10.1099/vir.0.043711-0.
- Todt, D.; Friesland, M.; Moeller, N.; Praditya, D.; Kinast, V.; Bruggemann, Y.; Knegendorf, L.; Burkard, T.; Steinmann, J.; Burm, R., et al. Robust hepatitis E virus infection and transcriptional response in human hepatocytes. Proc Natl Acad Sci U S A 2020, 117, 1731-1741, doi:10.1073/pnas.1912307117.
- Purcell, R.H.; Nguyen, H.; Shapiro, M.; Engle, R.E.; Govindarajan, S.; Blackwelder, W.C.; Wong, D.C.; Prieels, J.P.; Emerson, S.U. Pre-clinical immunogenicity and efficacy trial of a recombinant hepatitis E vaccine. Vaccine 2003, 21, 2607-2615, doi:10.1016/s0264-410x(03)00100-2.
- Tsarev, S.A.; Emerson, S.U.; Tsareva, T.S.; Yarbough, P.O.; Lewis, M.; Govindarajan, S.; Reyes, G.R.; Shapiro, M.; Purcell, R.H. Variation in course of hepatitis E in experimentally infected cynomolgus monkeys. J Infect Dis 1993, 167, 1302-1306, doi:10.1093/infdis/167.6.1302.
- Halbur, P.G.; Kasorndorkbua, C.; Gilbert, C.; Guenette, D.; Potters, M.B.; Purcell, R.H.; Emerson, S.U.; Toth, T.E.; Meng, X.J. Comparative pathogenesis of infection of pigs with hepatitis E viruses recovered from a pig and a human. J Clin Microbiol 2001, 39, 918-923, doi:10.1128/jcm.39.3.918-923.2001.
- Feagins, A.R.; Opriessnig, T.; Huang, Y.W.; Halbur, P.G.; Meng, X.J. Cross-species infection of specific-pathogen-free pigs by a genotype 4 strain of human hepatitis E virus. J Med Virol 2008, 80, 1379-1386, doi:10.1002/jmv.21223.
- Allweiss, L.; Gass, S.; Giersch, K.; Groth, A.; Kah, J.; Volz, T.; Rapp, G.; Schöbel, A.; Lohse, A.W.; Polywka, S., et al. Human liver chimeric mice as a new model of chronic hepatitis E virus infection and preclinical drug evaluation. J Hepatol 2016, 64, 1033-1040, doi:10.1016/j.jhep.2016.01.011.
- Sayed, I.M.; Verhoye, L.; Cocquerel, L.; Abravanel, F.; Foquet, L.; Montpellier, C.; Debing, Y.; Farhoudi, A.; Wychowski, C.; Dubuisson, J., et al. Study of hepatitis E virus infection of genotype 1 and 3 in mice with humanised liver. Gut 2017, 66, 920-929, doi:10.1136/gutjnl-2015-311109.

