 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Amnon Erez | -- | 1238 | 2024-03-20 16:15:14 | | | |
| 2 | Camila Xu | Meta information modification | 1238 | 2024-03-21 03:01:08 | | |
Video Upload Options
Dormancy is defined as the sum of processes that create a situation wherein embryonic tissues are unable to grow, even under environmental conditions that generally favor growth and development. The effects of temperature to break dormancy, from work done on peach, can be summarized as follows: Effective temperatures are between −2 °C and 13 °C, and the most effective being 4–8 °C with reduced efficiency at higher and lower temperatures. Moderate temperatures between 13 °C and 16 °C that will not break dormancy alone, when occurring in a daily cycle after previous chilling, enhance the effect of chilling. On the other hand, temperatures higher than 18 °C in a daily cycle will nullify former chilling. This negative effect of high temperatures increases the longer the duration and the higher the temperature. However, when cycles are longer than a day, the chilling effect is final and cannot be nullified by high temperatures. Apart from the effects of temperature on dormancy, light effects on dormancy in peach trials had also been detected Clearly, the dormant vegetative buds perceive light signals and react to light during dormancy and bud breaks. On the other hand, flower buds in peach were found to be non-responsive to light. Dormancy in vegetative buds is induced by short days. During endodormancy, the limitation of light and even total darkness enhance bud breaks in spring, compared to buds receiving natural light. But darkness in spring will prevent vegetative bud break even following sufficient chilling during winter. So, there is an analogy between chilling and darkness.
1. Introduction
2. Temperatures Effect on Dormancy
-
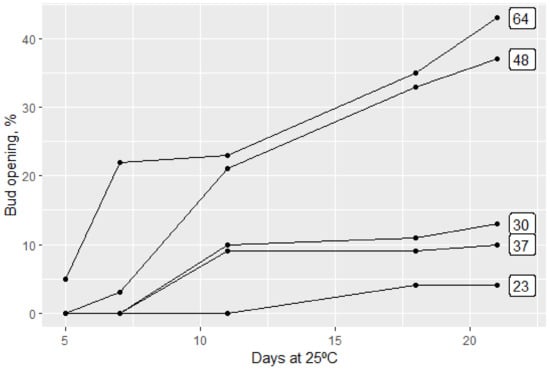
Effective temperatures to break dormancy in the peach are between −2 °C and 13 °C, and the most effective being 4–8 °C with reduced efficiency at higher and lower temperatures.
-
Moderate temperatures between 13 °C and 16 °C that will not break dormancy alone, when occurring in a daily cycle after previous chilling, enhance the effect of chilling. On the other hand, temperatures higher than 18 °C in a daily cycle will nullify former chilling.
-
This negative effect of high temperatures increases the longer the duration and the higher the temperature. However, when cycles are longer than a day, the chilling effect is final and cannot be nullified by high temperatures.
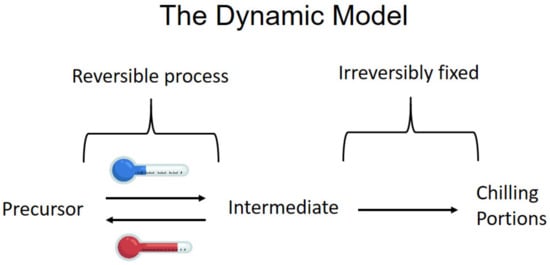
3. Light Effects on Dormancy
References
- Samish, R.M. The use of dinitrocresol-mineral oil sprays for the control of prolonged rest in apple orchards. J. Pomol. Hortic. Sci. 1945, 21, 164–179.
- Samish, R.M.; Lavee, S. The chilling requirement of fruit trees. Proc. XVI Int. Hort. Cong. 1962, 5, 372–388.
- Erez, A. Means to compensate for insufficient chilling to improve bloom and leafing. Acta Hortic. 1995, 395, 81–96.
- Erez, A.; Lavee, S. Breaking the dormancy of deciduous fruit trees in subtropical climates. Proc. XIX Inter. Hort. Cong. 1974, 3, 69–78.
- Erez, A.; Zur, A. Breaking the rest of apple buds by narrow-distillation-range oil and dinitro-o-cresol. Sci. Hortic. 1981, 14, 47–54.
- Erez, A.; Yablowitz, Z. Effect of dormancy agents with armobreak in the peach. Acta Hort. 1997, 441, 183–190.
- Erez, A.; Yablowitz, Z.; Aronovitz, A.; Hadar, A. Dormancy breaking chemicals: Efficiency with reduced phytotoxicity. Acta Hort. 2008, 772, 105–112.
- Crane, O.; Sar Shalom, A.; Erez, A. Coping with global warming effects on reduced winter chilling for deciduous fruit trees. Acta Hort. 2022, 1346, 91–100.
- Lang, G.A.; Early, J.D.; Martin, G.C.; Darnell, R.L. Endo-, para-, and ecodormancy: Physiological terminology and classification for dormancy research. HortScience 1987, 22, 371–377.
- Faust, M.; Erez, A.; Rowland, L.J.; Wang, S.Y.; Norman, H.A. Bud dormancy in perennial fruit trees; Physiological basis for dormancy induction maintenance and release. HortScience 1997, 32, 623–629.
- Erez, A.; Couvillon, G.A.; Hendershott, C.H. Quantitative chilling enhancement and negation in peach buds by high temperatures in a daily cycle1. J. Am. Soc. Hortic. Sci. 1979, 104, 536–540.
- Erez, A.; Couvillon, G.A.; Hendershott, C.H. The effect of cycle length on chilling negation by high temperatures in dormant peach leaf buds1. J. Am. Soc. Hortic. Sci. 1979, 104, 573–576.
- Erez, A.; Couvillon, G.A. Characterization of the influence of moderate temperatures on rest completion in peach. J. Am. Soc. Hort. Sci. 1987, 112, 677–680.
- Couvillon, G.A.; Erez, A. Effect of level and duration of high temperatures on rest in the peach. J. Am. Soc. Hortic. Sci. 1985, 110, 579–581.
- Weinberdger, J.H. Chilling requirements of peach varieties. Proc. Amer. Soc. Hort. Sci. 1950, 56, 122–128.
- Richardson, E.A.; Seeley, S.D.; Walker, D.R. A model for estimating the completion of rest for Redhaven and Elberta peach trees. Hortscience 1974, 9, 331–332.
- Fishman, S.; Erez, A.; Couvillon, G.A. The temperature dependence of dormancy breaking in plants: Mathematical analysis of a two-step model involving a cooperative transition. J. Theor. Biol. 1987, 124, 473–483.
- Fishman, S.; Erez, A.; Couvillon, G.A. The temperature dependence of dormancy breaking in plants: Simulation of processes studied under controlled temperatures. J. Theor. Biol. 1987, 126, 309–322.
- Erez, A.; Couvillon, G.A. Evaporative cooling to improve rest breaking of nectarine buds by counteracting high daytime temperatures. HortScience 1983, 18, 480–481.
- Luedeling, E.; Zhang, M.; McGranahn, G.; Leslie, C. Validation of winter chill models using historic records of walnut phenology. Agric. Forest Meteor. 2009, 149, 1854–1864.
- Luedeling, E. Climate change impacts on winter chill for temperate fruit and nut production: A review. Sci. Hortic. 2012, 144, 218–229.
- Erez, A.; Samish, R.M.; Lavee, S. The role of light in leaf and flower bud break of the peach (Prunus persica). Physiol. Plant. 1966, 19, 650–659.
- Erez, A.; Lavee, S.; Samish, R.M. The Effect of limitation in light during the rest period on leaf bud break of the peach (Prunus persica). Physiol. Plant. 1968, 21, 759–764.


