Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Kong, J.; Lyu, H.; Ouyang, Q.; Shi, H.; Zhang, R.; Xiao, S.; Guo, D.; Zhang, Q.; Chen, X.; Zhou, C.; et al. Regulation of NcRNAs on Ferroptosis. Encyclopedia. Available online: https://encyclopedia.pub/entry/56133 (accessed on 25 June 2026).
Kong J, Lyu H, Ouyang Q, Shi H, Zhang R, Xiao S, et al. Regulation of NcRNAs on Ferroptosis. Encyclopedia. Available at: https://encyclopedia.pub/entry/56133. Accessed June 25, 2026.
Kong, Jinghua, Hao Lyu, Qian Ouyang, Hao Shi, Rui Zhang, Shuai Xiao, Dong Guo, Qi Zhang, Xing-Zhen Chen, Cefan Zhou, et al. "Regulation of NcRNAs on Ferroptosis" Encyclopedia, https://encyclopedia.pub/entry/56133 (accessed June 25, 2026).
Kong, J., Lyu, H., Ouyang, Q., Shi, H., Zhang, R., Xiao, S., Guo, D., Zhang, Q., Chen, X., Zhou, C., & Tang, J. (2024, March 12). Regulation of NcRNAs on Ferroptosis. In Encyclopedia. https://encyclopedia.pub/entry/56133
Kong, Jinghua, et al. "Regulation of NcRNAs on Ferroptosis." Encyclopedia. Web. 12 March, 2024.
Copy Citation
Ferroptosis is a non-apoptotic mode of cell death driven by membrane lipid peroxidation and is characterized by elevated intracellular levels of Fe2+, ROS, and lipid peroxidation. Studies have shown that ferroptosis is related to the development of multiple diseases, such as cancer, neurodegenerative diseases, and acute myeloid leukemia. Ferroptosis plays a dual role in the occurrence and development of these diseases. Ferroptosis mainly involves iron metabolism, ROS, and lipid metabolism. Various mechanisms, including epigenetic regulation, have been reported to be deeply involved in ferroptosis. Abnormal epigenetic modifications have been reported to promote tumor onset or other diseases and resistance to chemotherapy drugs.
DNA methylation
RNA methylation
non-coding RNA
histone modification
1. Introduction
Non-coding RNAs (NcRNAs) play a significant role in regulating cellular processes. NcRNAs are non-coding transcripts with limited protein-coding potential and exert essential cellular functions through different molecular mechanisms [1]. In a broad sense, they can be subdivided into short and long ncRNAs and their biological functions, including short-stranded microRNAs (miRNAs), long-stranded non-coding RNAs (lncRNAs), and circular RNAs (circRNAs) [2][3].
MiRNAs are small ncRNAs, about 22 nucleotides long, widely found in eukaryotes and conserved throughout evolution. Single miRNAs can regulate multiple target genes, and multiple miRNAs can also regulate the same gene. The main function of miRNAs is the post-transcriptional regulation of gene expression by binding to complementary target mRNA sequences, leading to translational repression or mRNA degradation that halts protein synthesis [4][5]. It has been shown that miRNAs may also induce gene expression by binding to target sequences and acting as translation activators [6]. Although miRNAs were not known over 30 years ago, they now regulate the expression of over 60% of protein coding genes. [7]. Exosome is a general term for many extracellular vesicles, a type of vesicle actively secreted by the cell and encapsulated by a phospholipid bilayer in which miRNAs are encapsulated. Almost every cell or tissue in the animal body can release exosomes externally, and miRNAs can be transported to various target cells and target organs through exosomal vehicles [8][9]. An abnormal expression of miRNA is often correlated with cardiovascular, autoimmune, infectious, and neurodegenerative diseases [10][11]. MiRNAs are also involved in cancer development, acting as tumor suppressors or oncogenes [12]. Emerging studies have shown that miRNAs participate in the critical regulation of ferroptosis in cancer, but the mechanism of their regulatory role needs to be further investigated.
LncRNAs are a class of heterogeneous ncRNAs that are more than 200 nucleotides in length. They are similar to mRNAs in transcriptional and post-transcriptional mechanisms [13]. According to recent studies, LncRNAs play a crucial role in regulating cellular processes by interacting with other molecules such as DNA, RNA, and proteins. This finding sheds light on the important functions of lncRNAs in cellular processes [14][15]. H19 [16] and Xist [17] were first discovered in lncRNAs in the 1980s and 1990s. In the beginning of the 21st century, when the characteristics of ncRNAs started to exceed protein-coding genes, the role of lncRNAs began to be noticed [18]. Aberrant lncRNA expression involves all hallmarks of cancer, including sustained angiogenesis and dysregulated cellular metabolism [19][20]. In addition, there is growing evidence for their importance in regulating ferroptosis.
CircRNAs are single-stranded, covalently closed ncRNA molecules with different characteristics from other ncRNAs [21]. Initially, they were thought to be simply splicing disturbances and procedural errors produced by irregular splicing. Thus, their biological relevance was ignored. CircRNAs are rich in miRNA-binding sites and act as miRNA sponges, competing with target mRNAs for binding miRNAs, thereby inhibiting the degradation of target mRNAs [22][23]. CircRNAs play essential roles in various biological functions, such as miRNA sponges, transcriptional regulators, and RNA-binding proteins. CircRNAs are associated with developing many normal and pathological cellular processes and diseases [24], and it has been shown that they are implicated in various ferroptosis regulatory mechanisms (Figure 1).
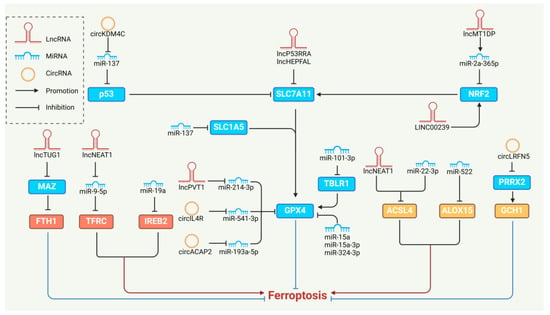
Figure 1. The regulation of ferroptosis by NcRNAs. NcRNAs target metabolizable molecules such as ACSL4 and ALOX15 in lipid metabolism to regulate ferroptosis. NcRNAs regulate ferroptosis in classical and non-classical signaling pathways, such as the p53/NRF2-SLC7A11-GPX4 axis and GCH1. NcRNAs target iron-related proteins such as FTH1 [25], TFRC [26], and IREB2 [27], and regulate ferroptosis in iron metabolism.
2. MiRNAs and Ferroptosis
The long-chain non-coding RNA lncPVT1 directly binds to miR-214-3p to inhibit its expression, whereas miR-214-3p promotes ferroptosis by targeting the degradation of GPX4 [28]. The expression of miR-101-3p was downregulated in lung cancer. MiR-101-3p promotes ferroptosis by targeting TBL1-related protein 1 (TBLR1) to downregulate GPX4 and upregulate prostaglandin-endoperoxide synthase 2 (PTGS2). By developing nanomedicines, miR-101-3p can be delivered to tumor cells in vivo for ferroptosis restoration and ultimately inhibit tumor proliferation [29]. MiR-324-3p was significantly downregulated in lung cancer cell lines compared to normal cells. MiR-324-3p induced ferroptosis and enhanced the sensitivity of cisplatin to ferroptosis via targeted GPX4 [30]. MiR-324-3p was upregulated by metformin in breast cancer cell lines and downregulated GPX4 to induce ferroptosis [31]. In colorectal cancer, miR-15a-3p promotes ferroptosis by inhibiting GPX4 and increasing the abundance of ROS, Fe2+, and MDA [32]. MiR-15a inhibited GPX4 expression in pancreatic cancer, leading to increased intracellular levels of lactate dehydrogenase, Fe2+, and ROS, thereby promoting ferroptosis. In conclusion, the induction of ferroptosis by these miRNAs through the regulation of GPX4 provides a basis for investigating therapeutic strategies for various cancers [33].
Exosomes play a crucial role in the communication between proximal and distal organs, regulating diseases through paracrine mechanisms. Cancer-associated fibroblasts inhibit ferroptosis in gastric cancer cells by targeting ALOX15 via the exosomal secretion of miR-522 and preventing lipid ROS accumulation [34]. Emerging studies in melanoma cells showed that miR-137 inhibited lipid peroxidation and iron accumulation by directly targeting solute carrier family 1 member 5 (SLC1A5). The non-Xc-system member SLC1A5 is a neutral amino acid transport protein for alanine, serine, cysteine, and glutamine [35][36]. Recent studies have shown that miR-22-3P expression is significantly upregulated in cardiomyocytes and plasma exosomes from mice with chronic myocardial infarction and patients with heart failure. The overexpression of miR-22-3p abolished erastin-induced ferroptosis in vitro. The ACSL4 is a crucial gene for fatty acid metabolism and a target gene of miR-22-3p in tumor cells. Myocardial infarction (MI) inhibits erastin-induced ferroptosis by releasing miR-22-3p-enriched exosomes derived from cardiomyocytes [37]. Thus, targeting exosome-mediated cardiomyocyte/tumor pathology communication may provide a new avenue for antitumor therapy based on ferroptosis. Previous studies have shown that iron-responsive element-binding protein 2 (IREB2) has been identified as an inducer of ferroptosis. MiR-19a represses ferroptosis by inhibiting IREB2 in colorectal cancer [27]. In summary, miRNAs can regulate ferroptosis by degrading inducers or inhibitors of ferroptosis, and exploring drugs that can target these miRNAs will be a new direction for synergistic tumor therapy.
3. LncRNAs and Ferroptosis
It was shown that lncHEPFAL expression was reduced in hepatocellular carcinoma tissues. The results indicate that lncHEPFAL promotes ferroptosis by mediating the ubiquitinated-dependent degradation of SLC7A11 and subsequently increasing lipid ROS and Fe2+ [38]. The tumor suppressor lncP53RRA is lowly expressed in lung and liver cancer [39]. Emerging studies in lung cancer have shown that lncP53RRA interacts with Ras GTPase-activated protein-binding protein 1 (G3BP1) in the cytoplasm. lncP53RRA decreased p53 binding to G3BP1 in the cytoplasm and increased the accumulation of p53 in the nucleus to promote SLC7A11 transcription and inhibit ferroptosis [40]. LncMT1DP regulates erastin-induced ferroptosis by stabilizing miR-2a-365p and inhibiting NF-E2 p45-related factor 2(NRF2) MT1DP which induces ferroptosis in non-small-cell lung cancer by increasing the abundance of ROS, MDA, and Fe2+. To enhance drug efficacy, folate (FA)-modified liposome (FA-LP) nanoparticles containing erastin and lncRNA-MT1DP (E/M@FA-LPs) increased sensitivity to erastin-induced ferroptosis by delivering erastin and MT1DP [41]. Tumor resistance or self-protective mechanisms limit the treatment of tumors, and the combination of non-coding RNAs with tumor therapy-related drugs will be an effective means to improve the therapeutic effect.
LINC00239 is an abnormally highly expressed tumor-promoting factor in colorectal cancer tissues and promotes tumor development by decreasing erastin- and RSL3-induced ferroptosis. LINC00239 inhibits NRF2 ubiquitination and increases NRF2 protein stability by interacting with the Kelch structural domain of Keap1 [42], thereby inhibiting ferroptosis. Nuclear enriched transcript 1 (NEAT1) is an oncogenic lncRNA distributed around the nucleus that affects cancer cell proliferation, cell cycle, invasion, migration, and apoptosis. NEAT1 could bind to ACSL4 mRNA, decreasing ACSL4 and inhibiting ferroptosis. NEAT1 does not significantly affect the expression of other ferroptosis factors under erastin-induced conditions, such as SLC7A11, GPX4, and TfR1, which suggests that its inhibitory effect on ferroptosis is mediated exclusively through ACSL4 [43]. In another study, lncRNAs were critical mediators in regulating iron metabolism during ferroptosis. lncNEAT1 increased cellular iron concentration, while lncRNA PR11-89 decreased cellular iron concentration to regulate ferroptosis. The former sponge miR-9-5p upregulated the expression of TFRC and GOT1, and the latter sponge miR-129-5p upregulated the expression of PROM2 [26][44]. Dihydroartemisinin (DHA) is a semi-synthetic derivative of artemisinin. Studies have shown that it has anti-glioma activity by inducing apoptosis and inhibiting the proliferation, migration, and invasion of glioma cells. Recent studies have shown that DHA can exert antitumor effects by inducing ferroptosis in glioma cells. However, the mechanisms of attenuated ferroptosis have also been demonstrated in DHA-treated glioma cells [25]. The study revealed that the downregulation of lncRNA TUG1 in DHA-treated glioma cells directly led to the upregulation of MYC-associated zinc finger protein (MAZ), which promotes FTH1 to block ferroptosis. Emerging studies suggest that lncRNAs can affect ferroptosis by targeting ferroptosis-associated transcription factors or regulators. Targeting these lncRNAs to affect ferroptosis is a potential therapeutic strategy to enhance antitumor effects.
4. CircRNAs and Ferroptosis
CircKDM4C was significantly downregulated in patients with acute myeloid leukemia (AML). CircKDM4C in AML cell lines promotes ferroptosis and inhibits cell proliferation, migration, and invasion. CircKDM4C inhibits the expression of hsa-let-7b5p as a sponge in AML cell lines, resulting in the upregulation of p53, which is the target gene of hsa-let-7b-5p.The transcription of SLC7A11 is inhibited by p53, which promotes ferroptosis [45]. CircIL4R is highly expressed in hepatocellular carcinoma and promotes tumorigenesis caused by regulating the miR-541-3p/GPX4 axis to inhibit ferroptosis [46]. CircLRFN5 expression is downregulated in glioblastoma, and the overexpression of CircLRFN5 inhibits the survival and proliferation of glioma stem cells (GSCs) as well as tumorigenesis by inducing ferroptosis [47]. CircLRFN5 binds to the transcription factor pairing-related homology box 2 (PRRX2), which promotes the degradation of PRRX2 via the ubiquitin-proteasome system and contributes to the reduction of GCH1, which is a key factor in promoting BH4 production. Targeting circLRFN5 to induce ferroptosis would be a promising therapeutic option for glioblastoma. CircRNA ACAP2 inhibits ferroptosis during cervical cancer progression via the miR-193a-5p/GPX4 axis [48]. CircACAP2 directly interacts with miR-193a-5p targeted GPX4 as a competitive RNA (ceRNA) in cervical cancer cells. Meanwhile, CircACAP2 inhibited the expression of miR-193a-5p by sponge-wrapping it, thereby promoting GPX4 expression in cervical cancer cells. CircRNAs are directly or indirectly involved in amino acid metabolism, lipid metabolism, and iron metabolism in ferroptosis. Further investigation is required to determine whether circRNAs have an exact mechanism of action in different cells and tissues, as they are a promising therapeutic target.
References
- Qu, Z.; Adelson, D.L. Evolutionary conservation and functional roles of ncRNA. Front. Genet. 2012, 3, 205.
- Kim, T.; Reitmair, A. Non-Coding RNAs: Functional Aspects and Diagnostic Utility in Oncology. Int. J. Mol. Sci. 2013, 14, 4934–4968.
- Ling, H.; Vincent, K.; Pichler, M.; Fodde, R.; Berindan-Neagoe, I.; Slack, F.J.; Calin, G.A. Junk DNA and the long non-coding RNA twist in cancer genetics. Oncogene 2015, 34, 5003–5011.
- Almeida, M.I.; Reis, R.M.; Calin, G.A. MicroRNA history: Discovery, recent applications, and next frontiers. Mutat. Res. 2011, 717, 1–8.
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402.
- Vasudevan, S.; Tong, Y.; Steitz, J.A. Switching from repression to activation: MicroRNAs can up-regulate translation. Science 2007, 318, 1931–1934.
- Zhang, F.; Wang, D. The Pattern of microRNA Binding Site Distribution. Genes 2017, 8, 296.
- Zhang, Y.; Liu, D.; Chen, X.; Li, J.; Li, L.; Bian, Z.; Sun, F.; Lu, J.; Yin, Y.; Cai, X.; et al. Secreted monocytic miR-150 enhances targeted endothelial cell migration. Mol. Cell 2010, 39, 133–144.
- Yin, Y.; Cai, X.; Chen, X.; Liang, H.; Zhang, Y.; Li, J.; Wang, Z.; Chen, X.; Zhang, W.; Yokoyama, S.; et al. Tumor-secreted miR-214 induces regulatory T cells: A major link between immune evasion and tumor growth. Cell Res. 2014, 24, 1164–1180.
- Santamaria, X.; Taylor, H. MicroRNA and gynecological reproductive diseases. Fertil. Steril. 2014, 101, 1545–1551.
- Nguyen, M.T.; Lee, W. Role of MiR-325-3p in the Regulation of CFL2 and Myogenic Differentiation of C2C12 Myoblasts. Cells 2021, 10, 2725.
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222.
- Xing, J.; Liu, H.; Jiang, W.; Wang, L. LncRNA-Encoded Peptide: Functions and Predicting Methods. Front. Oncol. 2020, 10, 622294.
- Jonas, K.; Calin, G.A.; Pichler, M. RNA-Binding Proteins as Important Regulators of Long Non-Coding RNAs in Cancer. Int. J. Mol. Sci. 2020, 21, 2969.
- Bartonicek, N.; Maag, J.L.; Dinger, M.E. Long noncoding RNAs in cancer: Mechanisms of action and technological advancements. Mol. Cancer 2016, 15, 43.
- Bartolomei, M.S.; Zemel, S.; Tilghman, S.M. Parental imprinting of the mouse H19 gene. Nature 1991, 351, 153–155.
- Brown, C.J.; Ballabio, A.; Rupert, J.L.; Lafreniere, R.G.; Grompe, M.; Tonlorenzi, R.; Willard, H.F. A gene from the region of the human X inactivation centre is expressed exclusively from the inactive X chromosome. Nature 1991, 349, 38–44.
- Prensner, J.R.; Chinnaiyan, A.M. The emergence of lncRNAs in cancer biology. Cancer Discov. 2011, 1, 391–407.
- Teppan, J.; Barth, D.A.; Prinz, F.; Jonas, K.; Pichler, M.; Klec, C. Involvement of Long Non-Coding RNAs (lncRNAs) in Tumor Angiogenesis. Non-Coding RNA 2020, 6, 42.
- Balihodzic, A.; Barth, D.A.; Prinz, F.; Pichler, M. Involvement of Long Non-Coding RNAs in Glucose Metabolism in Cancer. Cancers 2021, 13, 977.
- Ashekyan, O.; Abdallah, S.; Shoukari, A.A.; Chamandi, G.; Choubassy, H.; Itani, A.R.S.; Alwan, N.; Nasr, R. Spotlight on Exosomal Non-Coding RNAs in Breast Cancer: An In Silico Analysis to Identify Potential lncRNA/circRNA-miRNA-Target Axis. Int. J. Mol. Sci. 2022, 23, 8351.
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388.
- Liu, C.X.; Chen, L.L. Circular RNAs: Characterization, cellular roles, and applications. Cell 2022, 185, 2016–2034.
- Verduci, L.; Tarcitano, E.; Strano, S.; Yarden, Y.; Blandino, G. CircRNAs: Role in human diseases and potential use as biomarkers. Cell Death Dis. 2021, 12, 468.
- Gong, H.; Gao, M.; Lin, Y.; Liu, J.; Hu, Z.; Liu, J. TUG1/MAZ/FTH1 Axis Attenuates the Antiglioma Effect of Dihydroartemisinin by Inhibiting Ferroptosis. Oxidative Med. Cell. Longev. 2022, 2022, 7843863.
- Wei, X.B.; Jiang, W.Q.; Zeng, J.H.; Huang, L.Q.; Ding, H.G.; Jing, Y.W.; Han, Y.L.; Li, Y.C.; Chen, S.L. Exosome-Derived lncRNA NEAT1 Exacerbates Sepsis-Associated Encephalopathy by Promoting Ferroptosis Through Regulating miR-9-5p/TFRC and GOT1 Axis. Mol. Neurobiol. 2022, 59, 1954–1969.
- Fan, H.; Ai, R.; Mu, S.; Niu, X.; Guo, Z.; Liu, L. MiR-19a suppresses ferroptosis of colorectal cancer cells by targeting IREB2. Bioengineered 2022, 13, 12021–12029.
- He, G.N.; Bao, N.R.; Wang, S.; Xi, M.; Zhang, T.H.; Chen, F.S. Ketamine Induces Ferroptosis of Liver Cancer Cells by Targeting lncRNA PVT1/miR-214-3p/GPX4. Drug Des. Dev. Ther. 2021, 15, 3965–3978.
- Luo, Y.; Niu, G.; Yi, H.; Li, Q.; Wu, Z.; Wang, J.; Yang, J.; Li, B.; Peng, Y.; Liang, Y.; et al. Nanomedicine promotes ferroptosis to inhibit tumour proliferation in vivo. Redox Biol. 2021, 42, 101908.
- Deng, S.H.; Wu, D.M.; Li, L.; Liu, T.; Zhang, T.; Li, J.; Yu, Y.; He, M.; Zhao, Y.Y.; Han, R.; et al. miR-324-3p reverses cisplatin resistance by inducing GPX4-mediated ferroptosis in lung adenocarcinoma cell line A549. Biochem. Biophys. Res. Commun. 2021, 549, 54–60.
- Hou, Y.; Cai, S.; Yu, S.; Lin, H. Metformin induces ferroptosis by targeting miR-324-3p/GPX4 axis in breast cancer. Acta Biochim. Et. Biophys. Sin. 2021, 53, 333–341.
- Liu, L.; Yao, H.; Zhou, X.; Chen, J.; Chen, G.; Shi, X.; Wu, G.; Zhou, G.; He, S. MiR-15a-3p regulates ferroptosis via targeting glutathione peroxidase GPX4 in colorectal cancer. Mol. Carcinog. 2022, 61, 301–310.
- Xu, P.; Wang, Y.; Deng, Z.; Tan, Z.; Pei, X. MicroRNA-15a promotes prostate cancer cell ferroptosis by inhibiting GPX4 expression. Oncol. Lett. 2022, 23, 67.
- Zhang, H.; Deng, T.; Liu, R.; Ning, T.; Yang, H.; Liu, D.; Zhang, Q.; Lin, D.; Ge, S.; Bai, M.; et al. CAF secreted miR-522 suppresses ferroptosis and promotes acquired chemo-resistance in gastric cancer. Mol. Cancer 2020, 19, 43.
- Scalise, M.; Pochini, L.; Console, L.; Losso, M.A.; Indiveri, C. The Human SLC1A5 (ASCT2) Amino Acid Transporter: From Function to Structure and Role in Cell Biology. Front. Cell Dev. Biol. 2018, 6, 96.
- Luo, M.; Wu, L.; Zhang, K.; Wang, H.; Zhang, T.; Gutierrez, L.; O’Connell, D.; Zhang, P.; Li, Y.; Gao, T.; et al. miR-137 regulates ferroptosis by targeting glutamine transporter SLC1A5 in melanoma. Cell Death Differ. 2018, 25, 1457–1472.
- Yuan, Y.; Mei, Z.; Qu, Z.; Li, G.; Yu, S.; Liu, Y.; Liu, K.; Shen, Z.; Pu, J.; Wang, Y.; et al. Exosomes secreted from cardiomyocytes suppress the sensitivity of tumor ferroptosis in ischemic heart failure. Signal Transduct. Target. Ther. 2023, 8, 121.
- Zhang, B.; Bao, W.; Zhang, S.; Chen, B.; Zhou, X.; Zhao, J.; Shi, Z.; Zhang, T.; Chen, Z.; Wang, L.; et al. LncRNA HEPFAL accelerates ferroptosis in hepatocellular carcinoma by regulating SLC7A11 ubiquitination. Cell Death Dis. 2022, 13, 734.
- Chen, C.; Zheng, Q.; Kang, W.; Yu, C. Long non-coding RNA LINC00472 suppresses hepatocellular carcinoma cell proliferation, migration and invasion through miR-93-5p/PDCD4 pathway. Clin. Res. Hepatol. Gastroenterol. 2019, 43, 436–445.
- Koppula, P.; Zhuang, L.; Gan, B. Cystine transporter SLC7A11/xCT in cancer: Ferroptosis, nutrient dependency, and cancer therapy. Protein Cell 2021, 12, 599–620.
- Gai, C.; Liu, C.; Wu, X.; Yu, M.; Zheng, J.; Zhang, W.; Lv, S.; Li, W. MT1DP loaded by folate-modified liposomes sensitizes erastin-induced ferroptosis via regulating miR-365a-3p/NRF2 axis in non-small cell lung cancer cells. Cell Death Dis. 2020, 11, 751.
- Han, Y.; Gao, X.; Wu, N.; Jin, Y.; Zhou, H.; Wang, W.; Liu, H.; Chu, Y.; Cao, J.; Jiang, M.; et al. Long noncoding RNA LINC00239 inhibits ferroptosis in colorectal cancer by binding to Keap1 to stabilize Nrf2. Cell Death Dis. 2022, 13, 742.
- Wu, H.; Liu, A. Long non-coding RNA NEAT1 regulates ferroptosis sensitivity in non-small-cell lung cancer. J. Int. Med. Res. 2021, 49, 300060521996183.
- Luo, W.; Wang, J.; Xu, W.; Ma, C.; Wan, F.; Huang, Y.; Yao, M.; Zhang, H.; Qu, Y.; Ye, D.; et al. LncRNA RP11-89 facilitates tumorigenesis and ferroptosis resistance through PROM2-activated iron export by sponging miR-129-5p in bladder cancer. Cell Death Dis. 2021, 12, 1043.
- Dong, L.H.; Huang, J.J.; Zu, P.; Liu, J.; Gao, X.; Du, J.W.; Li, Y.F. CircKDM4C upregulates P53 by sponging hsa-let-7b-5p to induce ferroptosis in acute myeloid leukemia. Environ. Toxicol. 2021, 36, 1288–1302.
- Xu, Q.; Zhou, L.; Yang, G.; Meng, F.; Wan, Y.; Wang, L.; Zhang, L. CircIL4R facilitates the tumorigenesis and inhibits ferroptosis in hepatocellular carcinoma by regulating the miR-541-3p/GPX4 axis. Cell Biol. Int. 2020, 44, 2344–2356.
- Jiang, Y.; Zhao, J.; Li, R.; Liu, Y.; Zhou, L.; Wang, C.; Lv, C.; Gao, L.; Cui, D. CircLRFN5 inhibits the progression of glioblastoma via PRRX2/GCH1 mediated ferroptosis. J. Exp. Clin. Cancer Res. 2022, 41, 307.
- Liu, Y.; Li, L.; Yang, Z.; Wen, D.; Hu, Z. Circular RNA circACAP2 Suppresses Ferroptosis of Cervical Cancer during Malignant Progression by miR-193a-5p/GPX4. J. Oncol. 2022, 2022, 5228874.
More
Information
Subjects:
Cell Biology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
355
Revisions:
2 times
(View History)
Update Date:
12 Mar 2024
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No