Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Gábor Nagy-Grócz | -- | 2566 | 2024-03-04 09:16:10 | | | |
| 2 | Fanny Huang | Meta information modification | 2566 | 2024-03-05 08:28:27 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Nagy-Grócz, G.; Spekker, E.; Vécsei, L. Connections between Intestinal Flora and Kynurenine Pathway. Encyclopedia. Available online: https://encyclopedia.pub/entry/55804 (accessed on 24 June 2026).
Nagy-Grócz G, Spekker E, Vécsei L. Connections between Intestinal Flora and Kynurenine Pathway. Encyclopedia. Available at: https://encyclopedia.pub/entry/55804. Accessed June 24, 2026.
Nagy-Grócz, Gábor, Eleonóra Spekker, László Vécsei. "Connections between Intestinal Flora and Kynurenine Pathway" Encyclopedia, https://encyclopedia.pub/entry/55804 (accessed June 24, 2026).
Nagy-Grócz, G., Spekker, E., & Vécsei, L. (2024, March 04). Connections between Intestinal Flora and Kynurenine Pathway. In Encyclopedia. https://encyclopedia.pub/entry/55804
Nagy-Grócz, Gábor, et al. "Connections between Intestinal Flora and Kynurenine Pathway." Encyclopedia. Web. 04 March, 2024.
Copy Citation
The intestinal flora has been the focus of numerous investigations recently, with inquiries not just into the gastrointestinal aspects but also the pathomechanism of other diseases such as nervous system disorders and mitochondrial diseases. Mitochondrial disorders are the most common type of inheritable metabolic illness caused by mutations of mitochondrial and nuclear DNA. Metabolites of the kynurenine pathway are linked to many disorders, such as depression, schizophrenia, migraine, and also diseases associated with impaired mitochondrial function. The kynurenine pathway includes many substances, for instance kynurenic acid and quinolinic acid.
kynurenine pathway
intestinal flora
1. Introduction
The gut microbiota, also referred to as the intestinal flora, has become a focal point in healthcare. One of the most noteworthy advancements in recent gut microbiota research has been the revelation that the assembly of microorganisms in our digestive system has the capacity to influence various aspects of brain development and function.
Examining the neurobiological mechanisms responsible for the degree of influence exerted by microbial organisms on host physiology, brain function, and behavior has become a top research priority. Researchers are currently exploring various pathways and potential mechanisms that may govern the interactions between intestinal flora and the brain.
One central focus in this area of investigation is how the microbiota regulates the availability of circulating tryptophan (Trp). Trp is an essential amino acid that is responsible for the synthesis of serotonin (5-HT) and, moreover, is an initial compound of the kynurenine pathway (KP). The KP has a crucial role in many disorders, such as immunological [1], neurological [2], and mitochondrial diseases [3]. Mitochondrial disorders are chronic genetic illnesses that occur when mitochondria in cells fail to produce sufficient energy due to damage to their own DNA.
There is limited direct evidence establishing a clear and causal connection between the gut microbiota and mitochondrial disorders. However, research in both areas has been progressing, and there is growing interest in understanding potential links between them.
2. Connections between the Intestinal Flora and the Kynurenine Pathway
2.1. Intestinal Flora
Intestinal flora, also known as the gut microbiota or gut microbiome, refers to the diverse community of microorganisms that inhabit the human gastrointestinal tract, primarily the colon. These microorganisms consist of bacteria, viruses, fungi, and other microorganisms, with bacteria being the most abundant and well-studied component. The human gut is home to trillions of microorganisms, with an estimated 1000 to 1150 different species of bacteria.
The initial colonization of microbes primarily occurs during the process of childbirth. Newborns delivered through the vaginal route are exposed to bacteria from their mother’s feces and vaginal region, while infants born via cesarean section initially encounter bacteria from the hospital environment and their mother’s skin [4]. It is important to note that despite the long-lasting belief that the prenatal environment is completely sterile, recent research has shown that before breastfeeding, the placenta, the amniotic fluid, and meconium can carry a certain amount of bacteria [5]. Studies have detected, in the infant gut, the presence of bacteria such as Enterobacteriaceae, Bifidobacterium and Bacteroides [6].
The infant gut microbiota remains highly changeable and dynamic until around the age of two when the consumption of solid foods begins [4]. Afterward, around the third year of life, as the diet becomes more diverse, the microbiota stabilizes and starts resembling the microbial compositions found in adults [5]. In adulthood, healthy people’s gut microbiota is primarily populated by four main phyla: Firmicutes, Actinobacteria, Bacteroidetes, and Verrucomicrobia [7]. The composition of the gut microbiota in healthy young adults and middle-aged individuals is characterized by a high diversity of bacterial species [8]. However, as individuals age, their gut microbiota undergoes changes, with a higher proportion of Bacteroides spp. and distinct patterns of Clostridium groups identified in the elderly compared to younger adults [9]. Bacteria in these species can also metabolize Trp into substances of the KP [10].
Therefore, during infancy and old age, gut microbiota composition is highly dynamic and experiences significant shifts, whereas in healthy young adults and middle-aged individuals, it tends to be more stable and diverse. Even during adulthood, the composition of the gut microbiota can undergo significant changes over the course of a year [11]. This has led to debates regarding the best way to characterize and monitor an individual’s gut microbiota composition. The concept of “enterotypes” (three core clusters of bacterial genera: Bacteroides, Prevotella, and Ruminococcus) is not universally accepted due to variations between individuals and challenges in categorizing an individual’s gut microbial composition within one specific enterotype [11]. An alternative approach suggests that the gut microbiota composition reflects a core set of functional profiles, where certain bacterial species play a more critical role in influencing health and disease [12].
This diversity plays a crucial role in maintaining a healthy gut. So gut microorganisms are an intricate microbial community recognized as a crucial influencer of neurodevelopment [13], and their involvement in the development of excitotoxicity is highly probable [14]. The potential contribution of gut microbiota to brain development can be described as follows [15]. On the one hand, Trp serves as a precursor for various biologically significant metabolites with crucial physiological roles, and it holds significant implications for the development of the central nervous system (CNS). Serotonin (5-hydroxytryptamine, 5-HT), originating from Trp via the action of Trp hydroxylase, regulates essential neurodevelopmental approaches [15]. These processes are under the influence of short-chain fatty acids (SCFAs) produced by the gut microbiome, such as acetate, propionate, and butyrate [16][17]. These substances can influence the brain by modifying the levels of neurotransmitter precursors [18] and via the vagal nerve [19]. On the other hand, neurotrophins such as brain-derived neurotrophic factor (BDNF), which are closely associated with neurodevelopment and neuroprotection, play crucial roles in promoting neuronal survival, synaptic plasticity, and cognitive function [15]. Recently, it has been also shown that the KP is changed in BDNF knock-in mice [20], so there is a connection between the substances of the KP and the BDNF pathway. Additionally, it is proposed that gut microbiota may influence the expression of N-methyl-D-aspartate (NMDA) receptors and contribute to brain development [21].
To sum up, bacteria in the gut microbiome can metabolize Trp, leading to the production of various metabolites within the gut (Figure 1).
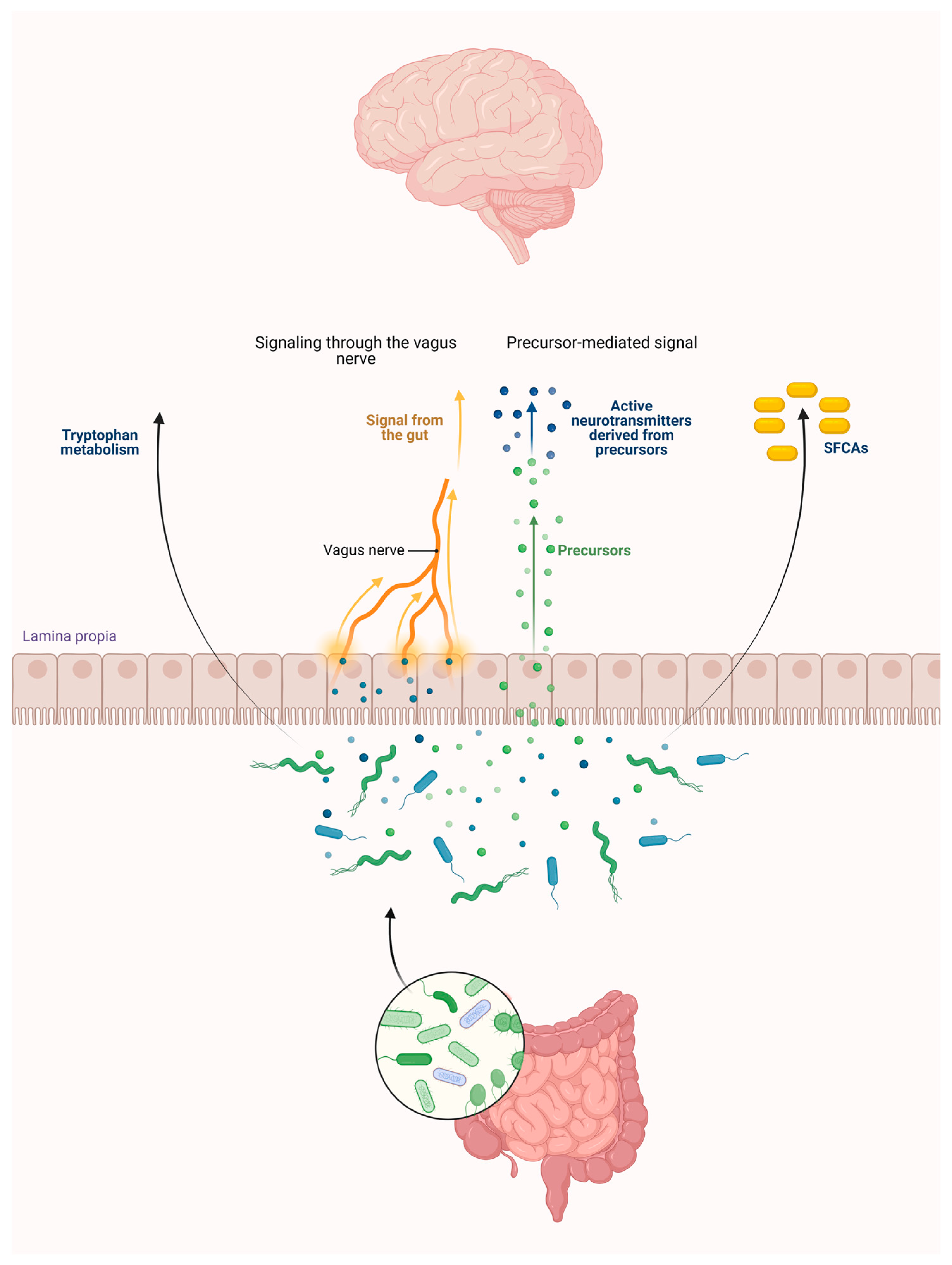
Figure 1. The gut microbiota influences the brain through tryptophan, short-chain fatty acids via the vagus nerve, and the precursor of neurotransmitters. Abbreviation: SCFAs—short-chain fatty acids.
2.2. The KP and Its Receptors
Trp is an essential amino acid that is crucial in the brain as it is the precursor of 5-HT. Several bacterial strains have the capability to enhance the production of Trp in the body, potentially playing a role in maintaining regular gut motility [22]. Nonetheless, the majority (more than 90%) of Trp in mammalian cells metabolizes in the KP and not towards the 5-HT (Figure 2). Kynurenic acid (KYNA) is one of the best-known substances of the KP, which was first described by Justus von Liebig in 1853 [23]. Liebig was a German scientist, who discovered KYNA in dog urine. It is important to know that KYNA is an endogenous glutamate receptor antagonist and has neuroprotective effects. This substance is created by kynurenine aminotransferases (KATs) from L-kynurenine (L-KYN), which is constructed by the formamidase enzyme from N-formylkynurenine. N-formylkynurenine, and is formed from L-Trp by two iron-dependent enzymes: indolamine 2,3-dioxygenase 1 and 2 (IDO1 and IDO2) and tryptophan 2,3-dioxygenase (TDO). It is noteworthy that, besides KYNA, L-KYN can transform into anthranilic acid (ANA) in reaction to kynureninase (3-HAO) and to 3-hydroxykynurenine (3-HK) when reacting to kynurenine 3-monooxygenase (KMO). KMO, in eukaryotic cells, is a mitochondrial protein located in the outer membrane of mitochondria [24]. ANA can further transform into 3-hydroxyanthranilic acid (3-HAA) when exposed to 3-hydroxyanthranilic acid hydroxylase. Furthermore, 3-HK can also be converted to 3-HAA by the kynureninase enzyme. In addition to this, 3-HK can also transform into xanthurenic acid. 3-HAA can further be converted to quinolinic acid (QUIN) when exposed to 3-hydroxyanthranillic acid 3,4-dioxygenase. In the last step of the KP, QUIN is transformed into nicotinamide adenine dinucleotide (NAD+) in reaction to quinolinic acid phosphoribosyl transferase. NAD+ has a pivotal role in mitochondrial energy management and redox reactions [25]. Different from KYNA, QUIN is an endogenous glutamate receptor agonist, which when produced by microglia [26], can provoke lipid peroxidation [27] and has a relevant role in the neurodegenerative process [28]. QUIN can exert its effects, probably by stimulating NMDA receptors, yielding the overproduction of free radicals and inhibiting the respiratory chain. Research shows that the enzymes and substances of the KP have pivotal roles in the pathomechanisms of migraine [29], Parkinson’s disease [30], schizophrenia [31] and mitochondrial dysfunction [32][33][34].
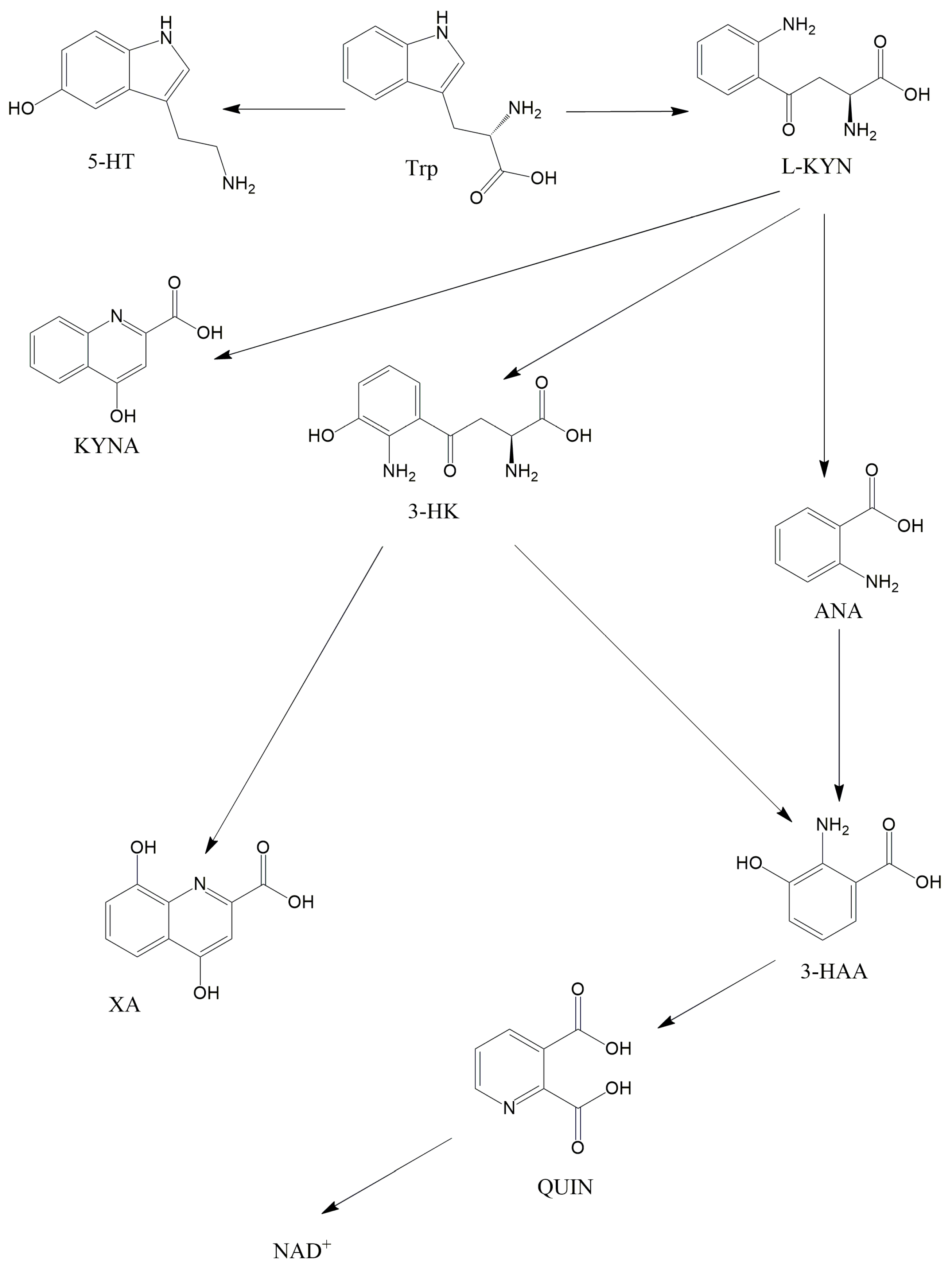
Figure 2. The kynurenine pathway. This figure illustrates the main metabolites of the KP. Abbreviations: 3-HAA—3-hydroxyanthranilic acid, 3-HK—3-hydroxykynurenine, 5-HT—serotonin, ANA—anthranilic acid, KYNA—kynurenic acid, L-KYN—L-kynurenine, NAD+—nicotinamide adenine dinucleotide, QUIN—quinolinic acid, Trp—tryptophan, XA—xanthurenic acid.
Kynurenines exert their effects on different receptors, such as glutamate receptors in a dose dependent manner, aryl hydrocarbon (AhR) receptors, G protein-coupled receptor 35 (GPR35), and α-7 nicotinic (α7 nACh) receptors (Figure 3). Glutamate receptors are a class of neurotransmitter receptors found in the CNS of animals, hence humans. Glutamate is the most abundant excitatory neurotransmitter in the brain, and its receptors play a fundamental role in various aspects of neuronal communication and synaptic plasticity. There are several types of glutamate receptors, but they are generally categorized into two major classes. The ionotropic glutamate receptors function as ligand-gated ion channels. This implies that upon binding with glutamate, they facilitate the movement of ions, including the influx of sodium (Na+) and calcium (Ca2+) as well as the efflux of potassium (K+) through the cell membrane. Ionotropic receptors mediate fast synaptic transmission and include three subtypes: NMDA receptors, α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors, and kainate receptors. The other types of glutamate receptors are metabotropic glutamate receptors, which are G protein-coupled receptors that are indirectly linked to ion channels through intracellular signaling pathways. They modulate synaptic transmission and play a role in the regulation of neuronal excitability. KYNA is an antagonist at the strychnine-insensitive glycine-binding site of NMDA receptors at low doses [35] and can also inhibit the glutamate-binding site of the NMDA receptors at higher doses [36]. In addition to this, KYNA has an antagonistic impact on kainate- and AMPA receptors [35]. The effect of KYNA on AMPA receptors is also concentration dependent, which means that KYNA can stimulate the receptors at nanomolar and micromolar concentrations but in different circumstances, namely between micromolar and millimolar concentrations, can inhibit the AMPA receptors [37][38].
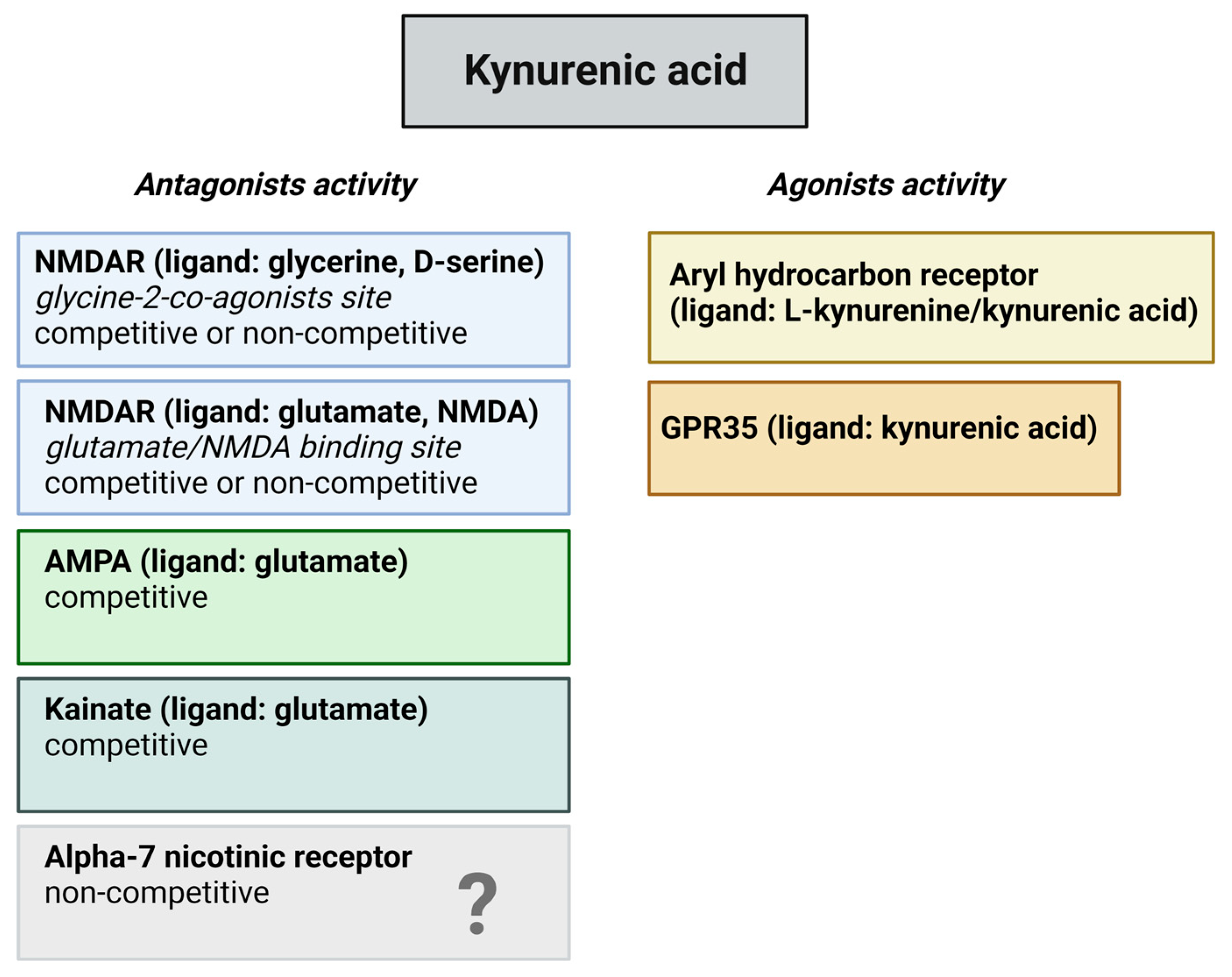
Figure 3. Potential binding sites for kynurenic acid. The figure provides an overview of potential receptors for kynurenic acid and their corresponding actions. Recent inquiries have raised uncertainties regarding its impact on the alpha-7 nicotinic receptor. Abbreviations: NMDAR—N-methyl-D-aspartate receptors, AMPA—α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptors, GPR35—G protein-coupled receptor 35.
AhR receptors are a family of proteins found in many species, including humans. AhR is a ligand-activated transcription factor that can bind to various aromatic hydrocarbons, including environmental pollutants like dioxins and polycyclic aromatic hydrocarbons (PAHs). When AhR binds to its ligands, it undergoes a conformational change and translocates to the cell nucleus, where it regulates the expression of various genes. The AhR receptor activation promotes equilibrium in host-microbe interactions, with indole derived from microbial Trp serving as a crucial ligand for this transcription factor [39]. Additionally, the AhR receptor is involved in the regulation of IDO and TDO expression [40][41]. Elevated AhR receptor activation has been noted in individuals with post-acute sequelae of SARS-CoV-2 infection, correlating with heightened IDO2 activity [42]. Additionally, a decline in mitochondrial function is evident in these patients. Experimental findings indicate that AhR receptor antagonists can diminish IDO2 activation [42]. Consequently, it can be inferred that modifications in the KP might impact mitochondrial function through AhR receptors.
GPR35 is a member of the G protein-coupled receptor family. The exact function of GPR35 is not fully understood, and it appears to have diverse roles in different tissues. Research has implicated GPR35 in immune responses, inflammatory processes, and gastrointestinal functions.
The α7 nACh receptor is a type of nicotinic acetylcholine receptor that plays a crucial role in neurotransmission within the CNS. Nicotinic receptors are ionotropic receptors that respond to the neurotransmitter acetylcholine, and they are named after nicotine, which also binds to and activates these receptors. Doubts regarding the effects of KYNA on α7 nACh receptors have been raised by Stone [43].
2.3. Role of the Intestinal Flora in the KP Metabolism
The relationship between diet, the KP, and the gut microbiome is a complex and emerging area of research in the fields of nutrition, metabolism, and microbiology. Various diets can significantly influence the composition and function of the gut microbiome [44]. For example, diets rich in fiber from fruits, vegetables, and whole grains can promote the growth of beneficial bacteria in the gut [45]. On the other hand, high-fat and high-sugar diets can alter the microbiome composition and may contribute to the development of metabolic disorders [46].
Research has shown that Trp, and thus the KP, has a unique role in the dual communication between microorganisms of the gut and the various organs of the host [47]. Bacteria in the gut can influence the activity of IDO1 in the gut, and some of these bacteria have enzymes very similar to those of the KP and thus contribute to the production of the metabolites of the KP [47]. It is widely recognized that the bidirectional communication between the microbiota and the immune system plays a crucial role in shaping the host’s intestinal immune response [48]. Consequently, mice that lack gut microbiota display a deficiency in their innate immune system. When compared to conventional mice, germ-free mice exhibit decreased degradation of Trp via the KP, leading to higher levels of available Trp and lower levels of KYNA in their bloodstream [49][50]. Additionally, the levels of circulating Trp and KYNA return to normal after microbial colonization in mice immediately after weaning [50].
In a recent study, Sun and his colleagues found that long-term high-fat diets disrupted the metabolism of Trp in the bloodstream in mice [51]. This disruption was characterized by a reduction in Trp levels and an increase in the activity of the KP. Notably, this aberrant Trp metabolism strongly correlated with the proliferation of the Proteobacteria phylum in the colon, so deviations in Trp metabolism can induce changes in the KP metabolism [51] as well. In this interesting research, the authors could abrogate the long-term high-fat diet shift with antibiotic treatment, but changes induced by the long-term high-fat diet could be transferred to mice with a standard diet through fecal transplantation, thus revealing the role of gut microbiota in this process. In another experimental setting, it was shown that the administration of Bacillus infantis to rats elevates Trp and KYNA levels but decreases the level of 5-Hydroxyindoleacetic acid [52], which is the main metabolite of 5-HT [53]. These data obviously prove that the gut microbiome influences the KP.
Alterations in the KP have been documented in several diseases linked to a disrupted microbiome. Patients diagnosed with irritable bowel syndrome (IBS) have reported heightened KP metabolism and altered microbiome [54][55][56], providing evidence of a connection between these two systems. The roles of altered microbiome and oxidative stress have also been described in patients with Crohn’s disease [57][58]. In a recent study, it has been shown that Trp has a protective impact on Crohn’s disease and IBS [59], by strengthening the role of the KP against mitochondrial stress and supporting a normal microbiome.
However, it is important to note that the interplay between diet, kynurenine, and the gut microbiome is still an active area of research and that the mechanisms involved are not fully understood. It is also important to consider individual variations in microbiome composition and responses to diet.
References
- Pires, A.S.; Sundaram, G.; Heng, B.; Krishnamurthy, S.; Brew, B.J.; Guillemin, G.J. Recent advances in clinical trials targeting the kynurenine pathway. Pharmacol. Ther. 2022, 236, 108055.
- Huang, Y.S.; Ogbechi, J.; Clanchy, F.I.; Williams, R.O.; Stone, T.W. IDO and Kynurenine Metabolites in Peripheral and CNS Disorders. Front. Immunol. 2020, 11, 388.
- Sas, K.; Szabó, E.; Vécsei, L. Mitochondria, Oxidative Stress and the Kynurenine System, with a Focus on Ageing and Neuroprotection. Molecules 2018, 23, 191.
- Borre, Y.E.; O’Keeffe, G.W.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Microbiota and neurodevelopmental windows: Implications for brain disorders. Trends Mol. Med. 2014, 20, 509–518.
- Rodríguez, J.M.; Murphy, K.; Stanton, C.; Ross, R.P.; Kober, O.I.; Juge, N.; Avershina, E.; Rudi, K.; Narbad, A.; Jenmalm, M.C.; et al. The composition of the gut microbiota throughout life, with an emphasis on early life. Microb. Ecol. Health Dis. 2015, 26, 26050.
- Adlerberth, I.; Wold, A.E. Establishment of the gut microbiota in Western infants. Acta Paediatr. 2009, 98, 229–238.
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14.
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230.
- Kennedy, P.J.; Cryan, J.F.; Dinan, T.G.; Clarke, G. Kynurenine pathway metabolism and the microbiota-gut-brain axis. Neuropharmacology 2017, 112 Pt B, 399–412.
- Agus, A.; Planchais, J.; Sokol, H. Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease. Cell Host Microbe 2018, 23, 716–724.
- Knights, D.; Ward, T.L.; McKinlay, C.E.; Miller, H.; Gonzalez, A.; McDonald, D.; Knight, R. Rethinking “enterotypes”. Cell Host Microbe 2014, 16, 433–437.
- Flint, H.J.; Scott, K.P.; Louis, P.; Duncan, S.H. The role of the gut microbiota in nutrition and health. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 577–589.
- Sharon, G.; Sampson, T.R.; Geschwind, D.H.; Mazmanian, S.K. The Central Nervous System and the Gut Microbiome. Cell 2016, 167, 915–932.
- Sharma, S.; Awasthi, A.; Singh, S. Altered gut microbiota and intestinal permeability in Parkinson’s disease: Pathological highlight to management. Neurosci. Lett. 2019, 712, 134516.
- Wang, Y.; Yuan, X.; Kang, Y.; Song, X. Tryptophan-kynurenine pathway as a novel link between gut microbiota and schizophrenia: A review. Trop. J. Pharm. Res. 2019, 18, 897–905.
- Reigstad, C.S.; Salmonson, C.E.; Rainey, J.F.; Szurszewski, J.H.; Linden, D.R.; Sonnenburg, J.L.; Farrugia, G.; Kashyap, P.C. Gut microbes promote colonic serotonin production through an effect of short-chain fatty acids on enterochromaffin cells. FASEB J. 2015, 29, 1395–1403.
- Anand, N.; Gorantla, V.R.; Chidambaram, S.B. The Role of Gut Dysbiosis in the Pathophysiology of Neuropsychiatric Disorders. Cells 2022, 12, 54.
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The role of short-chain fatty acids in health and disease. Adv. Immunol. 2014, 121, 91–119.
- de Lau, L.M.; Bornebroek, M.; Witteman, J.C.; Hofman, A.; Koudstaal, P.J.; Breteler, M.M. Dietary fatty acids and the risk of Parkinson disease: The Rotterdam study. Neurology 2005, 64, 2040–2045.
- Ieraci, A.; Beggiato, S.; Ferraro, L.; Barbieri, S.S.; Popoli, M. Kynurenine pathway is altered in BDNF Val66Met knock-in mice: Effect of physical exercise. Brain Behav. Immun. 2020, 89, 440–450.
- Maqsood, R.; Stone, T.W. The Gut-Brain Axis, BDNF, NMDA and CNS Disorders. Neurochem. Res. 2016, 41, 2819–2835.
- Legan, T.B.; Lavoie, B.; Norberg, E.; Ley, I.C.; Tack, S.; Tompkins, T.A.; Wargo, M.J.; Mawe, G.M. Tryptophan-synthesizing bacteria enhance colonic motility. Neurogastroenterol. Motil. 2023, 35, e14629.
- Liebig, J. Uber Kynurensäure. Ann. Chem. 1853, 86, 125–126.
- Hirai, K.; Kuroyanagi, H.; Tatebayashi, Y.; Hayashi, Y.; Hirabayashi-Takahashi, K.; Saito, K.; Haga, S.; Uemura, T.; Izumi, S. Dual role of the carboxyl-terminal region of pig liver l-kynurenine 3-monooxygenase: Mitochondrial-targeting signal and enzymatic activity. J. Biochem. 2010, 148, 639–650.
- Castro-Portuguez, R.; Sutphin, G.L. Kynurenine pathway, NAD+ synthesis, and mitochondrial function: Targeting tryptophan metabolism to promote longevity and healthspan. Exp. Gerontol. 2020, 132, 110841.
- Guillemin, G.J.; Smythe, G.A.; Veas, L.A.; Takikawa, O.; Brew, B.J. A beta 1-42 induces production of quinolinic acid by human macrophages and microglia. Neuroreport 2003, 14, 2311–2315.
- Behan, W.M.H.; McDonald, M.; Darlington, L.G.; Stone, T.W. Oxidative stress as a mechanism for quinolinic acid-induced hippocampal damage: Protection by melatonin and deprenyl. Br. J. Pharmacol. 1999, 128, 1754–1760.
- Lugo-Huitrón, R.; Ugalde Muñiz, P.; Pineda, B.; Pedraza-Chaverrí, J.; Ríos, C.; Pérez-de la Cruz, V. Quinolinic acid: An endogenous neurotoxin with multiple targets. Oxid. Med. Cell Longev. 2013, 2013, 104024.
- Nagy-Grócz, G.; Zádor, F.; Dvorácskó, S.; Bohár, Z.; Benyhe, S.; Tömböly, C.; Párdutz, Á.; Vécsei, L. Interactions between the kynurenine and the endocannabinoid system with special emphasis on migraine. Int. J. Mol. Sci. 2017, 18, 1617.
- Lim, C.K.; Fernández-Gomez, F.J.; Braidy, N.; Estrada, C.; Costa, C.; Costa, S.; Bessede, A.; Fernandez-Villalba, E.; Zinger, A.; Herrero, M.T.; et al. Involvement of the kynurenine pathway in the pathogenesis of Parkinson’s disease. Prog. Neurobiol. 2017, 155, 76–95.
- Zádor, F.; Nagy-Grócz, G.; Kekesi, G.; Dvorácskó, S.; Szűcs, E.; Tömböly, C.; Horvath, G.; Benyhe, S.; Vécsei, L. Kynurenines and the Endocannabinoid System in Schizophrenia: Common Points and Potential Interactions. Molecules 2019, 24, 3709.
- González Esquivel, D.; Ramírez-Ortega, D.; Pineda, B.; Castro, N.; Ríos, C.; Pérez de la Cruz, V. Kynurenine pathway metabolites and enzymes involved in redox reactions. Neuropharmacology 2017, 112 Pt B, 331–345.
- Baran, H.; Staniek, K.; Bertignol-Spörr, M.; Attam, M.; Kronsteiner, C.; Kepplinger, B. Effects of Various Kynurenine Metabolites on Respiratory Parameters of Rat Brain, Liver and Heart Mitochondria. Int. J. Tryptophan Res. 2016, 9, 17–29.
- Maddison, D.C.; Alfonso-Núñez, M.; Swaih, A.M.; Breda, C.; Campesan, S.; Allcock, N.; Straatman-Iwanowska, A.; Kyriacou, C.P.; Giorgini, F. A novel role for kynurenine 3-monooxygenase in mitochondrial dynamics. PLoS Genet. 2020, 16, e1009129.
- Birch, P.J.; Grossman, C.J.; Hayes, A.G. Kynurenic acid antagonises responses to NMDA via an action at the strychnine-insensitive glycine receptor. Eur. J. Pharmacol. 1988, 154, 85–87.
- Kessler, M.; Terramani, T.; Lynch, G.; Baudry, M. A Glycine Site Associated with N-Methyl-d-Aspartic Acid Receptors: Characterization and Identification of a New Class of Antagonists. J. Neurochem. 1989, 52, 1319–1328.
- Prescott, C.; Weeks, A.M.; Staley, K.J.; Partin, K.M. Kynurenic acid has a dual action on AMPA receptor responses. Neurosci. Lett. 2006, 402, 108–112.
- Rózsa, E.; Robotka, H.; Vécsei, L.; Toldi, J. The Janus-face kynurenic acid. J. Neural Transm. 2008, 115, 1087–1091.
- Hubbard, T.D.; Murray, I.A.; Bisson, W.H.; Lahoti, T.S.; Gowda, K.; Amin, S.G.; Patterson, A.D.; Perdew, G.H. Adaptation of the human aryl hydrocarbon receptor to sense microbiota-derived indoles. Sci. Rep. 2015, 5, 12689.
- Bessede, A.; Gargaro, M.; Pallotta, M.T.; Matino, D.; Servillo, G.; Brunacci, C.; Bicciato, S.; Mazza, E.M.; Macchiarulo, A.; Vacca, C.; et al. Aryl hydrocarbon receptor control of a disease tolerance defence pathway. Nature 2014, 511, 184–190.
- Jaronen, M.; Quintana, F.J. Immunological Relevance of the Coevolution of IDO1 and AHR. Front. Immunol. 2014, 5, 521.
- Guo, L.; Appelman, B.; Mooij-Kalverda, K.; Houtkooper, R.H.; van Weeghel, M.; Vaz, F.M.; Dijkhuis, A.; Dekker, T.; Smids, B.S.; Duitman, J.W.; et al. COVID-19 Biobank study Group. Prolonged indoleamine 2,3-dioxygenase-2 activity and associated cellular stress in post-acute sequelae of SARS-CoV-2 infection. EBioMedicine 2023, 94, 104729.
- Stone, T.W. Does kynurenic acid act on nicotinic receptors? An assessment of the evidence. J. Neurochem. 2020, 152, 627–649.
- Spekker, E.; Nagy-Grócz, G. All Roads Lead to the Gut: The Importance of the Microbiota and Diet in Migraine. Neurol. Int. 2023, 15, 1174–1190.
- Cronin, P.; Joyce, S.A.; O’Toole, P.W.; O’Connor, E.M. Dietary Fibre Modulates the Gut Microbiota. Nutrients 2021, 13, 1655.
- Murphy, E.A.; Velazquez, K.T.; Herbert, K.M. Influence of high-fat diet on gut microbiota: A driving force for chronic disease risk. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 515–520.
- Dehhaghi, M.; Kazemi Shariat Panahi, H.; Guillemin, G.J. Microorganisms’ Footprint in Neurodegenerative Diseases. Front. Cell Neurosci. 2018, 12, 466.
- Gensollen, T.; Iyer, S.S.; Kasper, D.L.; Blumberg, R.S. How colonization by microbiota in early life shapes the immune system. Science 2016, 352, 539–544.
- Clarke, G.; Grenham, S.; Scully, P.; Fitzgerald, P.; Moloney, R.D.; Shanahan, F.; Dinan, T.G.; Cryan, J.F. The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Mol. Psychiatry 2013, 18, 666–673.
- Desbonnet, L.; Clarke, G.; Traplin, A.; O’Sullivan, O.; Crispie, F.; Moloney, R.D.; Cotter, P.D.; Dinan, T.G.; Cryan, J.F. Gut microbiota depletion from early adolescence in mice: Implications for brain and behaviour. Brain Behav. Immun. 2015, 48, 165–173.
- Sun, P.; Wang, M.; Liu, Y.X.; Li, L.; Chai, X.; Zheng, W.; Chen, S.; Zhu, X.; Zhao, S. High-fat diet-disturbed gut microbiota-colonocyte interactions contribute to dysregulating peripheral tryptophan-kynurenine metabolism. Microbiome 2023, 11, 154.
- Desbonnet, L.; Garrett, L.; Clarke, G.; Bienenstock, J.; Dinan, T.G. The probiotic Bifidobacteria infantis: An assessment of potential antidepressant properties in the rat. J. Psychiatr. Res. 2008, 43, 164–174.
- Kennedy, P.J.; Cryan, J.F.; Dinan, T.G.; Clarke, G. Irritable bowel syndrome: A microbiome-gut-brain axis disorder? World J. Gastroenterol. 2014, 20, 14105–14125.
- Clarke, G.; McKernan, D.P.; Gaszner, G.; Quigley, E.M.; Cryan, J.F.; Dinan, T.G. A Distinct Profile of Tryptophan Metabolism along the Kynurenine Pathway Downstream of Toll-Like Receptor Activation in Irritable Bowel Syndrome. Front. Pharmacol. 2012, 3, 90.
- Fitzgerald, P.; Cassidy, E.M.; Clarke, G.; Scully, P.; Barry, S.; Quigley, E.M.M.; Shanahan, F.; Cryan, J.; Dinan, T.G. Tryptophan catabolism in females with irritable bowel syndrome: Relationship to interferon-gamma, severity of symptoms and psychiatric co-morbidity. Neurogastroenterol. Motil. 2008, 20, 1291–1297.
- O’Mahony, S.M.; Clarke, G.; Borre, Y.E.; Dinan, T.G.; Cryan, J.F. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav. Brain Res. 2015, 277, 32–48.
- Bernstein, C.N. Treatment of IBD: Where we are and where we are going. Am. J. Gastroenterol. 2015, 110, 114–126.
- Xu, S.; Li, X.; Zhang, S.; Qi, C.; Zhang, Z.; Ma, R.; Xiang, L.; Chen, L.; Zhu, Y.; Tang, C.; et al. Oxidative stress gene expression, DNA methylation, and gut microbiota interaction trigger Crohn’s disease: A multi-omics Mendelian randomization study. BMC Med. 2023, 21, 179.
- Yu, F.; Du, Y.; Li, C.; Zhang, H.; Lai, W.; Li, S.; Ye, Z.; Fu, W.; Li, S.; Li, X.G.; et al. Association between metabolites in tryptophan-kynurenine pathway and inflammatory bowel disease: A two-sample Mendelian randomization. Sci. Rep. 2024, 14, 201.
More
Information
Subjects:
Neurosciences
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
637
Revisions:
2 times
(View History)
Update Date:
05 Mar 2024
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No