Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Stefano Stanca | -- | 2451 | 2024-01-31 05:31:21 | | | |
| 2 | Camila Xu | Meta information modification | 2451 | 2024-02-01 02:24:44 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Stanca, S.; Rossetti, M.; Bokulic Panichi, L.; Bongioanni, P. Endothelial Cells in Schizophrenia. Encyclopedia. Available online: https://encyclopedia.pub/entry/54558 (accessed on 24 July 2026).
Stanca S, Rossetti M, Bokulic Panichi L, Bongioanni P. Endothelial Cells in Schizophrenia. Encyclopedia. Available at: https://encyclopedia.pub/entry/54558. Accessed July 24, 2026.
Stanca, Stefano, Martina Rossetti, Leona Bokulic Panichi, Paolo Bongioanni. "Endothelial Cells in Schizophrenia" Encyclopedia, https://encyclopedia.pub/entry/54558 (accessed July 24, 2026).
Stanca, S., Rossetti, M., Bokulic Panichi, L., & Bongioanni, P. (2024, January 31). Endothelial Cells in Schizophrenia. In Encyclopedia. https://encyclopedia.pub/entry/54558
Stanca, Stefano, et al. "Endothelial Cells in Schizophrenia." Encyclopedia. Web. 31 January, 2024.
Copy Citation
Schizophrenia (SCZ) is an articulated psychiatric syndrome whose faded etiologic framework is characterized by a combination of genetic, epigenetic, and environmental factors. It is notoriously explained by an intertwining of a positive and negative symptomatology, from Crow’s SCZ classification in type I (with a syndromic picture marked by a positive clinical condition) and type II (with a negative evolution of the clinical conditions), and subsequent debate, until the last Diagnostic Statistic Manual classification.
schizophrenia
blood–brain barrier
endothelial cells
permeability
astrocytes
1. Introduction
Schizophrenia (SCZ) is an articulated psychiatric syndrome whose faded etiologic framework is characterized by a combination of genetic, epigenetic, and environmental factors. It is notoriously explained by an intertwining of a positive and negative symptomatology, from Crow’s SCZ classification in type I (with a syndromic picture marked by a positive clinical condition) and type II (with a negative evolution of the clinical conditions) [1], and subsequent debate [2][3][4], until the last Diagnostic Statistic Manual classification.
In managing schizophrenia, the Positive and Negative Syndrome Scale (PANSS) plays a crucial role in assessing the spectrum of symptoms. This includes positive symptoms such as hallucinations, delusions, disorganized perception, cognition, and behavior, as well as negative symptoms like anhedonia, abulia, apathy, and alogia [5]. A significant aspect of schizophrenia treatment is the varying response of these symptom categories to antipsychotic medication. While positive symptoms generally show a more favorable response to therapy, negative symptoms are often more resistant to drug treatment. This differential responsiveness to antipsychotics is a key consideration in the clinical management of schizophrenia.
The use of antipsychotics, which are divided into first-generation (typical) and second-generation (atypical), is primarily aimed at correcting neurotransmitter imbalances. However, their usage is associated with a range of side effects that affect cardiovascular, metabolic, endocrinologic, and neurologic systems [6]. Neurological side effects are particularly significant, including tardive dyskinesia, extrapyramidal syndrome, and cognitive impairments associated with antipsychotic use [7].
The interplay between these adverse effects and the response of schizophrenia symptoms to treatment is complex and warrants further elaboration. Notably, the neurological and cognitive side effects of antipsychotics may aggravate negative symptoms or even mimic them, complicating the clinical picture. This exacerbation of negative symptoms by the adverse effects of antipsychotics highlights a critical challenge in treating schizophrenia while minimizing harm from side effects. Such insight is vital for identifying new therapeutic targets and developing more effective psychiatric medications, thereby improving treatment strategies for both positive and negative symptoms and reducing adverse effects.
Due to its complexity, several theories have been suggested to trace the biochemical complexity of this neuropsychiatric disorder. These include the dopamine hypothesis [8], which posits an imbalance in dopamine levels; the glutamate hypothesis [9], suggesting altered glutamatergic neurotransmission; and the neurodevelopmental theory [10], which implicates developmental disruptions in the etiology of the disorder.
2. Endothelial Cells in Schizophrenia: When the First Line of Defense Starts to Give Up
The brain endothelium (Figure 1), far more than a simple barrier, is a complex, specialized structure pivotal to cognitive function. This unique aspect of the BBB [11], as first described by Ehrlich, Goldmann, and Lewandowski [12][13], is maintained by ECs in the central nervous system (CNS). These ECs, unlike those in other body areas, exhibit a higher density of tight junctions (TJs) [14][15], an absence of fenestrations, and reduced caveolar crowding, as highlighted by Ehrlich’s use of hydrophilic tracers [16] and Goldmann’s counterproofs [17]. Lewandowski’s subsequent work further clarified the importance of brain capillaries in controlling molecular flux [13][18]. In SCZ, BBB integrity becomes a focal point of concern. The BBB’s selective permeability is regulated by a complex interplay of cytokines and cellular components. Cytokines such as TNF-α and IFN-γ can impact the expression of occludin [19] and claudin in ECs [20][21][22], critical to maintaining the integrity of TJs. Astrocytes play a complementary role, as their dysfunction can lead to further BBB disruption [23]. This is particularly evident in the response of ECs and astrocytes to cytokines like IL-1β [24][25], which have been shown to affect BBB permeability and stability. The increased permeability of the BBB in SCZ, indicated by elevated albumin and IgG ratios in the CSF [26], points to a breakdown in the barrier’s selective function. This phenomenon could result from intrinsic endothelial abnormalities [27][28] or systemic influences on the BBB [29][30]. Peripheral cytokines are known to adversely affect the BBB [31], exacerbating its vulnerability in SCZ. Structural endothelial abnormalities, as noted in studies examining BBB components [32][33], further highlight the BBB’s susceptibility in this disorder. A detailed examination of the BBB in SCZ reveals an intricate relationship between ECs, immune cell activity, and cognitive processes. The heightened expression of cell adhesion molecules (CAMs) and integrins in SCZ [24][34] leads to increased lymphocyte recruitment, further implicating the immune system in the pathophysiology of SCZ. The role of ICAM-1 in the prefrontal cortex [35] and its elevated serum levels [36][37], correlated with cognitive decline, underscores the multifaceted nature of BBB dysfunction in SCZ. The broader implications of BBB alterations in SCZ extend to synaptic transmission and neuronal plasticity. Neuroinflammation, characterized by an influx of peripheral immune cells and a cascade of cytokines and chemokines [38][39][40], disrupts synaptic function and neuronal communication. This disruption is closely linked to the observed cognitive impairments in SCZ. The involvement of specific cytokines like TNF-α and IL-1β in modulating BBB permeability suggests a direct link between inflammatory processes and synaptic anomalies [41][42][43]. These findings provide a foundation for exploring therapeutic interventions targeting cytokine pathways, with the potential to mitigate both BBB dysfunction and its consequent impact on neuronal function. Moreover, the BBB’s role in SCZ is further complicated by the dynamic interactions within the neurovascular unit (NVU). The NVU, encompassing ECs, astrocytes, pericytes, and neurons, functions as a cohesive entity regulating cerebral blood flow and the neuronal microenvironment [44][45][46]. Dysregulation within the NVU, whether through cytokine imbalances or cellular dysfunction, can lead to altered neurotransmitter levels, disrupted neuronal signaling, and ultimately, the cognitive and behavioral manifestations of SCZ. The relationship between the BBB and neuroinflammation in SCZ is particularly noteworthy. Neuroinflammatory processes, including the activation of microglia and infiltration of peripheral immune cells, contribute to BBB disruption [47][48][49]. This creates a self-perpetuating cycle where BBB breakdown facilitates further neuroinflammation, exacerbating the psychiatric symptoms of SCZ. This focus is critical for understanding the potential role of cytokines in the pathogenesis of this condition. In SCZ, a disorder in which neuroinflammation and autoimmunity can play an important role in the affecting the neurotransmitter balance, cytokines have been investigated, thus bringing to light their increase in the psychosis pathogenetic framework [50][51]. TNF-α and IFN-γ levels are higher in subjects affected by SCZ as compared to controls [47][52], although these findings are not always supported [48]. In terms of cognitive performance, studies have evidenced a correlation between cytokines and cognition impairment. In this respect, in fact, PCR [49], IL-6 [53][54], TNF-α, and IL-1β are associated with a worsened cognitive performance [55]. However, given the complexity of the inflammatory picture, studies have found a correlation between TNF-α and the protection of cognitive functions [56]. Understanding these complex interactions provides crucial insights into the neurobiological underpinnings of SCZ and opens up new avenues for therapeutic intervention. In light of these considerations, future research and therapeutic strategies in SCZ must address the multifaceted nature of BBB dysfunction. This includes exploring ways to strengthen BBB integrity, modulate inflammatory responses, and restore NVU homeostasis. Potential approaches could involve targeting specific cytokine pathways, enhancing TJ protein functionality, and supporting astrocyte health to mitigate the effects of neuroinflammation on the BBB. In conclusion, the BBB’s role in SCZ is a tapestry woven from cellular interactions, cytokine dynamics, and neuroinflammatory processes, each thread contributing to the disorder’s complex pathology. A deeper understanding of these mechanisms is crucial for developing effective treatments for SCZ, moving beyond symptom management to addressing the underlying causes of this challenging condition.
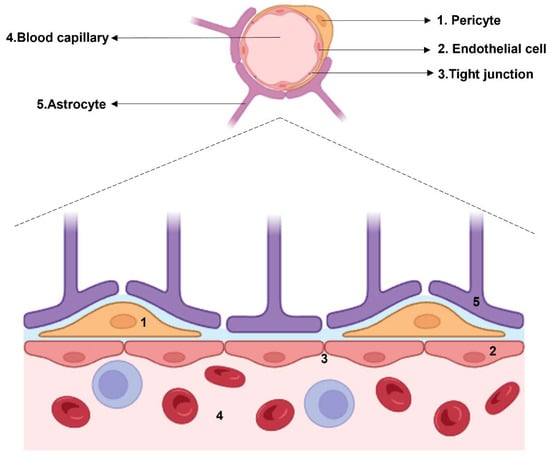
Figure 1. Histological structure of the BBB. The cellular components which surround the blood capillary are: pericytes, endothelial cells connected by tight junctions, and astrocytes.
3. Pericytes: The Cellular Interface in the Crosstalk between Endothelial Cells and Astrocytes
Pericytes, physically the closest cells to the ECs and sharing the same basal membrane, represent, together with astrocytes, an essential element in the constitution of the BBB. This is experimentally brought to light in a co-culture of a BBB syngeneic model, whose combination of pericytes and astrocytes with ECs determined the synthesis of the protein complexes involved in the formation of TJs and their consequent expression on the cellular surface [42]. ECs are linked to pericytes by N-cadherin and connexins [43]. Considering this strict association between ECs and pericytes, there is a peculiar microenvironment guaranteeing an EC–pericyte crosstalk. An increased permeability of this structure would lead to a presence in brain parenchyma of neurotoxic molecules [44], as well as neuroactive ones such as Glu itself, norepinephrine, epinephrine, and glycine (Gly) [45], leading to an increased exposure to several systemic agents involved in neuroinflammation [57]. Pericytes play a pivotal role at the neurovascular interface, providing both cerebral protection and vascular integrity. This dual function is highlighted by the occurrence of aneurysms in platelet-derived growth factor (PDGF)-deficient mice, as noted in recent studies [57]. Additionally, astrocytes constitute another critical component, significantly contributing to synaptogenesis and synaptic pruning. Their involvement underscores the complexity and importance of these cellular elements in maintaining neural and vascular health. Pericytes represent a significant interface between ECs and astrocytes in structuring NVU. They are crucial during angiogenesis, in BBB preservation, as well as in neurogenesis and neuroprotection. The NVU is essentially a network providing cellular–molecular dialogue, able to guarantee communication between the neuronal context and its environment. Pericytes are also crucial in providing extracellular vesicle (EV)-mediated intercellular communication [46][55]. The pericyte EVs are involved in the transport of nucleic acids, proteins, and fatty acids [46]. In other terms, they absolve to the transporting function, a peculiar characteristic of the BBB. Through this system of vehiculating brain-derived neurotrophic factor (BDNF), glia-derived neurotrophic factor (GDNF), and other neurotrophic factors, pericytes promote neuroprotection. A foundational aspect of this investigation involves examining the expression of specific markers, such as tetraspanins, CD8, CD9, and CD63, which are crucial for identification and understanding of these cellular interactions. This approach provides a nuanced perspective on the pathophysiology of SCZ. The above-mentioned molecules affect EVs and, consequently, neurotransmitter regulation by protein trafficking [58]. The importance of pericytes mirrors the peculiarity of EVs in the neuropsychiatric disorders. EVs, vehiculating miRNA, can act as modulator of the pathogenetic process [59][60].
4. Astrocytes, the Last Step before Neurons: A Cell Population between Vascularization and Cognition
Astrocytes cover an important role in structuring the synaptic network, affecting neuronal plasticity, neurotransmitter homeostasis from synaptogenesis to synaptic pruning. In the light of the tripartite synapsis theorization, according to which there is not only an astrocyte structural contribution to forming synapses, but also a mutual interaction between neurons and astrocytes [61], this macroglial cellular entity fulfills this role from the postnatal phase [62]. The pathogenetic involvement of astrocytes in SCZ reverberates its consequences on the BBB, together with the impact on its permeability and selectivity represented by the EC and pericyte damage occurring in the psychotic picture. In this respect, a reduction in the number of astrocytes in several brain areas from the cingulate cortex and motor cortex to the nucleus accumbens has been found [63]. Given the prominent role of astrocytes in synaptogenesis and synaptic pruning, on the one hand, their significance in the SCZ pathogenesis is clear [64] and, on the other, postulating an astrocytic disfunction in SCZ means framing SCZ as neurodevelopmental disorder in which the crucial phenomena involved in structuring the neural network are affected [65][66]. There are different levels of findings in favor of the pathogenetic role of astrocytes in the onset of SCZ, such as the existence of astrocyte-related molecules associated with the pathogenesis of SCZ [67], e.g., S100 calcium binding protein B (S100B) [68][69], whose levels are significantly higher in SCZ [70][71]; thrombospondin 1, important in synaptogenesis [72]; and serine racemase, the enzyme synthetizing D-serine, an important modulator of the ionotropic Glu N-methyl-D-aspartate receptor (NMDAR) [73][74]. The increased level of astrogliosis in SCZ has been marked in about 70% of SCZ subjects in postmortem investigations [63]. On a transcriptional level, an altered expression of glial fibrillary acidic protein (GFAP) mRNA has been detected in astrocytes in SCZ [75], confirmed by the reduced serum level of GFAP and S100B. Evidence of an experimental reduction in astrocytes results in a reduced spike, burst, and generally in decreased astrocyte activation [76]. In SCZ, in fact, important changes to astrocyte density and morphology have been underlined [63][77]. Therefore, it is possible to build a unitary model beyond the different findings concerning astrocytes and the GFAP levels in studies on SCZ: reactive astrogliosis is a peculiar finding in SCZ brains with enhanced levels of GFAP and S100B [63].
The reduced expression of Glu transporters in astrocytes elicits the neurotoxic effect of Glu. Astrocytes are crucial in Glu-ergic transmission: they can modulate NMDARs both positively through the D-serine pathway and negatively via kynurenic acid [63]. Due to their role in Glu metabolism, astrocytes are fundamental in the alveus of the Glu theory of SCZ. Postulated as a consequence of the psychotic-like effects of phencyclidine, this theoretical approach provides the idea that SCZ is associated substantially to a dysregulated effect of Glu, which is, as previously hinted, fundamental to the role played by astrocytes. On this matter, D-amino acid oxidase (DAO), the enzyme implicated in the catabolism of D-serine, expressed both in astrocytes and neurons [78], is overexpressed in SCZ with a consequent reduced level of D-serine and dysregulation of NMDARs [79]. D-serine is synthetized from L-serine, whose availability is the result of the activity of phosphoglycerate dehydrogenase (PHGDH), whose single-nucleotide polymorphism has been weakly associated with the onset of SCZ [80], although its impairment has been correlated to a reduced level of D-serine in the synaptic bouton [81][82]. Furthermore, due to Gly being another positive modulator of NMDARs, research has focused also on its transporter GlyT1 in astrocytes as possible target in drug therapy. This perspective has been confirmed by the antipsychotic effect GlyT-1 inhibitor sarcosine [83][84]. In confirmation of the diriment role played by astrocytes in SCZ, recent findings have also brought to light the importance of astrocytes in contributing to the equilibrium of DA [85]. Indeed, astrocytes express both D1- and D2-like receptors [86], but, more importantly, in the nucleus accumbens, their responsiveness to DA has been highlighted with a stimulation of the presynaptic ATP/adenosine receptor A1, associated with a downregulating effect on excitatory transmission [87]. The activating effect of DA on astrocytes has been evidenced also in the prefrontal cortex [88]. Finally, it is interesting to mention a study on disrupted astrocytes in schizophrenia 1 (DISC1), whose polymorphisms have been associated with SCZ [89][90]. According to this work, the elimination of such a molecule, important for neurotransmission, exclusively in astrocytes, has implications on the overexpression of D2-like receptors on the astrocytic membrane and on the downregulation of the DA-ergic activity in the basal ganglia.
References
- Crow, T.J. Positive and Negative Schizophrenic Symptoms and the Role of Dopamine. Br. J. Psychiatry 1980, 137, 383–386.
- Buchwald, K.; Narayanan, A.; Siegert, R.J.; Vignes, M.; Arrowsmith, K.; Sandham, M. Centrality Statistics of Symptom Networks of Schizophrenia: A Systematic Review. Psychol. Med. 2024, 1–13.
- Wolpe, N.; Vituri, A.; Jones, P.B.; Shahar, M.; Fernandez-Egea, E. The Longitudinal Structure of Negative Symptoms in Treatment Resistant Schizophrenia. Compr. Psychiatry 2024, 128, 152440.
- Di Luzio, M.; Pontillo, M.; Villa, M.; Attardi, A.G.; Bellantoni, D.; Di Vincenzo, C.; Vicari, S. Clinical Features and Comorbidity in Very Early-Onset Schizophrenia: A Systematic Review. Front. Psychiatry 2023, 14, 799.
- Kay, S.R.; Fiszbein, A.; Opler, L.A. The Positive and Negative Syndrome Scale (PANSS) for Schizophrenia. Schizophr. Bull. 1987, 13, 261–276.
- Stroup, T.S.; Gray, N. Management of Common Adverse Effects of Antipsychotic Medications. World Psychiatry 2018, 17, 341–356.
- Arana, G.W. An Overview of Side Effects Caused by Typical Antipsychotics. J. Clin. Psychiatry 2000, 61 (Suppl. S8), 5–11, Discussion: J. Clin. Psychiatry 2000, 61 (Suppl. 8), 12–13..
- Brisch, R.; Saniotis, A.; Wolf, R.; Bielau, H.; Bernstein, H.-G.; Steiner, J.; Bogerts, B.; Braun, A.K.; Jankowski, Z.; Kumaritlake, J.; et al. The Role of Dopamine in Schizophrenia from a Neurobiological and Evolutionary Perspective: Old Fashioned, but Still in Vogue. Front. Psychiatry 2014, 5, 47.
- Hu, W.; MacDonald, M.L.; Elswick, D.E.; Sweet, R.A. The Glutamate Hypothesis of Schizophrenia: Evidence from Human Brain Tissue Studies. Ann. N. Y. Acad. Sci. 2015, 1338, 38–57.
- Owen, M.J.; O’Donovan, M.C.; Thapar, A.; Craddock, N. Neurodevelopmental Hypothesis of Schizophrenia. Br. J. Psychiatry 2011, 198, 173–175.
- Daneman, R. The Blood–Brain Barrier in Health and Disease. Ann. Neurol. 2012, 72, 648–672.
- Kadry, H.; Noorani, B.; Cucullo, L. A Blood–Brain Barrier Overview on Structure, Function, Impairment, and Biomarkers of Integrity. Fluids Barriers CNS 2020, 17, 69.
- Bechmann, I.; Galea, I.; Perry, V.H. What Is the Blood–Brain Barrier (Not)? Trends Immunol. 2007, 28, 5–11.
- Abbott, N.J.; Patabendige, A.A.K.; Dolman, D.E.M.; Yusof, S.R.; Begley, D.J. Structure and Function of the Blood–Brain Barrier. Neurobiol. Dis. 2010, 37, 13–25.
- Engelhardt, B.; Liebner, S. Novel Insights into the Development and Maintenance of the Blood–Brain Barrier. Cell Tissue Res. 2014, 355, 687–699.
- Dyrna, F.; Hanske, S.; Krueger, M.; Bechmann, I. The Blood-Brain Barrier. J. Neuroimmune Pharmacol. 2013, 8, 763–773.
- Bentivoglio, M.; Kristensson, K. Tryps and Trips: Cell Trafficking across the 100-Year-Old Blood–Brain Barrier. Trends Neurosci. 2014, 37, 325–333.
- Lewandowsky, M. Zur Lehre Der Cerebrospinalflussigkeit. Z. Klin. Med. 1909, 40, 480–494.
- Alluri, H.; Peddaboina, C.S.; Tharakan, B. Evaluation of Tight Junction Integrity in Brain Endothelial Cells Using Confocal Microscopy. In Vascular Hyperpermeability: Methods and Protocols; Springer: New York, NY, USA, 2024; pp. 257–262.
- Abbott, N.J.; Rönnbäck, L.; Hansson, E. Astrocyte–Endothelial Interactions at the Blood–Brain Barrier. Nat. Rev. Neurosci. 2006, 7, 41–53.
- Friedman, A.; Kaufer, D. Blood-Brain Barrier Breakdown and Blood-Brain Communication in Neurological and Psychiatric Diseases. Cardiovasc. Psychiatry Neurol. 2011, 2011, 431470.
- Iadecola, C. The Neurovascular Unit Coming of Age: A Journey through Neurovascular Coupling in Health and Disease. Neuron 2017, 96, 17–42.
- Abdullahi, W.; Tripathi, D.; Ronaldson, P.T. Blood-Brain Barrier Dysfunction in Ischemic Stroke: Targeting Tight Junctions and Transporters for Vascular Protection. Am. J. Physiol.-Cell Physiol. 2018, 315, C343–C356.
- Stamatovic, S.M.; Johnson, A.M.; Keep, R.F.; Andjelkovic, A. V Junctional Proteins of the Blood-Brain Barrier: New Insights into Function and Dysfunction. Tissue Barriers 2016, 4, e1154641.
- Ohtsuki, S.; Yamaguchi, H.; Katsukura, Y.; Asashima, T.; Terasaki, T. MRNA Expression Levels of Tight Junction Protein Genes in Mouse Brain Capillary Endothelial Cells Highly Purified by Magnetic Cell Sorting. J. Neurochem. 2008, 104, 147–154.
- Zihni, C.; Mills, C.; Matter, K.; Balda, M.S. Tight Junctions: From Simple Barriers to Multifunctional Molecular Gates. Nat. Rev. Mol. Cell Biol. 2016, 17, 564–580.
- Martìn-Padura, I.; Lostaglio, S.; Schneemann, M.; Williams, L.; Romano, M.; Fruscella, P.; Panzeri, C.; Stoppacciaro, A.; Ruco, L.; Villa, A.; et al. Junctional Adhesion Molecule, a Novel Member of the Immunoglobulin Superfamily That Distributes at Intercellular Junctions and Modulates Monocyte Transmigration. J. Cell Biol. 1998, 142, 117–127.
- Poliak, S.; Matlis, S.; Ullmer, C.; Scherer, S.S.; Peles, E. Distinct Claudins and Associated PDZ Proteins Form Different Autotypic Tight Junctions in Myelinating Schwann Cells. J. Cell Biol. 2002, 159, 361–372.
- Kaya, M.; Ahishali, B. Basic Physiology of the Blood-Brain Barrier in Health and Disease: A Brief Overview. Tissue Barriers 2021, 9, 1840913.
- Yuan, S.; Liu, K.; Qi, Z. Occludin Regulation of Blood–Brain Barrier and Potential Therapeutic Target in Ischemic Stroke. Brain Circ. 2020, 6, 152.
- Ni, Y.; Teng, T.; Li, R.; Simonyi, A.; Sun, G.Y.; Lee, J.C. TNFα Alters Occludin and Cerebral Endothelial Permeability: Role of P38MAPK. PLoS ONE 2017, 12, e0170346.
- Utech, M.; Bruewer, M.; Parkos, C.; Hopkins, A.; Ivanov, A.; Nusrat, A. IFN-Gamma Induces Endocytosis of Epithelial Tight Junction Transmembrane Proteins into a Vacuolar Apical Compartment. J. Am. Coll. Surg. 2004, 199, 20.
- Ni, C.; Wang, C.; Zhang, J.; Qu, L.; Liu, X.; Lu, Y.; Yang, W.; Deng, J.; Lorenz, D.; Gao, P.; et al. Interferon-γ Safeguards Blood-Brain Barrier during Experimental Autoimmune Encephalomyelitis. Am. J. Pathol. 2014, 184, 3308–3320.
- Wang, Y.; Jin, S.; Sonobe, Y.; Cheng, Y.; Horiuchi, H.; Parajuli, B.; Kawanokuchi, J.; Mizuno, T.; Takeuchi, H.; Suzumura, A. Interleukin-1β Induces Blood–Brain Barrier Disruption by Downregulating Sonic Hedgehog in Astrocytes. PLoS ONE 2014, 9, e110024.
- Hartmann, C.; Schwietzer, Y.A.; Otani, T.; Furuse, M.; Ebnet, K. Physiological Functions of Junctional Adhesion Molecules (JAMs) in Tight Junctions. Biochim. Biophys. Acta (BBA)—Biomembr. 2020, 1862, 183299.
- Worzfeld, T.; Schwaninger, M. Apicobasal Polarity of Brain Endothelial Cells. J. Cereb. Blood Flow Metab. 2016, 36, 340–362.
- Cruz-Orengo, L.; Daniels, B.P.; Dorsey, D.; Basak, S.A.; Grajales-Reyes, J.G.; McCandless, E.E.; Piccio, L.; Schmidt, R.E.; Cross, A.H.; Crosby, S.D.; et al. Enhanced Sphingosine-1-Phosphate Receptor 2 Expression Underlies Female CNS Autoimmunity Susceptibility. J. Clin. Investig. 2014, 124, 2571–2584.
- Uranova, N.A.; Vikhreva, O.V.; Rachmanova, V.I.; Orlovskaya, D.D. Ultrastructural Alterations of Myelinated Fibers and Oligodendrocytes in the Prefrontal Cortex in Schizophrenia: A Postmortem Morphometric Study. Schizophr. Res. Treat. 2011, 2011, 325789.
- De Picker, L.J.; Victoriano, G.M.; Richards, R.; Gorvett, A.J.; Lyons, S.; Buckland, G.R.; Tofani, T.; Norman, J.L.; Chatelet, D.S.; Nicoll, J.A.R.; et al. Immune Environment of the Brain in Schizophrenia and during the Psychotic Episode: A Human Post-Mortem Study. Brain Behav. Immun. 2021, 97, 319–327.
- Cai, H.Q.; Weickert, T.W.; Catts, V.S.; Balzan, R.; Galletly, C.; Liu, D.; O’Donnell, M.; Shannon Weickert, C. Altered Levels of Immune Cell Adhesion Molecules Are Associated with Memory Impairment in Schizophrenia and Healthy Controls. Brain Behav. Immun. 2020, 89, 200–208.
- Weickert, T.; Cai, H.; O’Donnell, M.; Balzan, R.; Wells, R.; Liu, D.; Galletly, C.; Weickert, C.S. O1.5. ICAM-1 is increased in brain and peripheral levels of soluble ICAM-1 is related to cognitive deficits in schizophrenia. Schizophr. Bull. 2018, 44, S73–S74.
- Nakagawa, S.; Deli, M.A.; Kawaguchi, H.; Shimizudani, T.; Shimono, T.; Kittel, Á.; Tanaka, K.; Niwa, M. A New Blood–Brain Barrier Model Using Primary Rat Brain Endothelial Cells, Pericytes and Astrocytes. Neurochem. Int. 2009, 54, 253–263.
- Armulik, A.; Genové, G.; Mäe, M.; Nisancioglu, M.H.; Wallgard, E.; Niaudet, C.; He, L.; Norlin, J.; Lindblom, P.; Strittmatter, K.; et al. Pericytes Regulate the Blood–Brain Barrier. Nature 2010, 468, 557–561.
- Lacombe, R.J.S.; Chouinard-Watkins, R.; Bazinet, R.P. Brain Docosahexaenoic Acid Uptake and Metabolism. Mol. Asp. Med. 2018, 64, 109–134.
- Ahishali, B.; Kaya, M. Evaluation of Blood-Brain Barrier Integrity Using Vascular Permeability Markers: Evans Blue, Sodium Fluorescein, Albumin-Alexa Fluor Conjugates, and Horseradish Peroxidase. In Permeability Barrier: Methods and Protocols; Humana: New York, NY, USA, 2020; pp. 87–103.
- Sharma, K.; Zhang, Y.; Paudel, K.R.; Kachelmeier, A.; Hansbro, P.M.; Shi, X. The Emerging Role of Pericyte-Derived Extracellular Vesicles in Vascular and Neurological Health. Cells 2022, 11, 3108.
- Najjar, S.; Pahlajani, S.; De Sanctis, V.; Stern, J.N.H.; Najjar, A.; Chong, D. Neurovascular Unit Dysfunction and Blood–Brain Barrier Hyperpermeability Contribute to Schizophrenia Neurobiology: A Theoretical Integration of Clinical and Experimental Evidence. Front. Psychiatry 2017, 8, 83.
- Wilson, L.; Szigeti, A.; Kearney, A.; Clarke, M. Clinical Characteristics of Primary Psychotic Disorders with Concurrent Substance Abuse and Substance-Induced Psychotic Disorders: A Systematic Review. Schizophr. Res. 2018, 197, 78–86.
- Misiak, B.; Stramecki, F.; Gawęda, Ł.; Prochwicz, K.; Sąsiadek, M.M.; Moustafa, A.A.; Frydecka, D. Interactions Between Variation in Candidate Genes and Environmental Factors in the Etiology of Schizophrenia and Bipolar Disorder: A Systematic Review. Mol. Neurobiol. 2018, 55, 5075–5100.
- Khandaker, G.M.; Cousins, L.; Deakin, J.; Lennox, B.R.; Yolken, R.; Jones, P.B. Inflammation and Immunity in Schizophrenia: Implications for Pathophysiology and Treatment. Lancet Psychiatry 2015, 2, 258–270.
- Ermakov, E.A.; Melamud, M.M.; Buneva, V.N.; Ivanova, S.A. Immune System Abnormalities in Schizophrenia: An Integrative View and Translational Perspectives. Front. Psychiatry 2022, 13, 568.
- Najjar, S.; Pearlman, D.M.; Alper, K.; Najjar, A.; Devinsky, O. Neuroinflammation and Psychiatric Illness. J. Neuroinflamm. 2013, 10, 816.
- Borovcanin, M.M.; Jovanovic, I.; Radosavljevic, G.; Pantic, J.; Minic Janicijevic, S.; Arsenijevic, N.; Lukic, M.L. Interleukin-6 in Schizophrenia—Is There a Therapeutic Relevance? Front. Psychiatry 2017, 8, 221.
- Ribeiro-Santos, R.; de Campos-Carli, S.M.; Ferretjans, R.; Teixeira-Carvalho, A.; Martins-Filho, O.A.; Teixeira, A.L.; Salgado, J.V. The Association of Cognitive Performance and IL-6 Levels in Schizophrenia Is Influenced by Age and Antipsychotic Treatment. Nord. J. Psychiatry 2020, 74, 187–193.
- Patlola, S.R.; Donohoe, G.; McKernan, D.P. The Relationship between Inflammatory Biomarkers and Cognitive Dysfunction in Patients with Schizophrenia: A Systematic Review and Meta-Analysis. Prog. Neuropsychopharmacol. Biol. Psychiatry 2023, 121, 110668.
- Efffendy, E.; Amin, M.M.; Utami, N. Role of Tumor Necrosis Factor-Alpha in Schizophrenia and Cognitive Impairment. Open Access Maced. J. Med. Sci. 2021, 9, 160–163.
- Pollak, T.A.; Drndarski, S.; Stone, J.M.; David, A.S.; McGuire, P.; Abbott, N.J. The Blood–Brain Barrier in Psychosis. Lancet Psychiatry 2018, 5, 79–92.
- Scholz, C.; Jacob, C.P.; Buttenschon, H.N.; Kittel-Schneider, S.; Boreatti-Hümmer, A.; Zimmer, M.; Walter, U.; Lesch, K.; Mors, O.; Kneitz, S.; et al. Functional Variants of TSPAN8 Are Associated with Bipolar Disorder and Schizophrenia. Am. J. Med. Genet. Part. B Neuropsychiatr. Genet. 2010, 153B, 967–972.
- Xia, X.; Wang, Y.; Huang, Y.; Zhang, H.; Lu, H.; Zheng, J.C. Exosomal MiRNAs in Central Nervous System Diseases: Biomarkers, Pathological Mediators, Protective Factors and Therapeutic Agents. Prog. Neurobiol. 2019, 183, 101694.
- Mingardi, J.; La Via, L.; Tornese, P.; Carini, G.; Trontti, K.; Seguini, M.; Tardito, D.; Bono, F.; Fiorentini, C.; Elia, L.; et al. MiR-9-5p Is Involved in the Rescue of Stress-Dependent Dendritic Shortening of Hippocampal Pyramidal Neurons Induced by Acute Antidepressant Treatment with Ketamine. Neurobiol. Stress 2021, 15, 100381.
- Ferrucci, L.; Cantando, I.; Cordella, F.; Di Angelantonio, S.; Ragozzino, D.; Bezzi, P. Microglia at the Tripartite Synapse during Postnatal Development: Implications for Autism Spectrum Disorders and Schizophrenia. Cells 2023, 12, 2827.
- Chung, W.-S.; Allen, N.J.; Eroglu, C. Astrocytes Control Synapse Formation, Function, and Elimination. Cold Spring Harb. Perspect. Biol. 2015, 7, a020370.
- Tarasov, V.V.; Svistunov, A.A.; Chubarev, V.N.; Sologova, S.S.; Mukhortova, P.; Levushkin, D.; Somasundaram, S.G.; Kirkland, C.E.; Bachurin, S.O.; Aliev, G. Alterations of Astrocytes in the Context of Schizophrenic Dementia. Front. Pharmacol. 2020, 10, 1612.
- Mısır, E.; Akay, G.G. Synaptic Dysfunction in Schizophrenia. Synapse 2023, 77, e22276.
- Notter, T. Astrocytes in Schizophrenia. Brain Neurosci. Adv. 2021, 5, 239821282110091.
- de Oliveira Figueiredo, E.C.; Calì, C.; Petrelli, F.; Bezzi, P. Emerging Evidence for Astrocyte Dysfunction in Schizophrenia. Glia 2022, 70, 1585–1604.
- Chang, C.-Y.; Luo, D.-Z.; Pei, J.-C.; Kuo, M.-C.; Hsieh, Y.-C.; Lai, W.-S. Not Just a Bystander: The Emerging Role of Astrocytes and Research Tools in Studying Cognitive Dysfunctions in Schizophrenia. Int. J. Mol. Sci. 2021, 22, 5343.
- Bernstein, H.-G.; Steiner, J.; Bogerts, B. Glial Cells in Schizophrenia: Pathophysiological Significance and Possible Consequences for Therapy. Expert. Rev. Neurother. 2009, 9, 1059–1071.
- Hohoff, C.; Ponath, G.; Freitag, C.M.; Kästner, F.; Krakowitzky, P.; Domschke, K.; Koelkebeck, K.; Kipp, F.; von Eiff, C.; Deckert, J.; et al. Risk Variants in the S100B Gene Predict Elevated S100B Serum Concentrations in Healthy Individuals. Am. J. Med. Genet. Part. B Neuropsychiatr. Genet. 2010, 153B, 291–297.
- Hong, S.; Lee, E.E.; Martin, A.S.; Soontornniyomkij, B.; Soontornniyomkij, V.; Achim, C.L.; Reuter, C.; Irwin, M.R.; Eyler, L.T.; Jeste, D.V. Abnormalities in Chemokine Levels in Schizophrenia and Their Clinical Correlates. Schizophr. Res. 2017, 181, 63–69.
- Yelmo-Cruz, S.; Morera-Fumero, A.L.; Abreu-González, P. S100B and Schizophrenia. Psychiatry Clin. Neurosci. 2013, 67, 67–75.
- Park, H.J.; Kim, S.K.; Kim, J.W.; Kang, W.S.; Chung, J.-H. Association of Thrombospondin 1 Gene with Schizophrenia in Korean Population. Mol. Biol. Rep. 2012, 39, 6875–6880.
- Morita, Y.; Ujike, H.; Tanaka, Y.; Otani, K.; Kishimoto, M.; Morio, A.; Kotaka, T.; Okahisa, Y.; Matsushita, M.; Morikawa, A.; et al. A Genetic Variant of the Serine Racemase Gene Is Associated with Schizophrenia. Biol. Psychiatry 2007, 61, 1200–1203.
- Labrie, V.; Roder, J.C. The Involvement of the NMDA Receptor D-Serine/Glycine Site in the Pathophysiology and Treatment of Schizophrenia. Neurosci. Biobehav. Rev. 2010, 34, 351–372.
- Catts, V.S.; Wong, J.; Fillman, S.G.; Fung, S.J.; Shannon Weickert, C. Increased Expression of Astrocyte Markers in Schizophrenia: Association with Neuroinflammation. Aust. N. Z. J. Psychiatry 2014, 48, 722–734.
- Fritschi, L.; Lindmar, J.H.; Scheidl, F.; Lenk, K. Neuronal and Astrocytic Regulations in Schizophrenia: A Computational Modelling Study. Front. Cell Neurosci. 2021, 15, 718459.
- Kim, J.; Iwata, Y.; Plitman, E.; Caravaggio, F.; Chung, J.K.; Shah, P.; Blumberger, D.M.; Pollock, B.G.; Remington, G.; Graff-Guerrero, A.; et al. A Meta-Analysis of Transcranial Direct Current Stimulation for Schizophrenia: “Is More Better?”. J. Psychiatr. Res. 2019, 110, 117–126.
- Kwak, T.H.; Kang, J.H.; Hali, S.; Kim, J.; Kim, K.-P.; Park, C.; Lee, J.-H.; Ryu, H.K.; Na, J.E.; Jo, J.; et al. Generation of Homogeneous Midbrain Organoids with in Vivo—Like Cellular Composition Facilitates Neurotoxin-Based Parkinson’s Disease Modeling. Stem Cells 2020, 38, 727–740.
- Orzylowski, M.; Fujiwara, E.; Mousseau, D.D.; Baker, G.B. An Overview of the Involvement of D-Serine in Cognitive Impairment in Normal Aging and Dementia. Front. Psychiatry 2021, 12, 754032.
- Kim, J.-W.; Kim, H.-J.; Ban, J.Y.; Park, H.J.; Kim, S.K.; Kang, S.W.; Chung, J.-H.; Park, J.K.; Kim, J.W. Assessment between Phosphoglycerate Dehydrogenase Gene and Schizophrenia in Korean Population. Psychiatr. Genet. 2009, 19, 161.
- Eulenburg, V.; Armsen, W.; Betz, H.; Gomeza, J. Glycine Transporters: Essential Regulators of Neurotransmission. Trends Biochem. Sci. 2005, 30, 325–333.
- MacKay, M.-A.B.; Kravtsenyuk, M.; Thomas, R.; Mitchell, N.D.; Dursun, S.M.; Baker, G.B. D-Serine: Potential Therapeutic Agent and/or Biomarker in Schizophrenia and Depression? Front. Psychiatry 2019, 10, 25.
- Zhang, H.X.; Lyons-Warren, A.; Thio, L.L. The Glycine Transport Inhibitor Sarcosine Is an Inhibitory Glycine Receptor Agonist. Neuropharmacology 2009, 57, 551–555.
- Hashimoto, K. Glycine Transporter-1: A New Potential Therapeutic Target for Schizophrenia. Curr. Pharm. Des. 2011, 17, 112–120.
- Petrelli, F.; Dallérac, G.; Pucci, L.; Calì, C.; Zehnder, T.; Sultan, S.; Lecca, S.; Chicca, A.; Ivanov, A.; Asensio, C.S.; et al. Dysfunction of Homeostatic Control of Dopamine by Astrocytes in the Developing Prefrontal Cortex Leads to Cognitive Impairments. Mol. Psychiatry 2020, 25, 732–749.
- Corkrum, M.; Araque, A. Astrocyte-Neuron Signaling in the Mesolimbic Dopamine System: The Hidden Stars of Dopamine Signaling. Neuropsychopharmacology 2021, 46, 1864–1872.
- Corkrum, M.; Covelo, A.; Lines, J.; Bellocchio, L.; Pisansky, M.; Loke, K.; Quintana, R.; Rothwell, P.E.; Lujan, R.; Marsicano, G.; et al. Dopamine-Evoked Synaptic Regulation in the Nucleus Accumbens Requires Astrocyte Activity. Neuron 2020, 105, 1036–1047.e5.
- Pittolo, S.; Yokoyama, S.; Willoughby, D.D.; Taylor, C.R.; Reitman, M.E.; Tse, V.; Wu, Z.; Etchenique, R.; Li, Y.; Poskanzer, K.E. Dopamine Activates Astrocytes in Prefrontal Cortex via A1-Adrenergic Receptors. Cell Rep. 2022, 40, 111426.
- Wang, H.-Y.; Liu, Y.; Yan, J.-W.; Hu, X.-L.; Zhu, D.-M.; Xu, X.-T.; Li, X.-S. Gene Polymorphisms of DISC1 Is Associated with Schizophrenia: Evidence from a Meta-Analysis. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 81, 64–73.
- Mastrogiacomo, R.; Trigilio, G.; Devroye, C.; Dautan, D.; Ferretti, V.; Losi, G.; Caffino, L.; Orso, G.; Marotta, R.; Maltese, F.; et al. Dysbindin-1A Modulation of Astrocytic Dopamine and Basal Ganglia Dependent Behaviors Relevant to Schizophrenia. Mol. Psychiatry 2022, 27, 4201–4217.
More
Information
Subjects:
Physiology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
635
Revisions:
2 times
(View History)
Update Date:
01 Feb 2024
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No