Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Maria Luiza Mello | -- | 1754 | 2023-10-02 21:10:50 | | | |
| 2 | Camila Xu | Meta information modification | 1754 | 2023-10-07 03:43:45 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Mello, M.L.S. Heterochromatin and Nuclear Phenotypes of T. infestans. Encyclopedia. Available online: https://encyclopedia.pub/entry/49815 (accessed on 27 June 2026).
Mello MLS. Heterochromatin and Nuclear Phenotypes of T. infestans. Encyclopedia. Available at: https://encyclopedia.pub/entry/49815. Accessed June 27, 2026.
Mello, Maria Luiza S.. "Heterochromatin and Nuclear Phenotypes of T. infestans" Encyclopedia, https://encyclopedia.pub/entry/49815 (accessed June 27, 2026).
Mello, M.L.S. (2023, October 02). Heterochromatin and Nuclear Phenotypes of T. infestans. In Encyclopedia. https://encyclopedia.pub/entry/49815
Mello, Maria Luiza S.. "Heterochromatin and Nuclear Phenotypes of T. infestans." Encyclopedia. Web. 02 October, 2023.
Copy Citation
Triatoma infestans (Klug), the most important vector of Chagas disease, is a hemipteran insect belonging to the Triatominae subfamily of the Reduviidae family and one of the 84 species of the Triatomini tribe.
insect vector
somatic polyploidy
chromocenters
1. Introduction
Triatoma infestans (Klug), the most important vector of Chagas disease, is a hemipteran insect belonging to the Triatominae subfamily of the Reduviidae family and one of the 84 species of the Triatomini tribe. This species developed through the Andean and non-Andean evolutionary lineages with different geographic distributions in South America, genomic size, and main repetitive DNA [1]. Both lineages dispersed from Bolivian highlands, from where they had originated. Forty-two satellite DNA families are present in varying amounts in both lineages [2].
The Andean lineage was primarily dispersed in the high-altitude valleys of Bolivia and Peru and in lower altitudes of southern Peru; the non-Andean lineage occupied lowland regions in Argentina, Brazil, Chile, Paraguay, Uruguay, and Bolivia and certain higher altitude regions in Argentina until chemical interventions for insect control were undertaken in the past decades, which drastically reduced these groups [2]. Although these lineages differ in nuclear DNA content per haploid genome (1487 Gbp, non-Andean lineage; 1936 Gbp, Andean lineage) and in the amounts of satellite DNA families, they have the same number of holokinetic chromosomes (2n = 20 + XY or XX) [2].
In addition to the importance of T. infestans as a vector for Trypanosoma cruzi infection, it has been revealed as a model for studies on unusual cellular processes involving somatic polyploidy, nuclear and cell fusion, presence of large heterochromatin bodies (chromocenters), diversity of nuclear phenotypes, cell death, epigenetic markers, and chromatin response to histone deacetylase inhibition.
2. Somatic Polyploidy
Insects of most orders develop somatic polyploidy, in which DNA endoreplication steps advance and permit nuclear and cellular volume enlargement and organ growth without intervention of mitosis [3]. This phenomenon occurs mainly during the larval stages in insects undergoing complete metamorphosis (holometabola), or in nymphal instars of insects undergoing simple metamorphosis (hemimetabola) [3]. Hemipterans develop through hemimetaboly. Somatic polyploidy involving an integral doubling of the genome does not occur in all insect groups; specific genomic regions, as in the case of the heterochromatin, may replicate differentially [4].
Somatic polyploidy may be considered an adaptive process through which the cells save the physiological time and protein synthesis that would be required if a mitotic spindle and nuclear membrane were fabricated as in the case of ordinary somatic division [3]. Some authors have hypothesized that it is a strategy for increasing gene copy numbers, which would facilitate high rates of biosynthesis and metabolism [4]. In healthy mammal tissues, somatic polyploidy has been considered important for cells to function under stress, protecting their viability and permitting their adaptation under new environmental conditions [5][6].
The DNA endoreplication steps that occur in the nuclei of the Malpighian tubule cells of fully-nourished T. infestans specimens are arrested at the end of the fifth nymphal instar, when Feulgen-DNA 32 C and 64 C optimal ploidy degrees are attained per nucleus compared with the Feulgen-DNA content of nuclei in the first nymphal stages and in spermatids [7][8]. Feulgen-DNA classes or ploidy degrees that are identified as 1 C, 2 C, 4 C, and so on, are associated with the DNA content of haploid, diploid, tetraploid, and so on, nuclei of the same species [9]. However, in fasted insects, the ordinary polyploidization pattern detected in the Malpighian tubules is surpassed because nuclear and cellular fusions occur even with low incidence in specimens reared in the laboratory [10]. Then, ploidy degrees shift to considerably higher values, as demonstrated by DNA cytophotometric quantification, visual detection of the presence of giant nuclei (Figure 1a), or counting of the total number of nuclei/organ that decrease significantly [11]. Giant nuclei may attain a ploidy degree as high as 1024 C [11]. Consequently, the total number of nuclei may show a 40% reduction [12]. Additionally, nuclear and cell fusions have been detected in BTC-32 cells cultivated from embryonic tissues of T. infestans at the London School of Hygiene and Tropical Medicine [13]. The presence of giant nuclei is not restricted to T. infestans; it has also been reported in the fat body cells of Rhodnius prolixus, another species of the Reduviidae family [14].
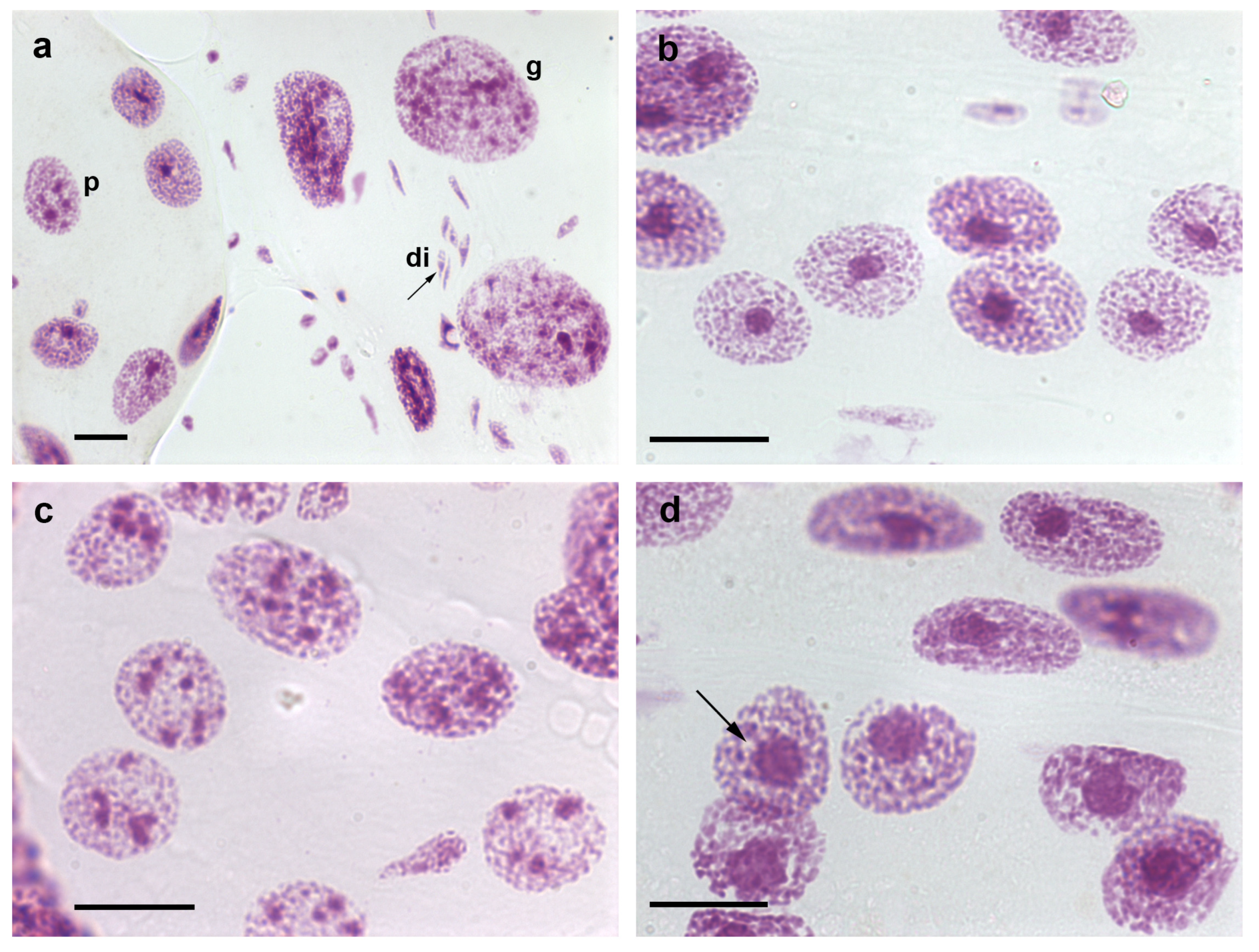
Figure 1. Nuclear phenotypes of Feulgen-stained cells of T. infestans. The Feulgen reaction stains DNA specifically [9]. Co-occurrence of diploid fibroblasts (di), and polyploid (p) and giant (g) Malpighian tubule cell nuclei is shown in (a). Chromocenters are well evident as conspicuous condensed chromatin bodies in single- (b,d) and multi-chromocentered nuclei (c). A nuclear phenotype showing chromocenter heterochromatin unpacking ((d), arrow) can be compared with images of nuclei with a chromocenter usual size ((b) or at the up corner of (d)). Bars, 20 µm.
Initially, nuclear fusion is facilitated by the binucleate composition of the somatic cells [15]. When fasting periods are prolonged, cell fusion and additional nuclear fusions occur in the Malpighian tubules. Cell fusion may be favored by the presence of the viral particles that infect these cells, especially when the insects are fed avian blood under laboratory conditions [15][16]. Cell fusion has been suggested to represent an attempt at cell survival that opposes cell death [11][14]; however, both phenomena may coexist in the same organ [10].
3. Heterochromatin and Nuclear Phenotypes
Condensed chromatin aggregates comprising constitutive heterochromatin (chromocenters) are present in T. infestans cells from the first nymphal instar [7]. These bodies increase in size through DNA endoreplication steps that do not always accompany the DNA doubling process in the euchromatin (DNA underreplication), representing a constitutive heterochromatin characteristic [7]. Other indicators of the constitutive heterochromatin nature of these condensed chromatin bodies are as follows: the absence of RNA transcriptional activity in most regions of these bodies, as detected using radioautography; abundance of repetitive DNA; and differential response to the Feulgen reaction compared with the characteristics of euchromatin [7][17][18].
The chromocenters of T. infestans are usually composed of copies of the large A, B, and C autosomes plus the X and Y sex chromosomes [19][20]. One additional autosome pair has been reported as part of this heterochromatin type under specific insect rearing conditions [21]. Additionally, polymorphism and variable amount of heterochromatin have been reported for T. infestans [22][23][24].
From the first to the third nymphal instar, only one chromocenter is present in the Malpighian tubules of T. infestans; from this stage onward, a portion of the cells exhibit more than one chromocenter (Figure 1b–d) [7][8]. The process of formation of multi-chromocenters apparently involves budding or declustering from the single chromocenter body [25] and does not lead to the micronuclei production that results in cell death as reported for other cell systems [26]. The nuclear phenotype diversification and the size attained by chromocenters in the Malpighian tubules of nymphs and adults have not been identified in other species of the Reduviidae family studied till date [27]. These unexplained phenomena could not be shown to be associated with diversified functions exerted by the Malpighian tubules, which are excretory organs eliminating the byproducts of a blood diet that does not change during the insect lifetime.
Notably, free DNA phosphates showed differential availability for the electrostatic binding of toluidine blue molecules under Mg2+ competitive staining conditions, when the single-chromocentered nuclei were compared with nuclei bearing multiple chromocenters [28]. This finding revealed that the dye-binding sites in the DNA of chromocenters in the multi-chromocentered nuclei are more exposed and easier to block by Mg2+ ions than those in the DNA of chromocenters in single-chromocentered nuclei probably owing to differences in the packing states of the DNA-protein complexes involved [29].
T. infestans nuclei constitute an attractive model for studying the 3D position of chromocenters because of the large volume of these bodies inside the nuclei. Using confocal microscopy and quantitative measurement of the 3D position of the chromocenters in the Malpighian tubule cells of T. infestans, the proximity of these bodies to the nuclear periphery was demonstrated during the insect lifetime [25]. The spatial distribution of chromocenters in relation to the nuclear periphery was confirmed using electron microscopy [25].
The proximity of chromocenters to the nuclear periphery has been detected in the cells of several other organisms [29][30][31][32], including species of the Reduviidae family which exhibit very small chromocenters, as in the case of Panstrongylus megistus [33]; however, the reason for this phenomenon remains unclear [34]. The concept that proximity of a chromocenter to the nuclear periphery is associated with overall gene silencing [35][36] does not fully apply to T. infestans, because 18 active S rDNA sites were detected in the chromocenter surface including the region facing the nuclear periphery, according to videos obtained after 3D fluorescence in situ hybridization [25]. In T. infestans cells, the nucleolus was found to encircle chromocenters (Figure 2a,b) [37]. Additionally, the hypothesis that heterochromatin acts as a gene-silencing compartment has been questioned [30]. Variability relative to the small distance between the edge of the chromocenters and the nuclear periphery raises the suspicion of the mobility of the heterochromatin body with respect to the nuclear envelope [25].
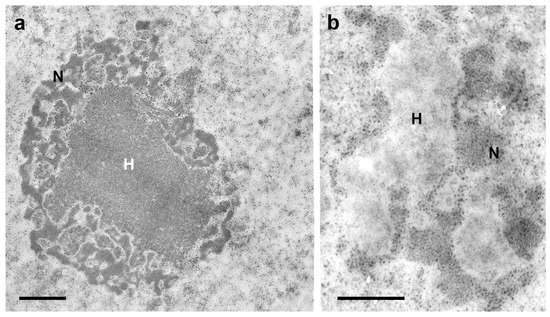
Figure 2. Electron microscopy images of a chromocenter (H)-nucleolus (N) association in Malpighian tubule cell nuclei of fully-nourished fifth instar nymphs of T. infestans (a,b). DNA bleaching in H using Bernard’s EDTA treatment method (b) is compared with an untreated control (a). This method demonstrates electron dense RNA-rich areas (nucleolar components) [37] that surround the electron lucent DNA-rich heterochromatin zones. (M.L.S. Mello and H. Dolder—unpublished images). Bars, 1 µm (a); 0.5 µm (b).
Unraveling of the chromocenter heterochromatin has been observed in a small number of nuclei in the Malpighian tubules of T. infestans under rearing conditions in the laboratory (Figure 1d) [10]. However, the frequency of this phenotype was observed to be significantly increased in fasted nymphs and in specimens subjected to heat shock [10][11]. This phenomenon, which affects the heterochromatin in other cell systems and under different physiological conditions has been hypothesized to represent an attempt to activate silent genes possibly present in this chromatin compartment and achieve cell survival [38][39].
Jagannathan et al. [26][40], through studies in Drosophila and mouse cells, proposed that satellite DNA and their binding proteins may physically integrate the entire chromosome complement into a chromocenter network thus ensuring encapsulation of all chromosomes within a single cell. Notably, images of the epithelial cell nuclei of T. infestans suggest confluence of chromosomes that do not properly integrate the chromocenters into this heterochromatin cluster (Figure 3a–c). This hypothesis can also be suggested when examining published electron microscopy images of T. infestans cell nuclei [37].
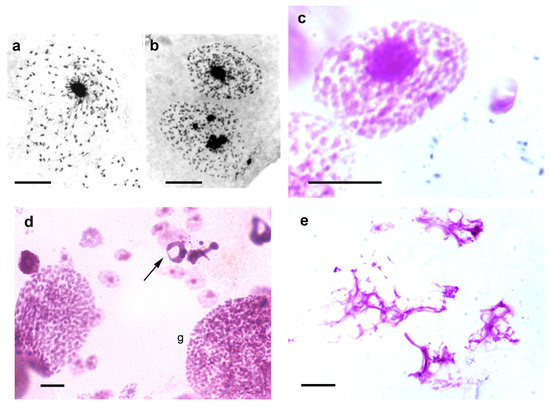
Figure 3. Images suggesting chromosome confluence to chromocenters (a–c), and cell death images (d,e) in Malpighian tubules of T. infestans nymphs. (a,b). Toluidine blue-stained cell nuclei. (c–e). Feulgen-stained cells. Apoptosis ((d), arrow) and necrosis (e) are evident. Both Toluidine blue and Feulgen stainings indicate DNA-containing nuclear elements [9][41]. g, giant nucleus. Bars, 10 µm (a–c,e); 20 µm (d).
References
- Bargues, M.D.; Kiisiowicz, D.R.; Panzera, F.; Noireau, F.; Marcilla, A.; Perez, R.; Rojas, M.G.; O’Connor, J.E.; Gonzales-Candelas, F.; Galvao, C.; et al. Origin and phylogeography of the Chagas disease main vector Triatoma infestans based on nuclear rDNA sequences and genome size. Infect. Genet. Evol. 2006, 6, 436–462.
- Pita, S.; Panzera, F.; Mora, P.; Vel, J.; Cuadrado, A.; Sanchez, A.; Palomeque, T.; Lorite, P. Comparative repeatome analysis on Triatoma infestans Andean and Non-Andean lineages, main vector of Chagas disease. PLoS ONE 2017, 12, e0181635.
- Mello, M.L.S. Somatic polyploidy in insects. Ciênc. Cult. 1970, 22, 348–350.
- Frawley, L.E.; Orr-Weaver, T.L. Polyploidy. Curr. Biol. 2015, 25, R353–R358.
- Anatskaya, O.V.; Vinogradov, A.E. Somatic polyploidy promotes cell function under stress and energy depletion: Evidence from tissue-specific mammal transcriptome. Funct. Integr. Genom. 2010, 10, 433–446.
- Anatskaya, O.V.; Vinogradov, A.E. Polyploidy as a fundamental phenomenon in Evolution, Development, Adaptation and Diseases. Int. J. Mol. Sci. 2022, 23, 3542.
- Mello, M.L.S. Nuclear behavior in the Malpighian tubes of Triatoma infestans (Reduv., Hemiptera). Cytologia 1971, 36, 42–49.
- Mello, M.L.S. Feulgen-DNA values and ploidy degrees in the Malpighian tubes of some triatomids. Rev. Bras. Pesq. Méd. Biol. 1975, 8, 101–107.
- Mello, M.L.S.; Vidal, B.C. The Feulgen reaction: A brief review and new perspectives. Acta Histochem. 2017, 119, 603–609.
- Mello, M.L.S.; Tavares, M.C.H.; Dantas, M.M.; Rodrigues, V.L.C.C.; Maria-Engler, S.S.; Campos, S.P.; Garcia, N.L. Cell death and survival alterations in Malpighian tubules of Triatoma infestans following heat shock. Biochem. Cell Biol. 2001, 79, 709–717.
- Mello, M.L.S. Nuclear fusion and change in chromatin packing state in response to starvation in Triatoma infestans. Rev. Bras. Genét. 1989, 12, 485–498.
- Andrade, C.G.T.J.; Mello, M.L.S. Phenotypes and number of Malpighian tubule nuclei in Triatoma infestans Klug along development and starvation. Rev. Bras. Genét. 1987, 10, 449–457.
- Mello, M.L.S.; Pudney, M. Polyploidization of the BTC-32 cell line from Triatoma infestans (Hemiptera, Reduviidae). Genetica 1987, 74, 131–136.
- Wigglesworth, V.B. Polyploidy and nuclear fusion in the fat body of Rhodnius (Hemiptera). J. Cell Sci. 1967, 2, 603–616.
- Dolder, H.; Mello, M.L.S. Dados preliminares sobre partículas semelhantes a vírus em células de triatomíneos (Preliminary data on virus-like particles in triatomini cells). Rev. Saúde Públ. 1978, 12, 104–109.
- Dolder, H.; Mello, M.L.S. Virus-like particles in the Malpighian tubes of blood-sucking insects. Cell. Mol. Biol. 1978, 23, 299–310.
- Mello, M.L.S. Patterns of lability towards acid hydrolysis in heterochromatin and euchromatin of Triatoma infestans Klug. Cell. Mol. Biol. 1979, 24, 1–16.
- Alvarenga, E.M.; Mondin, M.; Martins, J.A.; Rodrigues, V.L.C.C.; Vidal, B.C.; Mello, M.L.S. Spatial distribution of AT- and GC-rich DNA within interphase cell nuclei of Triatoma infestans Klug. Micron 2011, 42, 568–578.
- Schreiber, G.; Bogliolo, A.R.; Pinto, A.C. Cytogenetics of Triatominae: Karyotype, DNA content, nuclear size and heteropyknosis of autosomes. Braz. J. Biol. 1972, 32, 255–263.
- Solari, J. Autosomal synaptonemal complexes and sex chromosomes without axes in Triatoma infestans (Reduviidae, Hemiptera). Chromosoma 1979, 72, 225–240.
- Morielle-Souza, A.; Azeredo-Oliveira, M.T.V. Differential characterization of holocentric chromosomes in triatomines (Heteroptera, Triatominae) using different staining techniques and fluorescent in situ hybridization. Genet. Mol. Res. 2007, 6, 713–720.
- Hirai, H.; Shomo, Y.; Arias, A.R.; Tada, I. Constitutive heterochromatin polymorphism of a Triatoma infestans strain, a main vector insect of Chagas disease. Jpn. J. Sant. Zool. 1991, 42, 301–303.
- Panzera, F.; Perez, R.; Panzera, Y.; Ferrandis, I.; Ferreiro, M.J.; Calleros, L. Cytogenetics and genome evolution in the subfamily Triatominae (Hemiptera, Reduviidae). Cytogenet. Genome Res. 2010, 128, 77–87.
- Bardella, V.B.; da Rosa, J.A.; Vanzela, A.L.L. Origin and distribution of AT-rich repetitive DNA families in Triatoma infestans (Heteroptera). Infect. Genet. Evol. 2014, 23, 106–114.
- Imperador, C.H.L.; Bardella, V.B.; dos Anjos, E.H.; Rodrigues, V.L.C.C.; Cabral-de-Mello, D.C.; Mello, M.L.S. Spatial distribution of heterochromatin bodies in the nuclei of Triatoma infestans (Klug). Microsc. Microanal. 2020, 26, 567–574.
- Jagannathan, M.; Cummings, R.; Yamashita, Y.M. A conserved function for pericentromeric satellite DNA. eLife 2018, 7, e34122.
- Andrade, C.G.T.J. Fenótipos Nucleares com ênfase na Heterocromatina, e Frequências Nucleares, em Algumas Espécies de Triatomíneos. (Nuclear Phenotypes with Emphasis in Heterochromatin, and Nuclear Frequencies in Some Triatomine Species). Masters’ Thesis, University of Campinas, Campinas, Brazil, 1984.
- Mello, M.L.S.; Vidal, B.C. Critical electrolyte concentration of the heterochromatin and euchromatin of Triatoma infestans. Cytobios 1989, 59, 87–93.
- Mayer, R.; Brero, A.; von Hase, J.; Schroeder, T.; Cremer, T.; Dietzel, S. Common themes and cell type specific variations of higher order chromatin arrangements in the mouse. BMC Cell Biol. 2005, 6, 44.
- Jost, K.L.; Bertulat, B.; Rapp, A.; Brero, A.; Hardt, T.; Domaing, P.; Gösele, C.; Schulz, H.; Hübner, N.; Cardoso, M.C. Gene repositioning within the cell nucleus is not random and is determined by its genomic neighborhood. Epigenetics Chromatin 2015, 8, 36.
- Poulet, A.; Duc, C.; Voisin, M.; Desset, S.; Tutois, S.; Vanrobays, E.; Benoit, M.; Evans, D.E.; Probst, A.V.; Tatout, C. The LINC complex contributes to heterochromatin organization and transcriptional gene silencing in plants. J. Cell Sci. 2017, 130, 590–601.
- Brändle, F.; Frühbauer, B.; Jagannathan, M. Principles and functions of pericentromeric satellite DNA clustering into chromocenters. Semin. Cell Dev. Biol. 2022, 128, 26–39.
- Imperador, C.H.L.; Rodrigues, V.L.C.C.; Mello, M.L.S. The topological distribution of the chromocenter in Panstrongylus megistus (Burmeister) Malpighian tubule cells examined by confocal microscopy. Cytologia 2021, 86, 47–54.
- Allshire, R.C.; Madhani, H.D. Ten principles of heterochromatin formation and function. Nat. Rev. 2018, 19, 229–244.
- Bártova, E.; Krejci, J.; Harnicarová, A.; Galiová, G.; Kozubek, S. Histone modifications and nuclear architecture: A review. J. Histochem. Cytochem. 2008, 56, 711–721.
- Politz, J.C.R.; Scalzo, D.; Groudine, M. Something silent this way forms: The functional organization of the repressive nuclear compartment. Annu. Rev. Cell Dev. Biol. 2013, 29, 241–270.
- Mello, M.L.S.; Dolder, H.; Dias, C.A. Nuclear ultrastructure of Malpighian tubule cells in Triatoma infestans (Hemiptera, Reduviidae) under conditions of full nourishment and starvation. Braz. J. Genet. 1990, 13, 5–17.
- Sandritter, W.; Kiefer, G.; Salm, R.; Moore, G.W.; Grimm, H.; Kiefer, R. DNA in heterochromatin cytophotometric pattern recognition image analysis among cell nuclei in duct epithelium and in carcinoma of the human breast. Beitr. Pathol. 1974, 151, 87–96.
- Simões, L.C.G.; Cestari, A.N. Cromossomos politênicos: Sistemas experimentais in vivo e in vitro. (Polytene chromosomes: In vivo and in vitro experimental systems). Ciênc. Cult. 1982, 34, 480–485.
- Jagannathan, M.; Cummings, R.; Yamashita, Y.M. The modular mechanism of chromocenter formation in Drosophila. eLife 2019, 8, e43938.
- Vidal, B.C.; Mello, M.L.S. Toluidine blue staining for cell and tissue biology applications. Acta Histochem. 2019, 121, 101–112.
More
Information
Subjects:
Cell Biology
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
612
Revisions:
2 times
(View History)
Update Date:
07 Oct 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No