 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Rúben J. Moreira | -- | 4924 | 2023-08-09 21:39:05 | | | |
| 2 | Rúben J. Moreira | -939 word(s) | 3985 | 2023-08-10 15:12:12 | | | | |
| 3 | Peter Tang | + 5 word(s) | 3990 | 2023-08-11 02:53:27 | | |
Video Upload Options
Reduced testosterone (T) levels significantly contribute to male infertility, as this hormone plays a vital role in various functions throughout the male reproductive system. T is produced in the Leydig cells (LC) via testicular steroidogenesis. Dysfunctional LC can impair steroid synthesis and therefore fertility. Endocrine-disrupting chemicals (EDCs) are noteworthy factors influencing steroidogenesis by interfering with hormonal signaling. The heavy metal chromium is classified as an EDC, existing as hexavalent (Cr(VI)) and trivalent chromium (Cr(III)). Despite the debate surrounding Cr(III), chromium picolinate (CrPic3), is used as a nutritional supplement due to its antioxidant and antidiabetic properties. Detrimental effects of CrPic3 on LC encompass enzyme inhibition in steroidogenesis and, while in other cell types, it was found to induce of mutagenesis and apoptosis. Notably, CrPic3 influences male fertility through modifications in reactive oxygen species, T levels, and sperm parameters (e.g., sperm motility and abnormal sperm count). Nonetheless, major gaps and inconsistencies exist in literature concerning its effects on male fertility. Thus, more research is essential to comprehend the mechanisms in which CrPic3 is involved that may be relevant to male fertility, ensuring the safety of this supplement for man.
1. Introduction
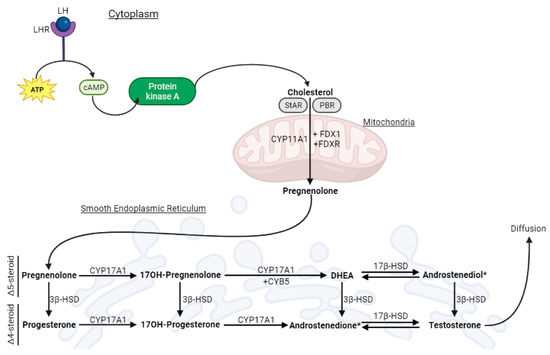
2. Leydig Cells and Testicular Steroidogenesis
3. Chromium: The Good, the Bad, and the Controversial
3.1. Cr(VI): A Toxic Form of Cr
3.2. Cr(III): A Controversial Essential Element
4. Positive and Adverse Effects of Supplementation with CrPic3
4.1. Antidiabetic Effects of CrPic3 and Its Implications on Male Fertility
4.2. Antioxidant Effects of CrPic3 in Cellular Systems
4.3. Possible Mechanisms for CrPic3 Toxicity
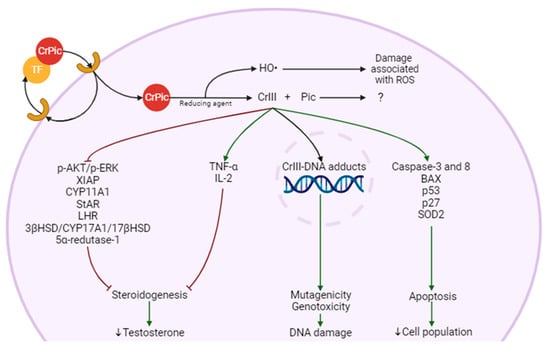
4.4. Impact of CrPic3 on Male Fertility
| Effects | Dose | Duration | Model | Reference |
|---|---|---|---|---|
| ↑ROS, FSH ↓T, HSD, LH Leydig cell damage Spermatid degeneration |
8 and 15 mg/kg body weight | 90 days | Male albino rats | [96] |
| ↑T | 0.2, 0.4, 0.6, 0.8, 1, or 1.2 mg/kg body weight | 84 days | Male Nile tilapia | [56] |
| ↑motility (total and progressive) ↓abnormal sperm |
0.009 mg/kg body weight | 63 days | Male rabbit bucks | [100] |
| Retraction of Leydig cells (artifact?) | 0.250, 0.375, and 0.500 mg/animal/day | 84 days | Santa Inês male lambs | [97] |
| ↑T | 0.8, or 1.2 mg/kg body weight | 84 days | Matrouh cocks | [99] |
| ↓abnormal sperm | 0.08181 mg/kg of food | 95 days | Breeding boar | [101] |
| ↓T ↓ reproductive organs weight ↑testicular Cr levels |
0.020 mg/0.1 kg body weight | 60 days | Wistar male rats | [98] |
Abbreviations: FSH—follicle-stimulating hormone; LH—luteinizing hormone; HSD—hydroxysteroid dehydrogenases; ROS—reactive oxygen species; T—testosterone.
Collectively, these studies suggest that CrPic3 has the potential to influence both LC and sperm cells, yet its precise mechanism of action remains largely unexplored.
5. Conclusions
In summary, Chromium (Cr) falls under the category of EDCs due to its potential to disrupt endocrine signaling. The role of Cr(III) is a subject of ongoing debate, with discussions about whether it should be considered essential or hazardous. Despite this controversy, Cr(III), often in the form of CrPic3, is used as a nutritional supplement, primarily for its antioxidative and antidiabetic properties, among others. Regarding male fertility, CrPic3 seems to impact the expression of a receptor and certain enzymes involved in steroidogenesis within LC, which could potentially lead to a reduction in T levels. Additionally, Cr(III) has been observed to induce changes in the production of inflammatory cytokines, notably TNF-α, exhibit mutagenic and genotoxic effects, and increase apoptosis in various cell types. Should these effects be confirmed in LC, it is plausible that they could interfere with steroid production, ultimately affecting male fertility.
References
- Zegers-Hochschild, F.; Adamson, G.D.; de Mouzon, J.; Ishihara, O.; Mansour, R.; Nygren, K.; Sullivan, E.; Vanderpoel, S.; International Committee for Monitoring Assisted Reproductive Technologies; World Health Organization. International Committee for Monitoring Assisted Reproductive Technology (ICMART) and the World Health Organization (WHO) revised glossary of ART terminology, 2009. Fertil. Steril. 2009, 92, 1520–1524.
- Sharlip, I.D.; Jarow, J.P.; Belker, A.M.; Lipshultz, L.I.; Sigman, M.; Thomas, A.J.; Schlegel, P.N.; Howards, S.S.; Nehra, A.; Damewood, M.D.; et al. Best practice policies for male infertility. Fertil. Steril. 2002, 77, 873–882.
- Agarwal, A.; Mulgund, A.; Hamada, A.; Chyatte, M.R. A unique view on male infertility around the globe. Reprod. Biol. Endocrinol. 2015, 13, 37.
- Ma, Y.; He, X.; Qi, K.; Wang, T.; Qi, Y.; Cui, L.; Wang, F.; Song, M. Effects of environmental contaminants on fertility and reproductive health. Int. J. Environ. Sci. Technol. 2019, 77, 210–217.
- Yilmaz, B.; Terekeci, H.; Sandal, S.; Kelestimur, F. Endocrine disrupting chemicals: Exposure, effects on human health, mechanism of action, models for testing and strategies for prevention. Rev. Endocr. Metab. Disord. 2020, 21, 127–147.
- Badr, F.M.; El-Habit, O. Heavy Metal Toxicity Affecting Fertility and Reproduction of Males. In Bioenvironmental Issues Affecting Men’s Reproductive and Sexual Health; Elsevier: Amsterdam, The Netherlands, 2018; Volume 35, pp. 293–304.
- Concepcion-Zavaleta, M.; Paz Ibarra, J.L.; Ramos-Yataco, A.; Coronado-Arroyo, J.; Concepcion-Urteaga, L.; Roseboom, P.J.; Williams, C.A. Assessment of hormonal status in male infertility. An update. Diabetes Metab. Syndr. Clin. Res. Rev. 2022, 16, 102447.
- Carnegie, C. Diagnosis of Hypogonadism: Clinical Assessments and Laboratory Tests. Rev. Urol. 2004, 6, S3–S8.
- Ohlander, S.J.; Lindgren, M.C.; Lipshultz, L.I. Testosterone and Male Infertility. Urol. Clin. N. Am. 2016, 43, 195–202.
- Maret, W. Chromium Supplementation in Human Health, Metabolic Syndrome, and Diabetes; De Gruyter: Berlin, Germany; Boston, MA, USA, 2019; Volume 19, pp. 231–251.
- Huhtaniemi, I.; Teerds, K. Leydig Cells. In Encyclopedia of Reproduction, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2018; Volume 1, pp. 30–38.
- Chapin, R.E. Reproductive System, Male. In Encyclopedia of Toxicology; Elsevier: Amsterdam, The Netherlands, 2014; Volume 4, pp. 82–92.
- Nassar, G.N.; Leslie, S.W. Physiology, Testosterone; StatPearls Publishing: Treasure Island, FL, USA, 2022.
- Walker, W.H. Molecular mechanisms of testosterone action in spermatogenesis. Steroids 2009, 74, 602–607.
- Flück, C.E.; Pandey, A.V. Testicular Steroidogenesis. In Endocrinology of the Testis and Male Reproduction; Springer: Berlin/Heidelberg, Germany, 2017; pp. 343–371.
- Rone, M.B.; Fan, J.; Papadopoulos, V. Cholesterol transport in steroid biosynthesis: Role of protein–protein interactions and implications in disease states. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2009, 1791, 646–658.
- Haider, S.G. Cell Biology of Leydig Cells in the Testis; Elsevier: Amsterdam, The Netherlands, 2004; Volume 233.
- Hauet, T.; Yao, Z.X.; Bose, H.S.; Wall, C.T.; Han, Z.; Li, W.; Hales, D.B.; Miller, W.L.; Culty, M.; Papadopoulos, V. Peripheral-type benzodiazepine receptor-mediated action of steroidogenic acute regulatory protein on cholesterol entry into leydig cell mitochondria. Mol. Endocrinol. 2005, 19, 540–554.
- Grinspon, R.P.; Bergada, I.; Rey, R.A. Male Hypogonadism and Disorders of Sex Development. Front. Endocrinol. 2020, 11, 211–225.
- Lee, H.G.; Kim, C.J. Classic and backdoor pathways of androgen biosynthesis in human sexual development. Ann. Pediatr. Endocrinol. Metab. 2022, 27, 83–89.
- Grinspon, R.P.; Bergada, I.; Rey, R.A. Male Hypogonadism and Disorders of Sex Development. Front. Endocrinol. 2020, 11, 211–225.
- Simpson, J.L. Disorders of the Gonads, Genital Tract, and Genitalia. In Emery and Rimoin’s Principles and Practice of Medical Genetics; Academic Press: Cambridge, MA, USA, 2013; pp. 1–45.
- Genchi, G.; Lauria, G.; Catalano, A.; Carocci, A.; Sinicropi, M.S. The Double Face of Metals: The Intriguing Case of Chromium. Appl. Sci. 2021, 11, 638.
- Bagchi, D.; Stohs, S.; Downs, B.; Bagchi, M.; Preuss, H. Cytotoxicity and oxidative mechanisms of different forms of chromium. Toxicology 2002, 180, 5–22.
- Pure Earth. World’s Worst Pollution Problems: The New Top Six Toxic Threats: A Priority List for Remediation. 2015. Available online: https://www.worstpolluted.org/docs/WWP15.pdf (accessed on 15 July 2023).
- Quievryn, G.; Messer, J.; Zhitkovich, A. Carcinogenic Chromium(VI) Induces Cross-Linking of Vitamin C to DNA in Vitro and in Human Lung A549 Cells. Biochemistry 2002, 41, 3156–3167.
- Quievryn, G.; Goulart, M.; Messer, J.; Zhitkovich, A. Reduction of Cr (VI) by cysteine Significance in human lymphocytes and formation of DNA damage in reactions with variable reduction rates. Mol. Cell. Biochem. 2001, 222, 107–118.
- Zablon, H.; VonHandorf, A.; Puga, A. Chromium exposure disrupts chromatin architecture upsetting the mechanisms that regulate transcription. Exp. Biol. Med. 2019, 244, 752–757.
- Wu, Y.H.; Lin, J.C.; Wang, T.Y.; Lin, T.J.; Yen, M.C.; Liu, Y.H.; Wu, P.L.; Chen, F.W.; Shih, Y.L.; Yeh, I.J. Hexavalent chromium intoxication induces intrinsic and extrinsic apoptosis in human renal cells. Mol. Med. Rep. 2020, 21, 851–857.
- Handa, K.; Jindal, R. Estimating the hepatotoxic impact of hexavalent chromium on Ctenopharyngodon idellus through a multi-biomarker study. Environ. Adv. 2021, 5, 100108.
- Yatera, K.; Morimoto, Y.; Ueno, S.; Noguchi, S.; Kawaguchi, T.; Tanaka, F.; Suzuki, H.; Higashi, T. Cancer Risks of Hexavalent Chromium in the Respiratory Tract. J. UOEH 2018, 40, 157–172.
- Chandra, A.K.; Chatterjee, A.; Ghosh, R.; Sarkar, M.; Chaube, S.K. Chromium induced testicular impairment in relation to adrenocortical activities in adult albino rats. Reprod. Toxicol. 2007, 24, 388–396.
- Abbas, T.; Khawaja, A.; Qaisar, F.; Sajjad, A. Effects of chromium on testes and protective role of mulberry. J. Sheikh Zayed Med. Coll. 2017, 8, 1200–1204.
- Chandra, A.K.; Chatterjee, A.; Ghosh, R.; Sarkar, M. Vitamin E-supplementation protect chromium (VI)-induced spermatogenic and steroidogenic disorders in testicular tissues of rats. Food Chem. Toxicol. 2010, 48, 972–979.
- Schwarz, K.; Mertz, W. Chromium(III) and the glucose tolerance factor. Arch. Biochem. Biophys. 1959, 85, 292–295.
- McCarty, M. The therapeutic potential of glucose tolerance factor. Med. Hypotheses 1980, 6, 1177–1189.
- Lin, C.C.; Tsweng, G.J.; Lee, C.F.; Chen, B.H.; Huang, Y.L. Magnesium, zinc, and chromium levels in children, adolescents, and young adults with type 1 diabetes. Clin. Nutr. 2016, 35, 880–884.
- Stearns, D.M. Is chromium a trace essential metal? Biofactors 2000, 11, 149–162.
- Vincent, J.B. Chromium: Celebrating 50 years as an essential element? Dalton Trans. 2010, 39, 3787–3794.
- Casalegno, C.; Schifanella, O.; Zennaro, E.; Marroncelli, S.; Briant, R. Collate literature data on toxicity of Chromium (Cr) and Nickel (Ni) in experimental animals and humans. EFSA Support. Publ. 2015, 12, 478E.
- Filippini, T.; Cilloni, S.; Malavolti, M.; Violi, F.; Malagoli, C.; Tesauro, M.; Bottecchi, I.; Ferrari, A.; Vescovi, L.; Vinceti, M. Dietary intake of cadmium, chromium, copper, manganese, selenium and zinc in a Northern Italy community. J. Trace Elem. Med. Biol. 2018, 50, 508–517.
- Witkamp, R. Biologically Active Compounds in Food Products and Their Effects on Obesity and Diabetes. In Chemistry, Molecular Sciences and Chemical Engineering; Elsevier: Amsterdam, The Netherlands, 2010; Volume 3, pp. 509–545.
- Komorowski, J.R.; Tuzcu, M.; Sahin, N.; Juturu, V.; Orhan, C.; Ulas, M.; Sahin, K. Chromium picolinate modulates serotonergic properties and carbohydrate metabolism in a rat model of diabetes. Biol. Trace Elem. Res. 2012, 149, 50–56.
- McLeod, M.; Golden, R. Chromium treatment of depression. Int. J. Neuropsychopharmacol. 2000, 3, 311–314.
- Sahin, K.; Tuzcu, M.; Orhan, C.; Sahin, N.; Kucuk, O.; Ozercan, I.H.; Juturu, V.; Komorowski, J.R. Anti-diabetic activity of chromium picolinate and biotin in rats with type 2 diabetes induced by high-fat diet and streptozotocin. Br. J. Nutr. 2013, 110, 197–205.
- Sundaram, B.; Aggarwal, A.; Sandhir, R. Chromium picolinate attenuates hyperglycemia-induced oxidative stress in streptozotocin-induced diabetic rats. J. Trace Elem. Med. Biol. 2013, 27, 117–121.
- Reed, J.; Bain, S.; Kanamarlapudi, V. A Review of Current Trends with Type 2 Diabetes Epidemiology, Aetiology, Pathogenesis, Treatments and Future Perspectives. Diabetes Metab. Syndr. Obes. Targets Ther. 2021, 14, 3567–3602.
- Martin, J.; Wang, Z.Q.; Zhang, X.H.; Wachtel, D.; Volaufova, J.; Matthews, D.E.; Cefalu, W.T. Chromium Picolinate Supplementation Attenuates Body Weight Gain and Increases Insulin Sensitivity in Subjects with Type 2 Diabetes. Diabetes Care 2006, 29, 1826–1832.
- Dong, F.; Kandadi, M.; Ren, J.; Sreejayan, N. Chromium (D-phenylalanine)3 Supplementation Alters Glucose Disposal, Insulin Signaling, and Glucose Transporter-4 Membrane Translocation in Insulin-Resistant Mice. J. Nutr. 2008, 138, 1846–1851.
- Wang, Y.Q.; Yao, M.H. Effects of chromium picolinate on glucose uptake in insulin-resistant 3T3-L1 adipocytes involve activation of p38 MAPK. J. Nutr. Biochem. 2009, 20, 982–991.
- Chen, W.Y.; Chen, C.J.; Liu, C.H.; Mao, F.C. Chromium supplementation enhances insulin signalling in skeletal muscle of obese KK/HlJ diabetic mice. Diabetes Obes. Metab. 2009, 11, 293–303.
- Jain, S.K.; Rains, J.L.; Croad, J.L. Effect of chromium niacinate and chromium picolinate supplementation on lipid peroxidation, TNF-alpha, IL-6, CRP, glycated hemoglobin, triglycerides, and cholesterol levels in blood of streptozotocin-treated diabetic rats. Free Radic. Biol. Med. 2007, 43, 1124–1131.
- Imanparasta, F.; Javaheric, J.; Kamankesha, F.; Rafieid, F.; Salehie, A.; Mollaaliakbaria, Z.; Rezaeia, F.; Rahimif, A.; Abbasig, E. The effects of chromium and vitamin D3 co-supplementation on insulin resistance and tumor necrosis factor-alpha in type 2 diabetes a randomized placebo-controlled trial. Appl. Physiol. Nutr. Metab. 2020, 45, 471–477.
- Rui, L.; Aguirre, V.; Kim, J.K.; Shulman, G.I.; Lee, A.; Corbould, A.; Dunaif, A.; White, M.F. Insulin/IGF-1 and TNF-alpha stimulate phosphorylation of IRS-1 at inhibitory Ser307 via distinct pathways. J. Clin. Investig. 2001, 107, 181–189.
- Suksomboon, N.; Poolsup, N.; Yuwanakorn, A. Systematic review and meta-analysis of the efficacy and safety of chromium supplementation in diabetes. J. Clin. Pharm. Ther. 2014, 39, 292–306.
- Mehrim, A.I. Physiological, biochemical and histometric responses of Nile tilapia (Oreochromis niloticus L.) by dietary organic chromium (chromium picolinate) supplementation. J. Adv. Res. 2014, 5, 303–310.
- Pattar, G.R.; Tackett, L.; Liu, P.; Elmendorf, J.S. Chromium picolinate positively influences the glucose transporter system via affecting cholesterol homeostasis in adipocytes cultured under hyperglycemic diabetic conditions. Mutat. Res. 2006, 610, 93–100.
- Shahinfar, H.; Amini, M.R.; Sheikhhossein, F.; Djafari, F.; Jafari, A.; Shab-Bidar, S. The effect of chromium supplementation on apolipoproteins: A systematic review and meta-analysis of randomized clinical trials. Clin. Nutr. ESPEN 2020, 40, 34–41.
- Dominiczak, M.H.; Caslake, M.J. Apolipoproteins: Metabolic role and clinical biochemistry applications. Ann. Clin. Biochem. 2011, 48, 498–515.
- Zhong, O.; Ji, L.; Wang, J.; Lei, X.; Huang, H. Association of diabetes and obesity with sperm parameters and testosterone levels: A meta-analysis. Diabetol. Metab. Syndr. 2021, 13, 109.
- Alves, M.G.; Oliveira, P. Diabetes Mellitus and male reproductive function: Where we stand? Int. J. Diabetol. Vasc. Dis. Res. 2023, 1, 1–2.
- Meneses, M.J.; Sousa, M.; Alves, M.; Oliveira, P. The Antidiabetic Drug Metformin and Male Reproductive Function: An Overview. Int. J. Diabetol. Vasc. Dis. Res. 2015, 1, 1–2.
- Vona, R.; Pallotta, L.; Cappelletti, M.; Severi, C.; Matarrese, P. The Impact of Oxidative Stress in Human Pathology: Focus on Gastrointestinal Disorders. Antioxidants 2021, 10, 201.
- Darbandi, M.; Darbandi, S.; Agarwal, A.; Sengupta, P.; Durairajanayagam, D.; Henkel, R.; Sadeghi, M.R. Reactive oxygen species and male reproductive hormones. Reprod. Biol. Endocrinol. 2018, 16, 87.
- Rato, L.; Oliveira, P.F.; Sousa, M.; Silva, B.M.; Alves, M.G. Chapter 2.6—Role of Reactive Oxygen Species in Diabetes-Induced Male Reproductive Dysfunction. In Oxidants, Antioxidants and Impact of the Oxidative Status in Male Reproduction; Henkel, R., Samanta, L., Agarwal, A., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 135–147.
- Doddigarla, Z.; Parwez, I.; Abidi, S.; Ahmad, J. Effect of Chromium Picolinate and Melatonin either in Single or in a Combination in Alloxan Induced Male Wistar Rats. J. Biomed. Sci. 2017, 6.
- Al-Bishri, W.M. Attenuating impacts of chromium and nano resveratrol against hyperglycemia induced oxidative stress in diabetic rats. Int. J. Pharm. Res. Allied Sci. 2017, 6, 61–69.
- Kolahian, S.; Sadri, H.; Shahbazfar, A.A.; Amani, M.; Mazadeh, A.; Mirani, M. The Effects of Leucine, Zinc, and Chromium Supplements on Inflammatory Events of the Respiratory System in Type 2 Diabetic Rats. PLoS ONE 2015, 10, e0133374.
- Jain, S.K.; Kahlon, G.; Morehead, L.; Dhawan, R.; Lieblong, B.; Stapleton, T.; Caldito, G.; Hoeldtke, R.; Levine, S.N.; Bass III, P.F. Effect of chromium dinicocysteinate supplementation on circulating levels of insulin, TNF-α, oxidative stress, and insulin resistance in type 2 diabetic subjects: Randomized, double-blind, placebo-controlled study. Mol. Nutr. Food Res. 2012, 56, 1333–1341.
- Saiyed, Z.M.; Lugo, J.P. Impact of chromium dinicocysteinate supplementation on inflammation, oxidative stress, and insulin resistance in type 2 diabetic subjects: An exploratory analysis of a randomized, double-blind, placebo-controlled study. Food Nutr. Res. 2016, 60, 31762.
- Clodfelder, B.J.; Vincent, J.B. The time-dependent transport of chromium in adult rats from the bloodstream to the urine. J. Biol. Inorg. Chem. 2005, 10, 383–393.
- Sun, Y.; Ramirez, J.; Woski, S.A.; Vincent, J.B. The binding of trivalent chromium to low-molecular-weight chromium-binding substance (LMWCr) and the transfer of chromium from transferrin and chromium picolinate to LMWCr. JBIC J. Biol. Inorg. Chem. 2000, 5, 129–136.
- Manygoats, K.R.; Yazzie, M.; Stearns, D.M. Ultrastructural damage in chromium picolinate-treated cells: A TEM study. Transmission electron microscopy. J. Biol. Inorg. Chem. 2002, 7, 791–798.
- Navin, A.K.; Aruldhas, M.M.; Navaneethabalakrishnan, S.; Mani, K.; Michael, F.M.; Srinivasan, N.; Banu, S.K. Prenatal exposure to hexavalent chromium disrupts testicular steroidogenic pathway in peripubertal F(1) rats. Reprod. Toxicol. 2021, 101, 63–73.
- Dworzanski, W.; Sembratowicz, I.; Cholewinska, E.; Tutaj, K.; Fotschki, B.; Juskiewicz, J.; Ognik, K. Effects of Different Chromium Compounds on Hematology and Inflammatory Cytokines in Rats Fed High-Fat Diet. Front. Immunol. 2021, 12, 614000.
- Leisegang, K.; Henkel, R. The in vitro modulation of steroidogenesis by inflammatory cytokines and insulin in TM3 Leydig cells. Reprod. Biol. Endocrinol. 2018, 16, 26.
- Suescun, M.O.; Rival, C.; Theas, M.S.; Calandra, R.S.; Lustig, L. Involvement of tumor necrosis factor-α in the pathogenesis of autoimmune orchitis in rats. Biol. Reprod. 2003, 68, 2114–2121.
- Hales, D.B. Testicular macrophage modulation of Leydig cell steroidogenesis. J. Reprod. Immunol. 2002, 57, 3–18.
- Wang, Y.; Chen, L.; Xie, L.; Li, L.; Li, X.; Li, H.; Liu, J.; Chen, X.; Mao, B.; Song, T. Interleukin 6 inhibits the differentiation of rat stem Leydig cells. Mol. Cell. Endocrinol. 2018, 472, 26–39.
- Blankert, S.A.; Coryell, V.H.; Picard, B.T.; Wolf, K.K.; Lomas, R.E.; Stearns, D.M. Characterization of Nonmutagenic Cr(III)-DNA Interactions. Chem. Res. Toxicol. 2003, 16, 847–854.
- Brown, S.; Lockart, M.M.; Thomas, C.S.; Bowman, M.K.; Woski, S.A.; Vincent, J.B. Molecular Structure of Binary Chromium(III)-DNA Adducts. ChemBioChem 2020, 21, 628–631.
- Lankford, E.; Thomas, C.S.; Marchi, S.; Brown, S.; Woski, S.A.; Vincent, J.B. Examining the Potential Formation of Ternary Chromium-Histidine-DNA Complexes and Implications for Their Carcinogenicity. Biol. Trace Elem. Res. 2022, 200, 1473–1481.
- Xu, G.; Shi, Y. Apoptosis signaling pathways and lymphocyte homeostasis. Cell Res. 2007, 17, 759–771.
- Jana, M.; Rajaram, A.; Rajaram, R. Chromium picolinate induced apoptosis of lymphocytes and the signaling mechanisms thereof. Toxicol. Appl. Pharmacol. 2009, 237, 331–344.
- Dębski, B.; Lamparska-Przybysz, M.; Gajewska, M. Influence of Cr(III)-picolinate, and Cr(III)-nicotinate on apoptosis induction in HBL-100 human mammary epithelial cells. Probl. Hig. Epidemiol. 2016, 97, 95–99.
- Sivakumar, K.K.; Stanley, J.A.; Arosh, J.A.; Pepling, M.E.; Burghardt, R.C.; Banu, S.K. Prenatal exposure to chromium induces early reproductive senescence by increasing germ cell apoptosis and advancing germ cell cyst breakdown in the F1 offspring. Dev. Biol. 2014, 388, 22–34.
- Shen, Y.; White, E. p53-Dependent Apoptosis Pathways. Adv. Cancer Res. 2001, 82, 55–84.
- Abbastabar, M.; Kheyrollah, M.; Azizian, K.; Bagherlou, N.; Tehrani, S.S.; Maniati, M.; Karimian, A. Multiple functions of p27 in cell cycle, apoptosis, epigenetic modification and transcriptional regulation for the control of cell growth: A double-edged sword protein. DNA Repair 2018, 69, 63–72.
- Pena-Blanco, A.; Garcia-Saez, A.J. Bax, Bak and beyond–mitochondrial performance in apoptosis. FEBS J. 2018, 285, 416–431.
- Li, P.; Nijhawan, D.; Budihardjo, I.; Srinivasula, S.M.; Ahmad, M.; Alnemri, E.S.; Wang, X. Cytochrome c and dATP-Dependent Formation of Apaf-1/Caspase-9 Complex Initiates an Apoptotic Protease Cascade. Cell 1997, 91, 479–489.
- Kim, A.H.; Khursigara, G.; Sun, X.; Franke, T.F.; Chao, M.V. Akt phosphorylates and negatively regulates apoptosis signal-regulating kinase 1. Mol. Cell. Biol. 2001, 21, 893–901.
- Chang, F.; Lee, J.T.; Navolanic, P.M.; Steelman, L.S.; Shelton, J.G.; Blalock, W.L.; Franklin, R.A.; McCubrey, J.A. Involvement of PI3K/Akt pathway in cell cycle progression, apoptosis, and neoplastic transformation: A target for cancer chemotherapy. Leukemia 2003, 17, 590–603.
- Franke, T.F.; Hornik, C.P.; Segev, L.; Shostak, G.A.; Sugimoto, C. PI3K/Akt and apoptosis: Size matters. Oncogene 2003, 22, 8983–8998.
- Gardai, S.J.; Whitlock, B.B.; Xiao, Y.Q.; Bratton, D.B.; Henson, P.M. Oxidants inhibit ERK/MAPK and prevent its ability to delay neutrophil apoptosis downstream of mitochondrial changes and at the level of XIAP. J. Biol. Chem. 2004, 279, 44695–44703.
- Tsa, M.C.; Lien, T.F. Chromium Picolinate did not Effect on the Proliferation and Differentiation of Myoblasts. Am. J. Anim. Vet. Sci. 2007, 2, 79–83.
- Rehab, M.E.; Ashraf, M.E. The protective effect of Panax ginseng against chromium picolonate induced testicular changes. Afr. J. Pharm. Pharmacol. 2014, 8, 346–355.
- Dallago, B.S.L.; Braz, S.; Marçola, T.G.; McManus, C.; Caldeira, D.F.; Campeche, A.; Gomes, E.F.; Paim, T.P.; Borges, B.O.; Louvandini, H. Blood Parameters and Toxicity of Chromium Picolinate Oral Supplementation in Lambs. Biol. Trace Elem. Res. 2015, 168, 91–102.
- Zakaria, A.D.; Fayed, A.H.; Hedaya, S.A.; Gad, S.B.; Hafez, M.H. Effect of chromium Picolinate on some reproductive aspects in male rats. Alex. J. Vet. Sci. 2011, 34, 113–124.
- Ezzat, W. Effect of supplementing diet with sodium bentonite and/or organic chromium on productive, physiological performance and immune response of matrouh chickens strain. 1- during growth period. Egypt. Poult. Sci. J. 2016, 36, 841–857.
- Dorra, T.; El-Serwy, A.; Ismail, F.; Nasif, A. Physical semen characteristics of california rabbit bucks administrated with different chromium levels. J. Anim. Poult. Prod. 2007, 32, 8931–8941.
- Horký, P.; Jančíková, P.; Zeman, L. The effect of a supplement of chromium (picolinate) on the level of blood glucose, insulin activity and changes in laboratory evaluation of the ejaculate of breeding boars. Acta Univ. Agric. Silvic. Mendel. Brun. 2013, 60, 49–56.
- Walker, W.H. Non-classical actions of testosterone and spermatogenesis. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 1557–1569.

