Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Michel Popoff | -- | 4956 | 2023-08-04 18:25:06 | | | |
| 2 | Camila Xu | Meta information modification | 4956 | 2023-08-07 03:31:44 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Popoff, M.R.; Faure, G.; Legout, S.; Ladant, D. Animal Toxin Historical Outlook from Paris Institut Pasteur. Encyclopedia. Available online: https://encyclopedia.pub/entry/47670 (accessed on 24 June 2026).
Popoff MR, Faure G, Legout S, Ladant D. Animal Toxin Historical Outlook from Paris Institut Pasteur. Encyclopedia. Available at: https://encyclopedia.pub/entry/47670. Accessed June 24, 2026.
Popoff, Michel R., Grazyna Faure, Sandra Legout, Daniel Ladant. "Animal Toxin Historical Outlook from Paris Institut Pasteur" Encyclopedia, https://encyclopedia.pub/entry/47670 (accessed June 24, 2026).
Popoff, M.R., Faure, G., Legout, S., & Ladant, D. (2023, August 04). Animal Toxin Historical Outlook from Paris Institut Pasteur. In Encyclopedia. https://encyclopedia.pub/entry/47670
Popoff, Michel R., et al. "Animal Toxin Historical Outlook from Paris Institut Pasteur." Encyclopedia. Web. 04 August, 2023.
Copy Citation
The term “toxin” is derived from the Greek word “toxon”, which refers to a bow, implying poisoned arrows, while “poison” is a broader term originating from the Latin “potionem” (meaning a drink). Initially, it denoted harmful liquid substances, and later, it encompassed any dangerous substances. The earliest civilizations were familiar with animal poisons, often using them in combination with plant toxins for hunting and fishing. They also recognized toxins for their potential therapeutic or psychoactive effects, capable of inciting fury, trances, love, and ecstasy.
Pasteur
animal toxin
venom
vaccine
serotherapy
therapeutic peptides
phospholipase A2
1. Introduction
Toxins and poisonings have been known since the most ancient times. During the Paleolithic era in Europe, hunters used poisons. Indeed, paleontologists have discovered unusual grooves on the tips of certain bones used as arrows, suggesting they contained plant or animal poisons. The use of poisons, predominantly from a plant origin, on arrowheads appears to have been widespread among various civilizations across all continents for hunting purposes [1]. The term “toxin” is derived from the Greek word “toxon”, which refers to a bow, implying poisoned arrows, while “poison” is a broader term originating from the Latin “potionem” (meaning a drink). Initially, it denoted harmful liquid substances, and later, it encompassed any dangerous substances. The earliest civilizations were familiar with animal poisons, often using them in combination with plant toxins for hunting and fishing. They also recognized toxins for their potential therapeutic or psychoactive effects, capable of inciting fury, trances, love, and ecstasy. Ancient medical texts such as the Indian Vedic books from 1500 BC demonstrate a considerable knowledge of poisonous substances, including animal poisons, during this era [1]. The term “venom” originates from the Latin name Venus, the goddess of love and beauty. Initially, the word was used to indicate a love potion. Later, it adopted a more ambiguous meaning as a remedy, psychoactive drug, or poison [2]. Nowadays, venom refers specifically to animal poisons delivered through biting. There is a remarkable diversity of venomous animals, including approximately 1450 species of fish, 1200 species of marine organisms, 400 snakes, 200 spiders, 75 scorpions, and 60 ticks [1].
2. Pasteur’s Era: The Discovery of Antivenom Serotherapy
Louis Pasteur was aware of the work on bacterial toxins conducted by Peter Ludvig Panum and von Bergman, who focused on toxins produced by putrefying bacteria [3]. However, L. Pasteur was primarily interested in preventing infectious diseases with attenuated microorganisms. For L. Pasteur, the main mechanism of pathogenicity was the development of microbes in the host, causing a depletion of vital substances [3].
In 1888, Cesaire Phisalix (1852–1906), a French military physician and scientist, joined the National Museum of Natural History (MNHN, Museum National d’Histoire Naturelle) in Paris in the laboratory of comparative pathology directed by Auguste Chauveau. The MNHN was created in 1636 as the royal garden for the preservation and training of medicinal plants. It was formally established as the National Museum of Natural History in 1793 during the French Revolution. Its activity extends to research, collection, and training in various fields such as botany, chemistry, mineralogy, and zoology, including the study of venomous animals. C. Phisalix began investigating salamander venom and later collaborated with Gabriel Bertrand on viper venoms. Between 1889 and 1891, C. Phisalix published five articles in which he described the neurotoxic, respiratory, and thermal effects of salamander poison on experimental animals, as well as the possibility of inducing immunity through successive injections of low doses of the poison. G. Bertrand (1867–1962), a French chemist and biologist, entered the MNHN in 1886. He initially served as an assistant in a laboratory of plant physiology applied to agriculture (1889–1890), and subsequently in a laboratory of chemistry applied to organic substances (1890). A strong friendship developed between C. Phisalix and G. Bertrand (Figure 1). G. Bertrand later joined the Institut Pasteur in 1900, where he developed biological chemistry. Encouraged by A. Chauveau, C. Phisalix began studying venoms with the idea that venoms are possibly similar to bacterial toxins. He hypothesized that the methods of toxigenic bacteria attenuation for vaccination purposes could be applied to venoms. Indeed, following the success of L. Pasteur in preventing fowl cholera (1880), anthrax in sheep (1881), and rabies (1885) with the attenuated bacteria or viruses responsible for these diseases, C. Phisalix discovered a way to inactivate venom by heating it at 80 °C for 5 min and subsequently using it to vaccinate guinea pigs. In 1894, C. Phisalix and G. Bertrand presented their work at the Society of Biology in Paris on the immunization of guinea pigs with heat-attenuated venoms, and the subsequent treatment of venom-intoxicated animals with the serum of immunized guinea pigs. This marked the beginning of antivenomous serotherapy [4][5]. Moreover, C. Phisalix and G. Bertrand discovered that, unlike salamanders, vipers contain substances in their blood that confer resistance to their own venom. They also identified a natural resistance to viper venom in two species of grass snakes. Venom inhibitors in the blood of numerous snakes and mammals were later characterized (see below).

Figure 1. Cesaire Phisalix and Gabriel Bertrand. (A) C. Phisalix (1852–1906); photo was taken by Marie Phisalix, Musée d’Histoire Naturelle de Moutiers–Haute–Pierre. (B) G. Bertrand (1867–1962) around 1905; photo was provided by the Institut Pasteur/Musée Pasteur.
Albert Calmette (1863–1933) (Figure 2), a French military physician, followed the course of “microbie technique” directed by Emile Roux at the Institut Pasteur, Paris, in 1890. The next year, E. Roux sent A. Calmette in Saigon to establish the first Institut Pasteur network site, initially dedicated to the preparation of anti-rabies and anti-smallpox vaccines. During his stay in Indochina, he had the opportunity to investigate snake venoms. Back to France in 1893, he was sent to Lille to install a second Institut Pasteur (Institut Pasteur Lille), which he directed from 1895 to 1919. He tried to develop an immunization against cobra venom by adapting techniques used for bacterial toxins. However, cobra venom is resistant to heating. He then tried inoculating escalating doses, starting from sublethal doses, as well as alternative ways to inactivate snake venom by using various chemical treatments. A. Calmette succeeded in obtaining complete immunization using cobra venom treated with sodium hypochlorite, and he also demonstrated that sera from immunized animals have preventive and therapeutic effects against venom intoxication. He presented his results on antivenomous therapy in 1894 at the same session of the Society of Biology as C. Phisalix and G. Bertrand. A rivalry between the two scientist groups emerged for several years. The French National Academy of Sciences awarded the Monthyon prize to C. Phisalix and G. Bertrand for the discovery of an antivenom serum in 1894. However, A. Calmette claimed that his protocol for venom attenuation was more efficient than that of C. Phisalix. Indeed, snake venoms are more or less thermostable, and the chemical attenuation was more effective. Unfortunately, C. Phisalix died prematurely at age 54, and A. Calmette emphasized the importance of the antivenom serotherapy and his results at the international level. A. Calmette published numerous articles in French, English, and German, and he presented the interest and efficiency of the antivenom serotherapy at the Royal College of Physicians and Surgeons of London in 1896. It was found later that Calmette’s protocol of immunization induced highly neutralizing specific IgG antibodies, whereas those of C. Phisalix and G. Bertrand triggered mainly IgM. A. Calmette thought that the sera obtained after immunization with one snake venom could protect against the venom from different snake species. He produced antisera against cobra venom on a large scale at the Institut Pasteur of Lille for therapeutic use in humans. Then, the production of antivenom sera was undertaken in various countries: Brazil (Butantan Institute, 1901), the USA (Philadelphia, 1902), Australia (Sydney, 1902), India (Hafkine Institute of Mumbai, 1903; Kasauli, 1907), South Africa (Johannesburg, 1903), and the UK (London, 1905) [4][5][6][7][8][9].

Figure 2. Albert Calmette (1863–1933) in his laboratory around 1920–1925. Photo was taken by Henri Manuel, Institut Pasteur/Musée Pasteur.
3. Period of 1923–1978: Development of Antivenom Sera and Initial Characterization of Venom Constituents
Gaston Ramon (1886–1963) (Figure 3), a French veterinarian and biologist, was recruited by E. Roux to produce horse antisera at the Institut Pasteur Garches Annex. The property of Garches, in the Paris suburb of Marnes la Coquette, was gifted to L. Pasteur by the government in 1884 for his studies on rabies. This property, which was part of an old castle (castle of Villeneuve l’Etang), sufficiently distant from residential areas, was more favorable for maintaining a kennel of rabid dogs than Pasteur’s laboratory at the Ecole Normale Supérieure in downtown Paris. The Institut Pasteur in Paris was founded through a public international subscription and was inaugurated in 1888. It was dedicated to the treatment of rabies, basic research into infectious diseases, and training on microorganisms (with the first “microbie technique” course taught by E. Roux in 1888). After Pasteur’s death in 1895, the Garches Annex was used for the production of antisera in horses and investigations into animal immunization (Figure 4). Louis Martin (1864–1946), a French physician and biologist, was the assistant of E. Roux (1893–1894) and contributed to the development of anti-diphtheria serotherapy. L. Martin was appointed as the deputy head of the laboratory responsible for the production of an anti-diphtheria toxin and, subsequently, an anti-tetanus toxin (1894–1909).

Figure 3. Gaston Ramon (1886–1933) in his laboratory around 1925. Photo was provided by the Institut Pasteur/Musée Pasteur.
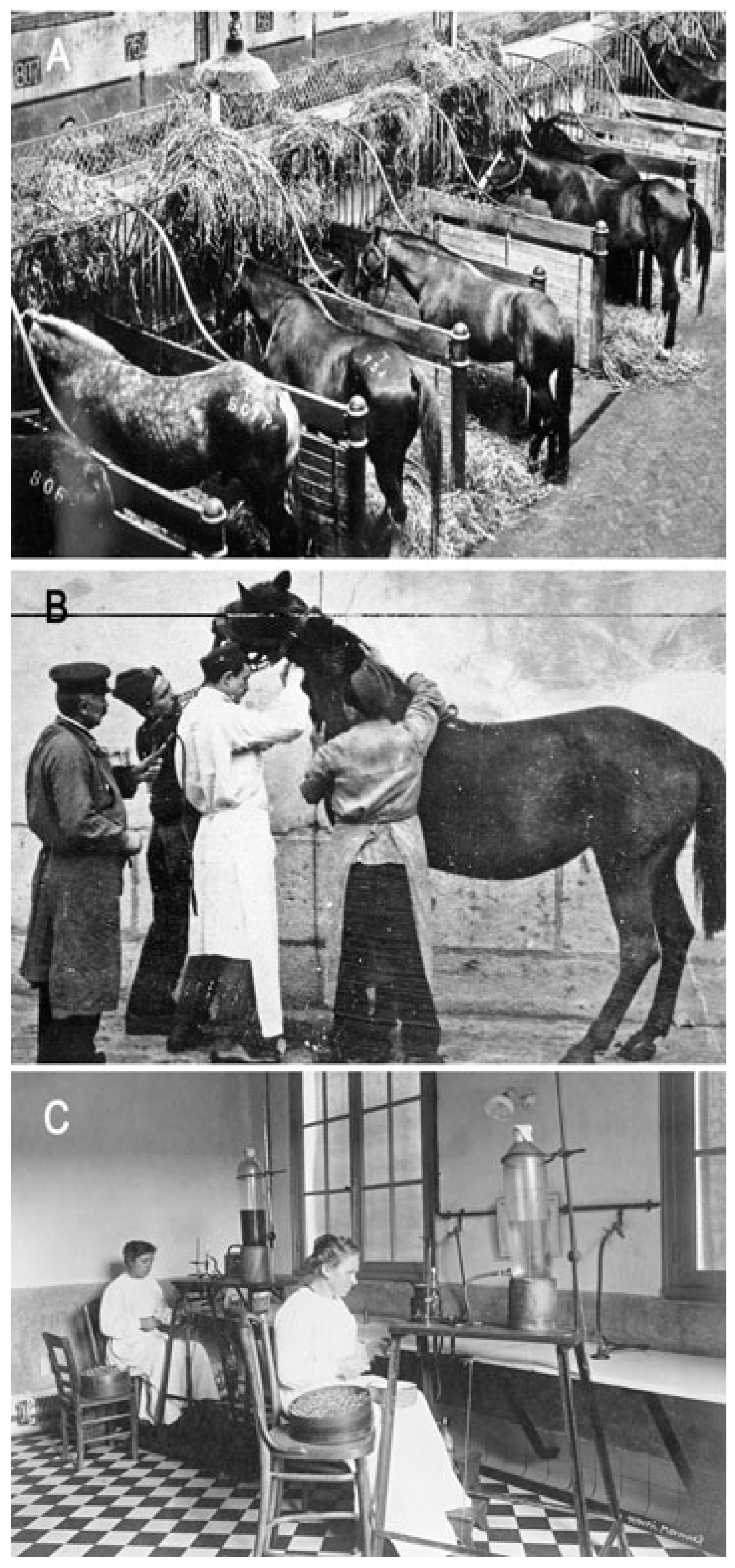
Figure 4. Institut Pasteur of Garches, production of horse antisera. (A) Horse stables; photo was provided by the Institut Pasteur/Musée Pasteur. (B) Serum sampling in a horse; photo was provided by the Institut Pasteur/Musée Pasteur. (C) Serum bottling; photo was taken by Henri Manuel, Institut Pasteur/Musée Pasteur.
G. Ramon was assigned to the serum production laboratory at the Garches Annex by E. Roux from 1911 to 1920 under the supervision of André-Romain Prévot [10]. In 1923, G. Ramon developed an innovative method of toxin inactivation based on a treatment with formalin. Thus, he demonstrated that the diphtheria toxin treated with a low dose of formalin became inactive, yet was able to induce a potent immunological response. He termed this novel form of toxin an “anatoxin” [3]. The idea to use formalin came from the fact that he applied this compound as an antiseptic for the preservation of therapeutic sera. He also used formalin for preserving standardized toxin samples, and he noticed that toxins treated with formalin were harmless, stable, and immunogenic [11]. This inactivation process was applied to other bacterial toxins, such as the tetanus toxin, as well as to venom. Indeed, in 1924, G. Ramon showed that snake venom could be inactivated by formalin to create a product that he referred as an “anavenom”. This method, which proved more reliable than a heat treatment or other chemical inactivation methods, was used for the immunization of horses and the production of antivenomous sera. Furthermore, G. Ramon introduced the concept of immunity adjuvants. He showed that the induction of local inflammation with calcium chloride or alum (aluminum hydroxide) could enhance the immune response to anatoxins. From 1926 to 1944, G. Ramon was the director of the Garches Annex, where he coordinated the production of anatoxins and antisera [12][13].
Paul Boquet, a French physician and scientist, joined the Institut Pasteur Garches Annex in 1933. He directed a laboratory specializing in antivenomous serotherapy, which was renamed the Laboratory of Venoms and Antivenomous Sera, and he held this position until 1978. P. Boquet conducted extensive research on the composition of snake venoms, notably from various species of Vipers and Naja. He developed novel in vivo and in vitro methods to discern the different toxic properties and enzymatic activities of venoms. The separation of venom constituents was achieved through electrophoresis and liquid chromatography. P. Boquet demonstrated through immunoelectrophoresis that certain toxic constituents are present in the venoms of multiple snake species, facilitating the cross-neutralization of snake venoms with antivenomous sera from different snake species. A key objective was to characterize the unique properties of venom constituents and to select the most effective antigens for creating polyvalent sera through horse immunizations. He proposed the classification of snake venoms into three groups based on their molecular size and serological properties. P. Boquet characterized the major lethal factor of the Naja snake’s venom, which is a small basic peptide of 61 amino acids called an alpha toxin, and he showed that it is produced by various Elapidae snakes. In collaboration with France Tazieff-Depierre, P. Boquet discovered that this factor exhibits activity similar to that of curare. Furthermore, the tritiated alpha toxin from the Naja snake (prepared by André Menez, CEA, France) was used to isolate the cholinergic receptor of the electric organ of Electrophorus by Jean Pierre Changeux, Institut Pasteur. P. Boquet also investigated the coagulation activity of venoms and phospholipases (PLAs), hypothesizing that PLAs might facilitate the entry of toxins into the nervous system. Moreover, P. Boquet supervised the production and control of antivenomous sera, which the Institut Pasteur distributed to numerous countries, notably Africa and Asia. In 1966, P. Boquet reported that 143 horses were used to produce 4345 L of antivenomous sera, leading to the preparation of 246,176 therapeutic doses of 10 mL each. He also contributed to the development and validation of international methods of antivenomous serum titration as well as the preparation of reference standards of venoms and sera in cooperation with the World Health Organization (WHO) [14][15][16][17][18][19][20][21][22][23][24][25][26][27].
France Tazieff-Depierre (1914–2006), a French pharmacist, entered the Institut Pasteur in 1934 in the Unit of Therapeutic Chemistry and then in the Unit of Protein Chemistry until 1979. Initially, she was interested in curararizing compounds and in the role of Ca++ in neurotransmitter release at the neuromuscular junction. She identified several classes of curararizing compounds, including those preventing acetylcholine activity and those with a depolarizing activity. These studies were completed by an investigation into the antagonists of curararizing compounds, such as acetylcholinesterases. F. Tazieff-Depierre showed that Ca++ activates acetylcholinesterases in vitro, but prevents their activity when fixed to the acetylcholine receptor. Starting in 1966, she was involved in animal toxins. P. Boquet asked her to analyze the mechanism of the paralytic effects of cobra venom. F. Tazieff-Depierre showed that the alpha toxin from Naja nigricollis binds with a high affinity to the acetylcholine receptor and that the paralytic effects are antagonized by anti-acetylcholinesterases, which increase the acetylcholine levels. In contrast, the gamma toxin from Naja nigricollis is cardiotoxic and induces muscle paralysis by the excessive Ca++ release from muscle fibers. The toxins from scorpion venom share a similar activity on striated muscle. Moreover, toxins from scorpions and sea anemones enhance the release of acetylcholine from neuronal endings through an increased intra-terminal Na+ concentration and the possible subsequent Ca++ release from internal stores, which triggers neurotransmitter release. Interestingly, due to their effect on acetylcholine release, these toxins are able to restore the neuromuscular transmission inhibited by botulinum toxin A. The numerous and pertinent works of F. Tazieff-Depierre shed light on the mode of action of some venom toxins, and also on the molecular mechanism of neurotransmission, notably on the role of Ca++ [28][29][30][31][32][33][34][35][36][37][38][39][40].
Several other scientists at the Institut Pasteur were also involved in venom research during this period. Camille Delezenne (1868–1932), a French physician, was recruited by Emile Duclaux to manage a physiology laboratory at the Institut Pasteur in 1900. He showed that the hemolytic and coagulase properties of certain snake venoms resulted from specific zinc-dependent enzymatic activities [41]. Marcel Rouvier (1898–1981) joined the Institut Pasteur in 1940 and successively directed the laboratories of the tetanus toxin, the diphtheria toxin, and the “Service des Anaérobies” from 1956 to 1968. Among his works on toxins, he studied the antigenicity of certain snake venoms, such as those from Vipera aspis. He also investigated various methods of preserving antivenomous sera for optimal therapeutic use, despite drastic field constraints such as exposure to heat [42].
4. Period of 1972–2004: Optimization of Serotherapy, Characterization of Biological Activity of Venom Components, and Natural Inhibitors from Snake Blood
Cassian Bon (1944–2008) (Figure 5) was born in Vietnam and completed his studies at the Ecole Normale Supérieure in Paris. In 1972, he joined the Institut Pasteur in Paris and worked on scorpion venoms with F. Tazieff-Depierre. This experience proved decisive, setting the course of his career on the study of venoms, primarily snake venoms, and antivenom sera across his dual scientific roles at the CNRS (Centre National de la Recherche Scientifique) and the Institut Pasteur [43]. C. Bon defended his PhD in 1979 on the major neurotoxins in snake venoms, ceruleotoxin and crototoxin. He later became a research scientist at the CNRS and led a venom laboratory at the Institut Pasteur. This laboratory was part of the cellular pharmacology unit directed by Bernardo-Boris Vargaftig (1937-), a Brazilian-born scientist who completed his medical studies in Sao Paulo (1963) and his scientific studies in Paris (PhD 1972). The research unit of B. Vargaftig evolved into the unit of “Pharmacology of the Mediators of Inflammation and Thrombosis” (1985–1997), and then the unit of “Biology, Cellular and Molecular Pharmacology of Pulmonary Inflammation” (1998–2004). B. Vargaftig is well known for his research on the role of the platelet-activating factor, particularly in bronchoconstriction and platelet aggregation.

Figure 5. Cassian Bon (1944–2008) in his laboratory in 1995. Photo was provided by the Institut Pasteur.
In 1990, C. Bon was appointed as the director of the “Unité des Venins” at the Institut Pasteur, a role he held until 2004 [43]. His research focused on the mechanism of the presynaptic neurotoxicity of Viperidae toxins in snake venom, and he later developed studies on antivenom immunotherapy, both in experimental and human cases. This line of inquiry followed the Pasteur tradition initiated by Albert Calmette in 1894. C. Bon, along with his collaborators, studied the functional chaperon role and synergistic action of the acidic subunit of crotoxin. Crotoxin is a heterodimeric β-neurotoxin from the venom of the South American rattlesnake Crotalus durissus terrificus. The venom was purchased from the Instituto Butantan (Sao-Paulo, SP, Brazil) and crotoxin was purified from the venom using low-pressure gel filtration and ion-exchange chromatography. It is a toxic protein formed by the non-covalent association of a basic PLA2 subunit of low toxicity (CB) and a nontoxic acidic subunit (CA) devoid of catalytic activity that potentiates the toxic effect of CB [44]. Crotoxin binds to the presynaptic membrane, inducing the complete failure of neuromuscular transmission by impairing the release of acetylcholine at the neuromuscular junctions [45]. The CA subunit enhances the capacity of CB to reach its target at the neuromuscular junction, thereby increasing its lethal potency, but it reduces the enzymatic activity of CB [46][47][48][49][50].
Grazyna Faure, a Pasteurian collaborator of C. Bon, discovered several isoforms of crotoxin in individual venom samples of Crotalus durissus terrificus [51]. She purified 16 natural isoforms of crotoxin and compared their molecular structures and biological activities [52]. The work of G. Faure and C. Bon demonstrated that crotoxin variants result from the random association of four isoforms of the CA subunit (CA1–4) and four isoforms of the CB subunit (CBa2, CBb, CBc, and CBd). They described two classes of crotoxin, Class I and Class II, which differ in their pharmacological properties [52]. The comparison of crotoxin isoforms revealed that the stability of the complex plays a major role in its pharmacological action [53]. The origins of the isoforms were also identified. Multiple CA isoforms result from post-translational modifications occurring on a precursor pro-CA, while CB isoforms result from the expression of different messenger RNAs [54][55][56]. The presence of various PLA2 isoforms with diverse pharmacological activities could be explained by the accelerated evolution of exon regions after the duplication of a common ancestral gene, indicating the rapid adaptation of snakes for defense and predation.
During work in the Unité des Venins, G. Faure and Igor Krizaj demonstrated that crotoxin binds with a high affinity to a protein receptor from the presynaptic membrane of neuromuscular junctions in the electric organ of Torpedo marmorata [57][58]. They purified a 48 kDa crotoxin acceptor protein from Torpedo (CAPT) and characterized its binding to the receptor by surface plasmon resonance (SPR) characterization [58][59]. This study showed the formation of a ternary complex, CA CB Receptor, and the dissociation of CA at equilibrium.
Furthermore, G. Faure and Jonas Perales in the Unité des Venins identified, purified, and characterized a natural PLA2 inhibitor, the crotoxin inhibitor from Crotalus serum (CICS), from the blood of Crotalus durissus terrificus [60][61][62]. The CICS is an acidic 130 kDa oligomeric glycoprotein formed by the non-covalent association of 23–25 kDa subunits. This natural PLA2 inhibitor protects the rattlesnake from its own venom. It neutralizes the lethal and PLA activities of crotoxin, CB, and other PLA2s from the Viperidae family by binding to the CB subunit and preventing the association of CB with CA. Since the molecular mechanism underlying the interaction between crotoxin and CICS seems to be identical to that of crotoxin with its protein receptor CAPT, it was suggested that CICS acts physiologically as a false soluble crotoxin receptor, retaining the toxin in the vascular system of the snake and thereby preventing its toxic effects on the neuromuscular junction [60].
Valerie Choumet and colleagues from the C. Bon unit investigated the immunological aspects of crotoxin and other neurotoxic PLA2s [63][64][65][66][67][68][69]. Notably, they discovered that the acidic subunit CA of crotoxin interacts with the basic, single-chain ammodytoxin or agkistrodotoxin from Viperidae venom, in agreement with the amino acid sequence similarities between CB and these single-chain toxins with PLA2 activity. They also produced monoclonal antibodies (mAbs) against the crotoxin CA and CB subunits and determined their dissociation constants, cross-reactivity, and neutralization ability. This research led them to propose functional regions in the crotoxin components, the toxic and enzymatic sites on CB, and to suggest interacting regions on the two components. Further crystallographic studies by G. Faure’s group established the CA-CB binding interface of crotoxin [70]. The knowledge of this interface could be useful in mapping the epitope of the neutralizing monoclonal antibody A56.36.
Subsequently, C. Bon and his colleagues at the Institut Pasteur explored the envenomation process and its treatment by serotherapy. They determined the toxicokinetics and toxicodynamics of snake and scorpion venom in experimental animal models such as rats and rabbits. The distribution of venom in the organism was analyzed after an injection by the intravenous or intramuscular route and in the absence or presence of mAb fragments, either able (Fab) or unable (Fab’2) to cross the renal route. Toxin levels were assessed using an enzyme-linked immunosorbent assay (ELISA) and radiolabeled proteins. The kinetic parameters were analyzed after the administration of varying toxin doses. After an intravenous injection, the toxin exhibited a biexponential decline corresponding to the distribution and subsequent elimination. An intramuscular injection allowed the toxin to reach the vascular compartment, after which it exhibited a monoexponential decrease. The severity of symptoms was correlated with the toxin levels in the plasma. An intravenous injection of antivenom serum was the most efficient route. Antivenom antibodies were able to neutralize the totality of the toxin in the vascular compartment. Fab fragments were less efficient than Fab’2 fragments, possibly due to their differing pharmacokinetics. It is likely that complexes of antibody fragments/venom constituents were eliminated by phagocytosis rather than the renal route due to their high molecular weight. Interestingly, treatment with Fab was found to induce oliguria, which might induce adverse effects of serotherapy in humans. When examining experimental scorpion envenomation in rats, it was found that a Fab’2 intravenous injection neutralized venom more rapidly than Fab. However, Fab was more effective when administrated intramuscularly. Furthermore, Fab was more efficient at preventing the early symptoms of envenomation than Fab’2. Both antibody fragments were equally effective at preventing late symptoms, irrespective of the administration route. Based on these results, it was recommended to intravenously inject a combination of Fab and Fab’2 for the treatment of scorpion envenomation [66][71][72][73][74][75][76][77][78][79][80]. This research allowed for the optimization of the envenomation treatment by serotherapy. C. Bon was a renowned international expert in this field and was regularly invited by the WHO to share his expertise on this topic.
C. Bon and his collaborators began exploring the involvement of snake venoms in blood coagulation, aiming to develop novel therapeutic molecules for treating thrombotic events. Phospholipids (PLs) play an important role in the coagulation process. Anionic PLs are exposed on the membrane of activated platelets and facilitate the formation of complexes containing PL and protein enzymes involved in the coagulation cascade. Certain mammalian-secreted PLA2s and some snake PLA2s exhibit an anticoagulant effect through PL hydrolysis and/or by competing with coagulation factors, thus preventing their assembly into complexes involved in the coagulation cascade. Anticoagulant PLA2s have been reported in venoms from Viperidae, Crotalidae, Elapidae, and Hydrophidae. An analysis of snake PLA2 has helped to better define the mode of action of the anticoagulant PLA2s. Conversely, snake venoms also contain metalloproteases that activate coagulation factors, inducing procoagulant activity. C. Bon and collaborators purified and characterized some of these proteases, such as the prothrombin activator from Bothrops atrox, a serine protease from Bungarus fasciatus that activates blood coagulation factor X, and a thrombin-like serine protease from Bothrops lanceolatus that activates fibrinogen into fibrin [81][82][83][84][85][86][87][88][89][90][91][92].
C. Bon was actively involved in teaching about venomous animals at the MNHN and about human envenomation and its treatment at various universities. He was active in the international network of the Institut Pasteur, which produced antivenomous sera, notably in Algeria, Tunisia, Morocco, and Iran. He was a member of numerous scientific societies, including the French Society of Biochemistry and Molecular Biology, the French Association of Pharmacologists, the Society of Neurosciences, the International Society of Toxinology, and the International Society of Thrombosis and Hemostasis. He also served on the scientific council of the Institut Pasteur of Paris (1995–1999) and the Institut Pasteur of Iran (1998–2004). C. Bon was a co-founder of the French Society for the Study of Toxins (SFET, Société Française d’Étude des Toxines) in 1992 and served as president from 2000 to 2008. In 2004, the unit of venoms at the Institut Pasteur was closed, and C. Bon moved to the Laboratory of Chemistry of Natural Substances of MNHN. He was warmly welcomed by Max Goyffon, finding a familiar environment for teaching. During his last years, he was primarily focused on teaching, advising the WHO on envenomation, and organizing SFET meetings [43][93].
5. Period of 2004–2023: Novel Therapeutic Peptides Based on Snake Toxins and Structural Analysis of Their Binding Interface with Biological Targets
G. Faure, an expert research associate at the Institut Pasteur, continued the work on the structure–function relationships and the mechanisms of action of snake venom toxins with various protein targets, including the coagulation factor Xa, proton-gated ion channels, and the cystic fibrosis transmembrane regulator (CFTR) chloride channel. She led a group on PLA2 toxins in the Structural Immunology Unit (2004–2011) directed by Graham Bentley, and later in the channel-receptors unit (2011–2023) directed by Pierre-Jean Corringer.
Collaborating with Frederick Saul in the Structural Immunology Unit, Faure’s group determined the crystal structure of the heterodimeric crotoxin from Crotalus durissus terrificus, isoform CA2CBb of Class I [70], two isoforms of the basic CB subunit of crotoxin (CBb and CBd) [70][94], and two isoforms of ammodytoxin (AtxA and AtxC) from Vipera ammodytes ammodytes [95]. The three-dimensional structure of crotoxin revealed the nature of the binding interface between the CA and CB subunits and allowed for the identification of the key amino acid residues responsible for significant differences in the stability, toxicity, and enzymatic activity of the two classes of crotoxin complexes [70][96].
Snake venom PLA2s exhibit a wide range of toxic and pharmacological effects, including neurotoxic (pre-synaptic or post-synaptic), myotoxic, and cardiotoxic activities; anticoagulant effects; the inhibition of platelet aggregation; hemolytic activity; internal hemorrhage; anti-hemorrhage activity; convulsing and hypotensive activity; the induction of edema; organ necrosis or tissue damage; bactericidal, anti-humoral, anti-HIV (human immunodeficiency virus), anti-Leishmania, and anti-Plasmodium activity; and anti-viral activity against the dengue and yellow fever viruses [97]. Faure’s group identified a number of Viperidae venom PLA2s that inhibit blood coagulation factor Xa (FXa) via a non-catalytic PL-independent mechanism [98]. The interaction sites on PLA2 and FXa were mapped using SPR protein–protein interaction measurements, mutagenesis studies, and molecular docking simulations [98][99]. Comparative structural studies of natural PLA2 isoforms, which differ in their neurotoxicity and anticoagulant activity, contributed to a better understanding of their mode of binding to human FXa and calmodulin [95][96][100]. Faure’s group also discovered that PLA2 binding to FXa prevents the oligomerization of PLA2 [101]. The identification of the anticoagulant sites on Atx and CB and an analysis of the spatial arrangement of the PLA2-FXa interface led to a better understanding of the hemostatic process. These results are important for the elaboration of novel anticoagulant agents (non-competitive FXa inhibitors) [100].
Working in the channel-receptors unit at Pasteur, Faure’s group along with other collaborators discovered that the CB subunit of crotoxin binds with a high affinity to novel protein targets such as the pentameric proton-gated channel GLIC [102] as well as the cystic fibrosis transmembrane regulator (CFTR) and its mutant ΔF508-CFTR, which is implicated in cystic fibrosis [103]. They demonstrated that CB is a negative allosteric modulator of GLIC [102] and a positive allosteric modulator of CFTR [103]. By a direct interaction with the nucleotide-binding domain NBD1 of CFTR, CB potentiates the chloride channel current and corrects the trafficking defect of misfolded ΔF508CFTR inside the cell [103]. Faure’s group identified the CB-ΔF508CFTR interface by molecular docking and by HDX-MS (hydrogen–deuterium exchange–mass spectrometry) studies [103]. For the therapeutic development of new anti-cystic fibrosis agents, G. Faure and collaborators used a structure-based in silico approach and designed peptides mimicking this CBb-ΔF508NBD1 interface [104]. Using electrophysiological and biophysical methods, they identified several peptides that interact with the ΔF508NBD1 domain of CFTR and increase the chloride channel activity [104]. These significant results provide a new class of CFTR potentiators and describe a novel approach for developing therapeutic peptides for the treatment of cystic fibrosis. Thus, the biochemical and structural characterization of the functional and pharmacological sites of snake venom PLA2 (Figure 6) or fragments of PLA2 in complexes with their biological targets is essential for the structure-based design of novel therapeutic agents [94].
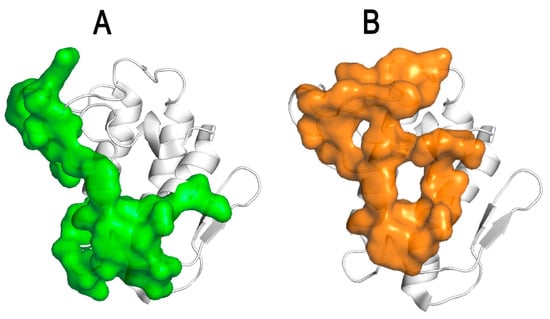
Figure 6. Identification of the pharmacological binding sites of CB, the PLA2 subunit of crotoxin, which is important for the structure-based design of new anticoagulant and anti-cystic fibrosis agents. (A) FXa-binding site of CB (in green) (adapted from Nemecz et al., 2020) [94]. (B) F508CFTR-binding site of CB (in orange) (adapted from Nemecz et al., 2020) [94].
References
- Charitos, I.A.; Gagliano-Candela, R.; Santacroce, L.; Bottalico, L. Venoms and Poisonings during the Centuries: A Narrative Review. Endocr. Metab. Immune Disord. Drug Targets 2022, 22, 558–570.
- Cilliers, L.; Retief, F.P. Poisons, poisoning and the drug trade in ancient Rome. Akroterion 2000, 45, 88–100.
- Cavaillon, J.M. From bacterial poisons to toxins: The early works of Pasteurians. Toxins 2022, 14, 759.
- Goyffon, M.; Chippaux, J.P. La découverte du sérum antivenimeux (10 février 1894). Biofutur 2008, 292, 32–35.
- Bochner, R. Paths to the discovery of antivenom serotherapy in France. J. Venom. Anim. Toxins Incl. Trop. Dis. 2016, 22, 20.
- Calmette, A. The Treatment of Animals Poisoned with Snake Venom by the Injection of Antivenomous Serum. Br. Med. J. 1896, 2, 399–400.
- Hawgood, B.J. Doctor Albert Calmette 1863–1933: Founder of antivenomous serotherapy and of antituberculous BCG vaccination. Toxicon 1999, 37, 1241–1258.
- Brygoo, E.R. La découverte de la sérothérapie antivenimeuse en 1894: Physalix et Bertrand ou Calmette? Bull. Ass. Anc. Elèves Inst. Pasteur 1985, 106, 10–22.
- Calmette, A. Contribution à l’étude du venin des serpents. Immunisation des animaux et traitement de l’envenimation. Ann. Inst. Pasteur 1894, 8, 275–291.
- Popoff, M.R.; Legout, S. Anaerobes and Toxins, a Tradition of the Institut Pasteur. Toxins 2023, 15, 43.
- Gilbrin, E. Gaston Ramon (1886–1963). Le soixantième anniversaire des anatoxines. Hist. Des Sci. Médicales 1984, 18, 53–60.
- Ramon, G.; Boquet, P.; Richou, R.; Delaunay-Ramon, R. Sur la production accélérée des sérums antivenimeux de différentes sortes au moyen des anavenins spécifiques et des substances stimulantes de l’immunité. Rev. Immunol. 1941, 6, 353–362.
- Ramon, G. Procédé pour accroïtre la production des antitoxines. Ann. Inst. Pasteur 1926, 40, 1–10.
- Boquet, P.; Détrait, J.; Farzanpay, R. Biochemical and immunological studies on snake venom. 3. Study of analogues of the Naja nigricollis venom alpha antigen. Ann. Inst. Pasteur 1969, 116, 522–542.
- Boquet, P.; Izard, Y.; Detrait, J. Studies on the spreading factor of snake venoms. C. R. Seances Soc. Biol. Fil. 1958, 152, 1363–1365.
- Cheymol, J.; Boquet, P.; Bourillet, F.; Detrait, J.; Roch-Arveiller, M. Comparison of the principal pharmacologic properties of different venoms of Echis carinatus (Viperides). Arch. Int. Pharmacodyn. Ther. 1973, 205, 293–304.
- Detrait, J.; Boquet, P. Separation of the constituents of Naja naja venin by electrophoresis. C. R. Hebd. Seances Acad. Sci. 1958, 246, 1107–1109.
- Detrait, J.; Izard, Y.; Boquet, P. Antigenic relations between a lethal factor in the venom of Echis carinata and neurotoxins in the venoms of Na ja na ja and Na ja nigricollis. C. R. Seances Soc. Biol. Fil. 1960, 154, 1163–1165.
- Tazieff-Depierre, F.; Pierre, J. Curarizing action of alpha toxin of Naja nigricollis. C. R. Acad. Hebd. Seances Acad. Sci. D 1966, 263, 1785–1788.
- Boquet, P.; Poilleux, G.; Dumarey, C.; Izard, Y.; Ronsseray, A.M. An attempt to classify the toxic proteins of Elapidae and Hydrophiidae venoms. Toxicon 1973, 11, 333–340.
- Boquet, P. Snake venoms. 2. The chemical constitution of snake venoms and immunity against venoms. Toxicon 1966, 3, 243–279.
- Boquet, P. Snake Venoms. I. Physiopathology of Envenomation and Biological Properties of Venoms. Toxicon 1964, 15, 5–41.
- Boquet, P.; Dumarey, C.; Joseph, D. The antigenicity of toxic proteins from the venoms of Elapidae and Hydrophiidae. C. R. Acad. Hebd. Seances Acad. Sci. D 1977, 284, 2439–2442.
- Boquet, P.; Izard, Y.J.M.; Meaume, J. Biochemical and immunological research on snale venom. I. Trial separations of the antigens of Naja nigricollis venom by Sephadex filtration. Ann. Inst. Pasteur 1966, 111, 719–732.
- Boquet, P.; Izard, Y.; Ronsseray, A.M. Attempt at classification of toxic proteins extracted from snake venoms. C. R. Acad. Hebd. Seances Acad. Sci. D 1970, 271, 1456–1459.
- Boquet, P.; Izard, Y.; Ronsseray, A.M. An attempt to classify by serological techniques the toxic proteins of low molecular weight extracted from Elapidae and Hydrophiidae venoms. Taiwan Yi Xue Hui Za Zhi 1972, 71, 307–310.
- Meunier, J.C.; Olsen, R.W.; Menez, A.; Fromageot, P.; Boquet, P.; Changeux, J.P. Some physical properties of the cholinergic receptor protein from Electrophorus electricus revealed by a tritiated alpha-toxin from Naja nigricollis venom. Biochemistry 1972, 11, 1200–1210.
- Tazieff-Depierre, F. Pharmacologic properties of toxins from scorpion venom (Androctonus australis). C. R. Acad. Hebd. Seances Acad. Sci. D 1968, 267, 240–243.
- Lièvremont, M.; Czajka, M.; Tazieff-Depierre, F. Calcium cycle at the neuromuscular junction. C. R. Acad. Hebd. Seances Acad. Sci. D 1969, 268, 379–382.
- Tazieff-Depierre, F.; Czajka, M.; Lowagie, C. Pharmacologic action of pure Naja nigricollis venom fractions and calcium liberation in striated muscles. C. R. Acad. Hebd. Seances Acad. Sci. D 1969, 268, 2511–2514.
- Tazieff-Depierre, F. Tetrodotoxin, calcium and scorpion venom. C. R. Acad. Hebd. Seances Acad. Sci. D 1970, 271, 1655–1657.
- Tazieff-Depierre, F. Scorpion venom, calcium and acetylcholine emission by nerve fibers in the ileum of guinea pigs. C. R. Acad. Hebd. Seances Acad. Sci. D 1972, 275, 3021–3024.
- Tazieff-Depierre, F.; Goudou, D.; Metezeau, P. Desensitization of the guinea pig ileum to scorpion venom by various concentrations of ions in the external medium. C. R. Acad. Hebd. Seances Acad. Sci. D 1974, 278, 2703–2706.
- Tazieff-Depierre, F.; Choucavy, M.; Métézeau, P. Pharmacologic properties of the toxins isolated from the sea anemone (Anemonia sulcata). C. R. Acad. Hebd. Seances Acad. Sci. D 1976, 283, 699–702.
- Tazieff-Depierre, F.; Métezeau, P. Effect of scorpion venom (Androctonus australis) on neuromuscular transmission inhibited by botulinum toxin in the frog. C. R. Acad. Hebd. Seances Acad. Sci. D 1977, 285, 1335–1338.
- Molgó, J.; Lemeignan, M.; Tazieff-Depierre, F. Enhancement by Anemonia sulcata toxin II of spontaneous quantal transmitter release from mammalian motor nerve terminals. Toxicon 1986, 24, 441–450.
- Métézeau, P.; Bourneau, R.; Mambrini, J.; Tazieff-Depierre, F. Effects of a neurotoxin isolated from the sea anemone, Anemonia sulcata, at frog neuromuscular junction (author’s transl). J. Physiol. 1979, 75, 873–879.
- Coraboeuf, E.; Deroubaix, E.; Tazieff-Depierre, T. Effect of toxin II isolated from scorpion venom on action potential and contraction of mammalian heart. J. Mol. Cell. Cardiol. 1975, 7, 643–653.
- Tazieff-Depierre, F. Cardiotoxicity in rats of purified gamma toxin isolated from venom of Naja nigricollis and of toxins extracted from scorpion venom. C. R. Acad. Hebd. Seances Acad. Sci. D 1975, 280, 1181–1184.
- Tazieff-Depierre, F.; Trethevie, E.R. Action of calcium chloride in the cardiotoxicity in the rat of purified gamma toxin isolated from Naja nigricollis venom. C. R. Acad. Hebd. Seances Acad. Sci. D 1975, 280, 137–140.
- Hallion, L.C. Delezenne. Annls Physiol. PhysicoChimie Biol. 1932, 8, 785–797.
- Lamy, R.; Rouyer, M. Relation entre les valeurs antigéniques des venims blancs et jaunes de Vipera aspis. Bull. Acad. Med. 1947, 131, 114–116.
- Goyffon, M.; Faure, G. Obituary: Cassian Bon—A scientist fascinated by toxins and their therapeutic potential. Toxicon 2008, 52, 400–403.
- Hendon, R.A.; Fraenkel-Conrat, H. Biological roles of the two components of crotoxin. Proc. Natl. Acad. Sci. USA 1971, 68, 1560–1563.
- Bon, C.; Changeux, J.P.; Jeng, T.W.; Fraenkel-Conrat, H. Postsynaptic effects of crotoxin and of its isolated subunits. Eur. J. Biochem. 1979, 99, 471–481.
- Bon, C.; Bouchier, C.; Choumet, V.; Faure, G.; Jiang, M.S.; Lambezat, M.P.; Radvanyi, F.; Saliou, B. Crotoxin, half-century of investigations on a phospholipase A2 neurotoxin. Acta Physiol. Pharmacol. Latinoam. 1989, 39, 439–448.
- Délot, E.; Bon, C. Differential effects of presynaptic phospholipase A2 neurotoxins on Torpedo synaptosomes. J. Neurochem. 1992, 58, 311–319.
- Délot, E.; Bon, C. Model for the interaction of crotoxin, a phospholipase A2 neurotoxin, with presynaptic membranes. Biochemistry 1993, 32, 10708–10713.
- Ollivier-Bousquet, M.; Radvanyi, F.; Bon, C. Crotoxin, a phospholipase A2 neurotoxin from snake venom, interacts with epithelial mammary cells, is internalized and induces secretion. Mol. Cell. Endocrinol. 1991, 82, 41–50.
- Radvanyi, F.R.; Bon, C. Catalytic activity and reactivity with p-bromophenacyl bromide of the phospholipase subunit of crotoxin. Influence of dimerization and association with the noncatalytic subunit. J. Biol. Chem. 1982, 257, 12616–12623.
- Faure, G.; Bon, C. Several isoforms of crotoxin are present in individual venoms from the South American rattlesnake Crotalus durissus terrificus. Toxicon 1987, 25, 229–234.
- Faure, G.; Bon, C. Crotoxin, a phospholipase A2 neurotoxin from the South American rattlesnake Crotalus durissus terrificus: Purification of several isoforms and comparison of their molecular structure and of their biological activities. Biochemistry 1988, 27, 730–738.
- Faure, G.; Harvey, A.L.; Thomson, E.; Saliou, B.; Radvanyi, F.; Bon, C. Comparison of crotoxin isoforms reveals that stability of the complex plays a major role in its pharmacological action. Eur. J. Biochem. 1993, 214, 491–496.
- Bouchier, C.; Boulain, J.C.; Bon, C.; Ménez, A. Analysis of cDNAs encoding the two subunits of crotoxin, a phospholipase A2 neurotoxin from rattlesnake venom: The acidic non enzymatic subunit derives from a phospholipase A2-like precursor. Biochim. Biophys. Acta 1991, 1088, 401–408.
- Faure, G.; Choumet, V.; Bouchier, C.; Camoin, L.; Guillaume, J.L.; Monegier, B.; Vuilhorgne, M.; Bon, C. The origin of the diversity of crotoxin isoforms in the venom of Crotalus durissus terrificus. Eur. J. Biochem. 1994, 223, 161–164.
- Faure, G.; Guillaume, J.L.; Camoin, L.; Saliou, B.; Bon, C. Multiplicity of acidic subunit isoforms of crotoxin, the phospholipase A2 neurotoxin from Crotalus durissus terrificus venom, results from posttranslational modifications. Biochemistry 1991, 30, 8074–8083.
- Krizaj, I.; Faure, G.; Gubensek, F.; Bon, C. Re-examination of crotoxin-membrane interactions. Toxicon 1996, 34, 1003–1009.
- Krizaj, I.; Faure, G.; Gubensek, F.; Bon, C. Neurotoxic phospholipases A2 ammodytoxin and crotoxin bind to distinct high-affinity protein acceptors in Torpedo marmorata electric organ. Biochemistry 1997, 36, 2779–2787.
- Faure, G.; Copic, A.; Le Porrier, S.; Gubensek, F.; Bon, C.; Krizaj, I. Crotoxin acceptor protein isolated from Torpedo electric organ: Binding properties to crotoxin by surface plasmon resonance. Toxicon 2003, 41, 509–517.
- Faure, G. Natural inhibitors of toxic phospholipases A(2). Biochimie 2000, 82, 833–840.
- Faure, G.; Villela, C.; Perales, J.; Bon, C. Interaction of the neurotoxic and nontoxic secretory phospholipases A2 with the crotoxin inhibitor from Crotalus serum. Eur. J. Biochem. 2000, 267, 4799–4808.
- Perales, J.; Villela, C.; Domont, G.B.; Choumet, V.; Saliou, B.; Moussatché, H.; Bon, C.; Faure, G. Molecular structure and mechanism of action of the crotoxin inhibitor from Crotalus durissus terrificus serum. Eur. J. Biochem. 1995, 227, 19–26.
- Bon, C.; Choumet, V.; Delot, E.; Faure, G.; Robbe-Vincent, A.; Saliou, B. Different evolution of phospholipase A2 neurotoxins (beta-neurotoxins) from Elapidae and Viperidae snakes. Ann. N. Y. Acad. Sci. 1994, 710, 142–148.
- Choumet, V.; Bouchier, C.; Délot, E.; Faure, G.; Saliou, B.; Bon, C. Structure and function relationship of crotoxin, a heterodimeric neurotoxic phospholipase A2 from the venom of a South-American rattlesnake. Adv. Exp. Med. Biol. 1996, 391, 197–202.
- Choumet, V.; Faure, G.; Robbe-Vincent, A.; Saliou, B.; Mazié, J.C.; Bon, C. Immunochemical analysis of a snake venom phospholipase A2 neurotoxin, crotoxin, with monoclonal antibodies. Mol. Immunol. 1992, 29, 871–882.
- Choumet, V.; Jiang, M.S.; Radvanyi, F.; Ownby, C.; Bon, C. Neutralization of lethal potency and inhibition of enzymatic activity of a phospholipase A2 neurotoxin, crotoxin, by non-precipitating antibodies (Fab). FEBS Lett. 1989, 244, 167–173.
- Choumet, V.; Lafaye, P.; Demangel, C.; Bon, C.; Mazié, J.C. Molecular mimicry between a monoclonal antibody and one subunit of crotoxin, a heterodimeric phospholipase A2 neurotoxin. Biol. Chem. 1999, 380, 561–568.
- Choumet, V.; Lafaye, P.; Mazié, J.C.; Bon, C. A monoclonal antibody directed against the non-toxic subunit of a dimeric phospholipase A2 neurotoxin, crotoxin, neutralizes its toxicity. Biol. Chem. 1998, 379, 899–906.
- Choumet, V.; Saliou, B.; Fideler, L.; Chen, Y.C.; Gubensek, F.; Bon, C.; Delot, E. Snake-venom phospholipase A2 neurotoxins. Potentiation of a single-chain neurotoxin by the chaperon subunit of a two-component neurotoxin. Eur. J. Biochem. 1993, 211, 57–62.
- Faure, G.; Xu, H.; Saul, F.A. Crystal structure of crotoxin reveals key residues involved in the stability and toxicity of this potent heterodimeric β-neurotoxin. J. Mol. Biol. 2011, 412, 176–191.
- Dekhil, H.; Wisner, A.; Marrakchi, N.; El Ayeb, M.; Bon, C.; Karoui, H. Molecular cloning and expression of a functional snake venom serine proteinase, with platelet aggregating activity, from the Cerastes cerastes viper. Biochemistry 2003, 42, 10609–10618.
- Jandrot-Perrus, M.; Lagrue, A.H.; Leduc, M.; Okuma, M.; Bon, C. Convulxin-induced platelet adhesion and aggregation: Involvement of glycoproteins VI and IaIIa. Platelets 1998, 9, 207–211.
- Krifi, M.N.; Savin, S.; Debray, M.; Bon, C.; El Ayeb, M.; Choumet, V. Pharmacokinetic studies of scorpion venom before and after antivenom immunotherapy. Toxicon 2005, 45, 187–198.
- Parry, M.A.; Jacob, U.; Huber, R.; Wisner, A.; Bon, C.; Bode, W. The crystal structure of the novel snake venom plasminogen activator TSV-PA: A prototype structure for snake venom serine proteinases. Structure 1998, 6, 1195–1206.
- Hammoudi-Triki, D.; Lefort, J.; Rougeot, C.; Robbe-Vincent, A.; Bon, C.; Laraba-Djebari, F.; Choumet, V. Toxicokinetic and toxicodynamic analyses of Androctonus australis hector venom in rats: Optimization of antivenom therapy. Toxicol. Appl. Pharmacol. 2007, 218, 205–214.
- Audebert, F.; Urtizberea, M.; Sabouraud, A.; Scherrmann, J.M.; Bon, C. Pharmacokinetics of Vipera aspis venom after experimental envenomation in rabbits. J. Pharmacol. Exp. Ther. 1994, 268, 1512–1517.
- Choumet, V.; Audebert, F.; Rivière, G.; Sorkine, M.; Urtizberea, M.; Sabouraud, A.; Scherrmann, J.M.; Bon, C. New approaches in antivenom therapy. Adv. Exp. Med. Biol. 1996, 391, 515–520.
- Rivière, G.; Choumet, V.; Audebert, F.; Sabouraud, A.; Debray, M.; Scherrmann, J.M.; Bon, C. Effect of antivenom on venom pharmacokinetics in experimentally envenomed rabbits: Toward an optimization of antivenom therapy. J. Pharmacol. Exp. Ther. 1997, 281, 1–8.
- Rivière, G.; Choumet, V.; Saliou, B.; Debray, M.; Bon, C. Absorption and elimination of viper venom after antivenom administration. J. Pharmacol. Exp. Ther. 1998, 285, 490–495.
- Calderón-Aranda, E.S.; Rivière, G.; Choumet, V.; Possani, L.D.; Bon, C. Pharmacokinetics of the toxic fraction of Centruroides limpidus limpidus venom in experimentally envenomed rabbits and effects of immunotherapy with specific F(ab’)2. Toxicon 1999, 37, 771–782.
- Stroka, A.; Donato, J.L.; Bon, C.; Hyslop, S.; de Araújo, A.L. Purification and characterization of a hemorrhagic metalloproteinase from Bothrops lanceolatus (Fer-de-lance) snake venom. Toxicon 2005, 45, 411–420.
- Lôbo de Araújo, A.; Kamiguti, A.; Bon, C. Coagulant and anticoagulant activities of Bothrops lanceolatus (Fer de lance) venom. Toxicon 2001, 39, 371–375.
- Wisner, A.; Braud, S.; Bon, C. Snake venom proteinases as tools in hemostasis studies: Structure-function relationship of a plasminogen activator purified from Trimeresurus stejnegeri venom. Haemostasis 2001, 31, 133–140.
- Braud, S.; Bon, C.; Wisner, A. Snake venom proteins acting on hemostasis. Biochimie 2000, 82, 851–859.
- Mounier, C.M.; Bon, C.; Kini, R.M. Anticoagulant venom and mammalian secreted phospholipases A(2): Protein-versus phospholipid-dependent mechanism of action. Haemostasis 2001, 31, 279–287.
- Mounier, C.; Franken, P.A.; Verheij, H.M.; Bon, C. The anticoagulant effect of the human secretory phospholipase A2 on blood plasma and on a cell-free system is due to a phospholipid-independent mechanism of action involving the inhibition of factor Va. Eur. J. Biochem. 1996, 237, 778–785.
- Hofmann, H.; Bon, C. Blood coagulation induced by the venom of Bothrops atrox. 2. Identification, purification, and properties of two factor X activators. Biochemistry 1987, 26, 780–787.
- Zhang, Y.; Wisner, A.; Xiong, Y.; Bon, C. A novel plasminogen activator from snake venom. Purification, characterization, and molecular cloning. J. Biol. Chem. 1995, 270, 10246–10255.
- Lôbo de Araújo, A.; Donato, J.L.; Bon, C. Purification from Bothrops lanceolatus (fer de lance) venom of a fibrino(geno)lytic enzyme with esterolytic activity. Toxicon 1998, 36, 745–758.
- Braud, S.; Parry, M.A.; Maroun, R.; Bon, C.; Wisner, A. The contribution of residues 192 and 193 to the specificity of snake venom serine proteinases. J. Biol. Chem. 2000, 275, 1823–1828.
- Zhang, Y.; Xiong, Y.L.; Bon, C. An activator of blood coagulation factor X from the venom of Bungarus fasciatus. Toxicon 1995, 33, 1277–1288.
- Hofmann, H.; Bon, C. Blood coagulation induced by the venom of Bothrops atrox. 1. Identification, purification, and properties of a prothrombin activator. Biochemistry 1987, 26, 772–780.
- Goyffon, M. Cassian Bon, 1944–2008. Rencontres Toxinologie 2008, RT16, 9–10.
- Nemecz, D.; Ostrowski, M.; Ravatin, M.; Saul, F.; Faure, G. Crystal Structure of Isoform CBd of the Basic Phospholipase A(2) Subunit of Crotoxin: Description of the Structural Framework of CB for Interaction with Protein Targets. Molecules 2020, 25, 5290.
- Saul, F.A.; Prijatelj-Znidarsic, P.; Vulliez-le Normand, B.; Villette, B.; Raynal, B.; Pungercar, J.; Krizaj, I.; Faure, G. Comparative structural studies of two natural isoforms of ammodytoxin, phospholipases A2 from Vipera ammodytes ammodytes which differ in neurotoxicity and anticoagulant activity. J. Struct. Biol. 2010, 169, 360–369.
- Faure, G.; Saul, F. Crystallographic characterization of functional sites of crotoxin and ammodytoxin, potent β-neurotoxins from Viperidae venom. Toxicon. 2012, 60, 531–538.
- Faure, G.; Porowinska, D.; Saul, F. Crototoxin from Crotalus durissus terrificus and Crotoxin-Related Proteins: Structure and Function Relationship. In Toxins and Drug Discovery; Gopalakrishnakone, P., Cruz, L.J., Luo, S., Eds.; Springer: Dordrecht, The Netherlands, 2017; pp. 3–20.
- Faure, G.; Gowda, V.T.; Maroun, R.C. Characterization of a human coagulation factor Xa-binding site on Viperidae snake venom phospholipases A2 by affinity binding studies and molecular bioinformatics. BMC Struct. Biol. 2007, 7, 82.
- Faure, G.; Xu, H.; Saul, F. Anticoagulant Phospholipase A2 Which Bind to the Specific Soluble Receptor Coagulation Factor Xa. In Toxins and Hemostasis: From Bench to Bedside; Kini, R.M., Clemetson, K.J., Merkland, F.S., McClane, M.A., Moritta, T., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 201–217.
- Faure, G.; Saul, F. Structural and Functional Characterization of Anticoagulant, FXa-binding Viperidae Snake Venom Phospholipases A2. Acta Chim. Slov. 2011, 58, 671–677.
- Ostrovski, M.; Žnidaršič, P.P.; Raynal, B.; Saul, F.; Faure, G. Human coagulation factor Xa prevents oligomerization of anti-coagulant phospholipases A2. Toxin Rev. 2014, 33, 42–47.
- Ostrowski, M.; Porowinska, D.; Prochnicki, T.; Prevost, M.; Raynal, B.; Baron, B.; Sauguet, L.; Corringer, P.J.; Faure, G. Neurotoxic phospholipase A2 from rattlesnake as a new ligand and new regulator of prokaryotic receptor GLIC (proton-gated ion channel from G. violaceus). Toxicon 2016, 116, 63–71.
- Faure, G.; Bakouh, N.; Lourdel, S.; Odolczyk, N.; Premchandar, A.; Servel, N.; Hatton, A.; Ostrowski, M.K.; Xu, H.; Saul, F.A.; et al. Rattlesnake Phospholipase A2 Increases CFTR-Chloride Channel Current and Corrects ∆F508CFTR Dysfunction: Impact in Cystic Fibrosis. J. Mol. Biol. 2016, 428, 2898–2915.
- Ravatin, M.; Odolczyk, N.; Servel, N.; Guijarro, J.I.; Tagat, E.; Chevalier, B.; Baatallah, N.; Corringer, P.J.; Lukács, G.L.; Edelman, A.; et al. Design of Crotoxin-Based Peptides with Potentiator Activity Targeting the ΔF508NBD1 Cystic Fibrosis Transmembrane Conductance Regulator. J. Mol. Biol. 2023, 435, 167929.
More
Information
Subjects:
Toxicology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
846
Revisions:
2 times
(View History)
Update Date:
07 Aug 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No