Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Andrés Arias | -- | 3077 | 2023-07-27 13:10:58 | | | |
| 2 | Camila Xu | Meta information modification | 3077 | 2023-07-28 02:22:00 | | | | |
| 3 | Camila Xu | Meta information modification | 3077 | 2023-09-05 10:31:25 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Arias, A.; Woodin, S.A.; Paxton, H. Diopatra. Encyclopedia. Available online: https://encyclopedia.pub/entry/47356 (accessed on 15 June 2026).
Arias A, Woodin SA, Paxton H. Diopatra. Encyclopedia. Available at: https://encyclopedia.pub/entry/47356. Accessed June 15, 2026.
Arias, Andrés, Sarah A. Woodin, Hannelore Paxton. "Diopatra" Encyclopedia, https://encyclopedia.pub/entry/47356 (accessed June 15, 2026).
Arias, A., Woodin, S.A., & Paxton, H. (2023, July 27). Diopatra. In Encyclopedia. https://encyclopedia.pub/entry/47356
Arias, Andrés, et al. "Diopatra." Encyclopedia. Web. 27 July, 2023.
Copy Citation
The genus Diopatra is a major driver in sedimentary systems, altering the structure of habitats and changing the frequency of refugia and predator access to prey. It is taken as prey by a variety of shorebirds, crustaceans, and fish and used worldwide as bait.
Annelida
Onuphidae
morphology
reproduction
ecology
1. Introduction
Bristle-worms or polychaetes are marine annelids that occur from the littoral zone to the deepest trenches, inhabiting soft and rocky bottoms or leading a pelagic life. Among them, members of the family Onuphidae Kinberg, 1865 [1] are among the most important polychaetous annelids in soft sediment communities worldwide. Members of the genus Diopatra Audouin and Milne Edwards, 1833 [2] have been coveted as fishing bait for almost two centuries and are known as ecosystem engineers, stabilizing sediments with their tubes and therefore increasing the structural complexity and biodiversity of their infaunal habitat [3]. Diopatra has attracted attention in the past decade not only for its surprising and unknown diversity [4][5][6], but also for its role as an indicator of climate change [7][8], a sentinel group for drugs in marine environments [9], and an example of protandric hermaphroditism [10][11][12].
2. History and Diversity
The first named species was Nereis cuprea Bosc, 1802 [13] from South Carolina, USA, stated to be very common in the area of Charleston. It was grouped with the ‘‘Néréides à bouches armée de mâchoires’‘ (nereids armed with a toothed mouth), and in 1833, it was transferred to the newly erected genus Diopatra. The presence of branchial filaments, arranged in a spiral around the central branchial trunk, is the main defining character of Diopatra, one of the oldest and most beautiful genera of polychaetes of the family Onuphidae. It is one of the few polychaete genera that is defined by a single characteristic and the status of which has never been amended or changed.
It is the largest known genus of the family, represented by 67 recognized species worldwide, of which 21 were described before 1900, 43 between 1900 and 2000, and 13 in the new millennium [14]. Although the genus is uniquely defined by its autapomorphy of possessing spiralled branchiae, specific identification is notoriously difficult as they are superficially very similar and lack clear diagnostic features. They can be distinguished only by combinations of characteristics that show various degrees of overlap and variability. This problem of species delineation has been recognized for a long time [15][16][17][18] and even led the famous French polychaetologist Pierre Fauvel [19] to make the statement that most described species of Diopatra constitute a single variable worldwide species, namely D. neapolitana (Delle Chiaje, 1841) [20].
Notable taxonomic revisions [16][17][21][22][23] have led to the recognition of new diagnostic characteristics, to which the advent of scanning electron microscopy (SEM) has contributed greatly. Most recently, new morphological character sets have been explored and evaluated including parapodial lobes and folds and maxillary characteristics [6]. However, the most significant aid to phylogenetic analyses came with integrative studies and the application of sequence-based genetic methods. The earliest genetic analyses investigated the identity of Western European species [4][7][12][18] and South American faunas [6]. The present SI will expand it to West African and North and South American Diopatra diversity (see papers by Hektoen et al., 2021; Sotka et al., 2023) [24][25].
3. Morphology
Diopatra, with its often bright colouration and spiralled branchiae, is one of the most beautiful onuphid genera (Figure 1). The smallest species measure only a few centimetres in length while a large live D. neapolitana can be 80 cm long with almost 400 chaetigers and over 1 cm in width [12]. The anterior part of the body bears a small head or prostomium with highly developed sensory structures (Figure 1D), followed by a strongly muscularized anterior region, grading into a softer median and posterior end. The animals are adapted to a tubicolous existence in which the anterior body can be partly everted from the tube for feeding and tube construction but can be rapidly withdrawn into the protective tube at the slightest signs of danger.

Figure 1. Photographs of Diopatra spp. living specimens (anterior ends): (A) D. aciculata, lateral view; (B) D. gallardoi, dorsal view; (C) D. aciculata, dorsal view; (D) detailed view of the prostomium of the same; (E) D. neapolitana, dorsal view.
The prostomium (Figure 2A) bears a pair of anterior sensory lips and five appendages with basal ringed ceratophores and distal ceratostyles. The two anterolateral appendages are palps, and the inner three are antennae. The ceratostyles are covered in combined sensory/secretory structures termed ‘Sinnesknospen’ or sensory buds [26]. Pflugfelder described their histology, showing that cilia project through the cuticle from a central sensory cell and that the ciliated cell is accompanied by one or two serous glands opening to the surface [26]. Although present in all onuphids, these are arranged in conspicuous rows in Diopatra (Figure 2C). Small eyespots are only present in juveniles; oval to almost circular structures that had previously been interpreted as eyes, located at the posterior part of the prostomium, are nuchal organs forming ciliated grooves (Figure 2B).
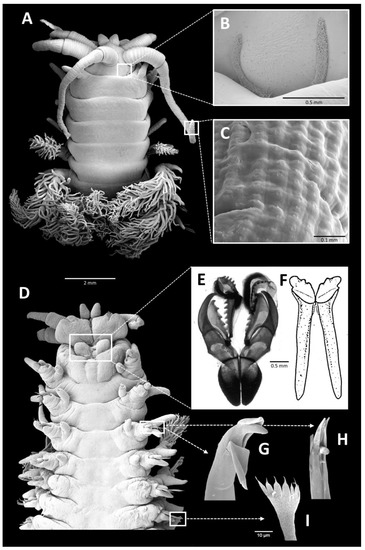
Figure 2. Diopatra neapolitana, scanning electron micrographs/drawings: (A) anterior end, dorsal view; (B) detailed view of nuchal groove; (C) detailed view of the sensory buds of antennae; (D) anterior end, ventral view; (E) maxillae; (F) mandibles; (G) falcate or unidentate moderately robust simple hook from modified parapodium; (H) bidentate lower slender pseudocompound hook from same; (I) pectinate chaeta.
The anterior chaetigers (Figure 2) are modified for quick propulsion from and retraction back into the tube. The well-developed longitudinal muscles provide the power, aided by the modified enlarged parapodia, which are directed anterolaterally and bear specialized hooks. The branchiae commence from parapodia 4 to 5 (Figure 2A,D) and reach their greatest development by about chaetiger 20. Thereafter, the filaments become reduced and are absent at about chaetiger 50–70. The median and posterior part of the body is basically a container for gut and reproductive products.
As an onuphid, Diopatra belongs to the order Eunicida, possessing a complex jaw apparatus consisting of a pair of ventral mandibles and dorsal maxillae (Figure 2E,F). The jaws are hardened cuticular structures, composed of calcium carbonate and/or scleroproteins. As a result of their durability, they have a good fossil record. While the earliest known eunicidan jaw elements of extinct families date from the latest Cambrian, fossils of the extant Onuphidae have not been identified but would be expected to be much younger [27][28]. Onuphid jaws become visible in three-day-old larvae. While the larval mandibles are retained and added on throughout the lifetime, the maxillae change to a brief juvenile version before the adult stage that will thereafter moult periodically to achieve growth [29]. The jaw kinematics of Diopatra spp. have been observed by filming individuals biting, and a comparison with Lumbrineris spp. was interpreted to be consistent with their differences in diet [30].
4. Geographical Distribution
The genus occurs in all major oceans but is best represented in warmer waters, where it is found in shallow depths. However, two specimens of Diopatra sp. from South Georgia Island, Antarctica have been reported from a depth of 217 m [31]. Although more than 60 species are recognized, many are only known from their original descriptions. Some of the best-known names, such as D. neapolitana, D. amboinensis Audouin and Milne Edwards, 1833 [2] and D. cuprea, are credited with worldwide distributions that are not trustworthy [25].
The most remarkable knowledge increase in the diversity and reproductive biology of Diopatra is from European waters. Whilst D. neapolitana was thought to be the only accepted European representative of the genus until quite recently [8][18], they presently number ten, with seven species from France, Portugal, and Spain described since 2010. The most diverse region is Macaronesia, a region comprising five eastern North Atlantic archipelagos: the Azores, Madeira, the Salvage Islands, the Canary Islands, and the Cape Verde Islands from which nine of the ten species have been reported [5].
West Africa is a very rich area that has been previously studied [15][32][33] and where new active studies are in progress. An integrated unpublished master’s study discovered 14 species of Diopatra, of which five were previously known and nine were new to science. Some of this work has been presented as a paper in this SI (see Hektoen et al., 2021) [24].
North America has a surprisingly small representation of Diopatra species, with only D. cuprea reported from its eastern coast [34]. However, this is an underrepresentation, since the famous D. cuprea represents a species complex consisting at least of five lineages (see paper by Sotka et al., 2023 in this SI) [25]. Nine species have been reported from the western USA and Mexico, largely as a result of the studies by Olga Hartman and Kristian Fauchald [16][21].
In the southern hemisphere, Diopatra fauna are well represented in Australia, where nine species have been reported [17]. Another centre of a rich Diopatra history is Brazil, where the reported 14 species from the first half of the 20th century were established by European workers. However, only half of these are presently accepted, but a keen group of polychaetologists is studying these fauna with integrated methods, which has recently resulted in the description of four new species and the strong suggestion that the range of D. cuprea may extend southward only into the Caribbean Sea [6].
5. Reproduction and Development
Diopatra species are annual breeders with discrete or interrupted breeding seasons. They were thought to be predominantly gonochoristic, as ripe specimens could be distinguished by the colour of their sexual products through their body wall; eggs were yellowish to greenish and sperm whitish to cream-coloured. However, an intensive study of D. neapolitana, combining field observations with a histological study of monthly collected individuals, revealed that the species is a sequential hermaphrodite of the protandry type. The studied Spanish population consisted of pure males in the smallest-sized class, hermaphrodites in the medium-sized classes, and pure females in the largest ones [12]. External detection of sperm or eggs only confirms the dominant sexuality of individuals at one time point. The reproductive cycle of D. neapolitana in northern Spain and its timing of development is fully described in Arias et al., 2016 [12]. See also the study by Escobar-Ortega et al. [35] in this SI, suggesting that water temperature is one of the most important drivers of the reproductive cycle for this species.
Further studies on other species of Diopatra suggest that the hermaphroditic condition may be widespread in this genus [10][11]. Another European Diopatra species, D. biscayensis (Fauchald et al. (2012)) [36] from northern Spain and France is a protandric simultaneous hermaphrodite [11]. This has also been suggested by Arias et al. (2013) [10] for the presumed gonochoristic population of D. marocensis (Paxton et al., 1995 [37]) from the Portuguese coasts studied by Pires et al. (2012) [38]. The sequential hermaphroditism can be explained by the ‘size advantage’ hypothesis, originally developed by Charnov (1982) [39] and later expanded by Ghiselin (1987) [40]. This hypothesis postulates that the reproductive success of an individual as a male or as a female is closely linked to its body size or age and that the relationship between reproductive success and size/age differs for each sex. Thus, protandry is expected when a large body size increases female fecundity more than male fertility. Protandry is the most common type of sequential hermaphroditism among polychaetes and marine invertebrates [41]. In Diopatra, a larger female can produce more eggs than a small female, and since they are broadcast spawners with random fertilization, the reproductive success of small and large males is likely to be almost the same, and this suppresses the advantages of protogyny.
Other studies of the reproductive biology of Diopatra spp. (mainly of D. neapolitana from different locations, e.g., [42][43]), despite assuming the gonochoristic condition of the species, have concluded unambiguously that within the studied populations, the smallest female found was larger than the smallest male and the male:female ratio deviated from the expected Fisher 1:1 sex ratio for gonochoristic or dioecious species. These observations strongly suggest protandry, leading to consider the possibility that sequential hermaphroditism is largely underreported for Diopatra species. Other studies in a smaller species of Diopatra, D. marocensis, revealed that this species was hermaphroditic. However, within the studied northern Iberian populations, no pure males or females were found, and the species was considered a simultaneous hermaphrodite [10]. This type of hermaphroditism was previously reported in Diopatra sp. from Sumatra [44]. Simultaneous hermaphroditism is typically correlated with brooding behaviour, direct or lecithotrophic development, and a sedentary habit [10][40]. Although these relationships are not very well defined, many members of the genus Diopatra satisfy these requirements [17]. Consequently, histological reproductive studies should be extended to more species within the genus which remain poorly known biologically.
Paxton (2016) [17] summarized the known patterns of larval release or retention across the genus. She described four known patterns, including (1) brooding within the tube, typical of small species presumably for protection of gametes and juveniles; (2) deposition of eggs not inside the parental tube but in a gelatinous matrix or sac attached to the distal end of the tube where initial development occurs; and (3) direct release without a retention stage. The latter two are typical of larger species such as D. neapolitana and D. biscayensis [11][12]. Table 1 is an updated version of Paxton’s original table. Two observations are immediately obvious. First, brooding may be to a very late stage juvenile such as those of D. marocensis Paxton et al., 1995 [37] (Figure 4) and D. tuberculantennata Budaeva and Fauchald, 2008 [45] which were collected with 34- and 28-chaetiger juveniles in adult tubes respectively [45][46]. Juvenile D. marocensis with 32–40 chaetigers were collected with adults in their own tubes in the sediment, indicating that once they leave the parental tube, they do not disperse any further but settle and build their own tubes among the adults [37].

Figure 4. Early development of the brooder D. marocensis from Villaviciosa estuary, Bay of Biscay: (A) protruding portion of tube in nature; (B) anterior end of mature worm; (C) tube dissection showing the brood; (D) detailed view of brooded fertilised eggs; (E) brooded juveniles of 8–10 chaetigers; (F) brooded juveniles of 12 chaetigers.
Table 1. Presence of a planktonic period in species of Diopatra. ~67 known species of Diopatra; only 14 with known larval type, i.e., brooded or released. Species marked with an asterisk are known to be hermaphroditic. Egg size in µm.
| Species | Locale | Egg Size | Planktonic Period | Reference |
|---|---|---|---|---|
| D. aciculata | S Australia | 230 | 4–5 days (settle at 4–6 chaetigers) |
Paxton and Safarik [29] |
| D. albimandibulata | Queensland, AU | 300 | brooding 3-chaetiger larvae |
Paxton [17] |
| D. biscayensis * | Bay of Biscay, SP | 260 | none to short, 1–2 days (released at 4–5 chaetigers, some +phototactic) |
Arias and Paxton [11] |
| D. cuprea | NE USA | 240 | 3–4 days (settle at 4 chaetigers) |
Allen [47] |
| D. gigova | W Australia | 1400 | none, brooding | Paxton [17] |
| D. lilliputiana | W Australia | 400–700 | none, brooded to ≥15 chaetigers | Paxton [17] |
| D. maculata | W Australia | 350 | gelatinous egg mass on outside of tube with 3–4-chaetiger larvae | Paxton [17] |
| D. marocensis * | Morocco, Aveiro PT | 600-620 | none, brooded to ≥23–34 chaetigers | Fadlaoui et al. [46], Pires et al. [38]; Arias et al. [10] |
| D. neapolitana * | Arcachon FR, Sardinia IT, Aveiro PT, N Spain, NW Spain | 240 | 3–4 days (settle at 3–5 chaetigers) |
Cazaux [48], Conti and Massa [49], Pires et al. [38], Arias et al. [12], Escobar-Ortega et al. [35] |
| D. nishii | Japan | 600–700 | none, brooded to ≥21 chaetigers | Paxton [17] |
| D. ornata | Catalina Island CA, USA | 235 | 4 days | Emerson [50], Fauchald [51] |
| D. sugokai | Maeshiba, Japan | 200 | 3 days (settle at 5 chaetigers) |
Choe [52]; Paxton [17] |
| D. tuberculantennata | Belize | ? | none, brooded to ≥25 chaetigers | Budaeva and Fauchald [45] |
| D. variabilis | Madras, India | 600 | none, brooded to ≥15 chaetigers | Krishnan [53] |
Second, it would appear that the planktonic period is quite short, two to four days, or non-existent for those with either a short duration egg mass external to the tube lumen or perhaps direct broadcast spawning (Table 1). Laboratory studies of D. cuprea by Allen (1959) [47] and of D. neapolitana by Cazaux (1970) [48] are still the classical references, with their detailed descriptions and illustrations of short-lived, free-swimming lecithotrophic larvae. In each case, the larvae have small eyespots and consist of 3–6 chaetigers by day three to five, when they start to settle and build their own tubes (Table 1).
6. Ecological Roles
The worms are tubicolous, building vertical tubes in the sediment that may extend their upper portion (known as the tube-cap) a few centimetres above the sediment (Figure 6). Their tubes vary from having scarcely any ornamentation, being mostly composed of silt and fine sand (Figure 6C), to being exquisitely ornamented with shells, seaweed, or any foreign material (Figure 6A,B), and often serve as good field characters as to species [11][49][50][51][52][53][54]. Almost unique to Diopatra is that materials such as shells are attached edge-on, imbricately, rather than flat, as in most tube-builder constructions.

Figure 6. Diopatra spp. habitats and tube types: (A) D. dentata from the rocky shore near Bondi (Sydney, SE Australia); (B) D. biscayensis from the sandy coves of Nouvelle-Aquitaine (Atlantic France, Bay of Biscay); (C) D. neapolitana from the sandy flats of Villaviciosa estuary (northern Spain, Bay of Biscay).
Mangum and Cox (1971) [55] investigated the feeding modalities and responses to chemical stimuli of D. cuprea, which has an extensively decorated and emergent tube-cap; its diet had been investigated previously by Mangum et al. (1968) [56], who reported seeing individuals browsing on material on the tube-cap and thought the tube-cap served in part as a food-catching device. Similarly, Brenchley and Tidball (1980) [57] showed that individuals of D. cuprea orient their tube-caps perpendicular to unidirectional flows, but under conditions with high densities, tube-caps are oriented parallel or perpendicular to one another and D. cuprea browse on one another’s tube-cap community. Shells being attached imbricately is known to increase the ability of D. ornata Moore, 1911 [58] to avoid predatory attacks, while attached algae do not [59]. The shell also changes the composition of the tube-cap community of D. cuprea [60]. Interestingly, Diopatra do not appear to discriminate between shell and algae as attachment material [59], and contrary to popular belief, tube-cap decoration does not appear to impart crypsis [61].
Diopatra cuprea and other Diopatra with emergent tube-caps act as ecosystem engineers by stabilizing the sediment with their tubes and altering flow dynamics with emergent tube-caps, therefore increasing the structural complexity and biodiversity of their infaunal habitat [3][62]. Additionally, the tubes and emergent tube-caps of Diopatra when dense provide shelter from disturbance and predation [63] and may actively facilitate the attachment of some seaweed species or other fouling fauna that cannot survive on soft bottoms without these structures [64][65][66]. Furthermore, Diopatra constitutes an important food source for many species, such as crabs, fish, and birds, as well as acting as a host for parasites [67][68][69][70].
Although the worms can be solitary, they often occur in aggregations that have been reported to be up to 21,800 per m2 of D. dexiognatha Paxton and Bailey-Brock, 1986 [71], a 1.5–5 cm worm, on the shore of Oahu, Hawaii [62][71]. At densities over 6 per 0.01 m2 of D. cuprea, a worm 10 to 20 times larger than D. dexiognatha, predation, disturbance, and erosion are dramatically altered [3][63], leading to increased infaunal abundances. Shorebirds, large crabs, Limulus, and some fish are all inhibited by such densities of Diopatra, while rays are not (Luckenbach, 1984; Woodin et al., 2019) [72][73]. A review of the ecology of Diopatra is presented in this SI (see paper by Berke, 2022) [74].
References
- Kinberg, J.G.H. Annulata nova. Kong Vetensk. Akad. Forhandl. Stockh. 1865, 21, 559–574.
- Audouin, J.V.; Milne Edwards, H. Classification des annélides, et description de celles qui habitent les côtes de la France. Ann. Sci. Nat. 1833, 28, 187–247.
- Woodin, S.A. Refuges, disturbance and community structure: A marine soft-bottom example. Ecology 1978, 59, 274–284.
- Pires, A.; Paxton, H.; Quintino, V.; Rodrigues, A.M. Diopatra (Annelida: Onuphidae) diversity in European waters with the description of Diopatra micrura, new species. Zootaxa 2010, 2395, 17–33.
- Paxton, H.; Arias, A. Unveiling a surprising diversity of the genus Diopatra Audouin & Milne Edwards, 1833 (Annelida: Onuphidae) in the Macaronesion region (eastern North Atlantic) with the description of four new species. Zootaxa 2017, 4300, 505–535.
- Seixas, V.C.; Steiner, T.M.; Solé-Cava, A.M.; Amaral, A.C.Z.; Paiva, P.C. Hidden diversity within the Diopatra cuprea complex (Annelida: Onuphidae): Morphological and genetics analyses reveal four new species in the south-west Atlantic. Zool. J. Linn. Soc. 2021, 191, 637–671.
- Berke, S.K. Functional Groups of Ecosystem Engineers: A Proposed Classification with Comments on Current Issues. Integr. Comp. Biol. 2010, 50, 147–157.
- Berke, S.K.; Mahon, A.R.; Lima, F.P.; Halanych, K.M.; Wethey, D.S.; Woodin, S.A. Range shifts and species diversity in marine ecosystem engineers: Patterns and predictions for European sedimentary habitats. Glob. Ecol. Biogeogr. 2010, 19, 223–232.
- Freitas, R.; Almeida, A.; Pires, A.; Velez, C.; Calisto, V.; Schneider, R.J.; Esteves, V.I.; Wrona, F.J.; Figueira, E.; Soares, A.M.V.M. The effects of carbamazepine on macroinvertebrate species: Comparing bivalves and polychaetes biochemical responses. Water Res. 2015, 85, 137–147.
- Arias, A.; Richter, A.; Anadón, N.; Paxton, H. Evidence of simultaneous hermaphroditism in the brooding Diopatra marocensis (Annelida: Onuphidae) from northern Spain. J. Mar. Biol. Assoc. UK 2013, 93, 1533–1542.
- Arias, A.; Paxton, H. The cryptogenic bait worm Diopatra biscayensis Fauchald et al., 2012 (Annelida: Onuphidae)—Revisiting its history, biology and ecology. Estuar. Coast. Shelf Sci. 2015, 163, 22–36.
- Arias, A.; Paxton, H.; Budaeva, N. Redescription and biology of Diopatra neapolitana (Annelida: Onuphidae), a protandric hermaphrodite with external spermaducal papillae. Estuar. Coast. Shelf Sci. 2016, 174, 1–17.
- Bosc, L.A.G. Histoire Naturelle des Vers: Contenant Leur Description et Leurs Moeurs, Avec Figures Dessinées D’Après Nature; DetervilleL: Paris, France, 1802; Volume 1, pp. 1–10.
- Read, G.; Fauchald, K. World Polychaete Database. Available online: https://www.marinespecies.org/polychaete (accessed on 6 April 2023).
- Augener, H. Polychaeta. Beitr. Kenntn. Meeresfauna Westafr. 1918, 2, 343–362.
- Hartman, O. Polychaetous annelids. Part 5. Eunicea. Allan Hancock Pacific Exped. 1944, 10, 1–238.
- Paxton, H. Diopatra Audouin and Milne Edwards (Polychaeta: Onuphidae) from Australia, with a discussion of developmental patterns in the genus. Beagle Rec. N. Territ. Mus. Arts Sci. 1993, 10, 115–154.
- Rodrigues, A.M.; Pires, A.; Mendo, S.; Quintino, V. Diopatra neapolitana and Diopatra marocensis from the Portuguese coast: Morphological and genetic comparison. Estuar. Coast. Shelf Sci. 2009, 85, 609–617.
- Fauvel, P. Contribution à la faune des Annélides Polychètes du Maroc. Mém. Soc. Sci. Nat. Maroc. 1936, 43, 1–143.
- Delle Chiaje, S. Descrizione e Notomia degli Animali Invertebrati della Silicia Citeriore osservati vivi negli anni 1822–1830. In Tomo 3 Molluschi Acfali, Bracciopedi, Ciropedi, Crostacei, Anellosi; Stabilimento Tipografico di C. Batelli e Comp: Naples, Italy, 1841; p. 142.
- Fauchald, K. Onuphidae (Polychaeta) from Western Mexico. Allan Hancock Monog. Mar. Biol. 1968, 3, 1–82.
- Paxton, H. The Diopatra chiliensis confusion—Redescription of D. chiliensis (Polychaeta, Onuphidae) and implicated species. Zool. Scr. 1998, 27, 31–48.
- Paxton, H. Diopatra Audouin and Milne Edwards (Polychaeta: Onuphidae) from Thailand. Phuket. Mar. Centr. Spec. Publ. 2002, 24, 101–114.
- Hektoen, M.H.; Willassen, E.; Budaeva, N. Phylogeny and cryptic diversity of Diopatra (Onuphidae, Annelida) in the East Atlantic. Biology 2021, 10, 327.
- Sotka, E.E.; Bell, T.; Berke, S. Cryptic mtDNA diversity of Diopatra cuprea (Onuphidae, Annelida) in the northwestern Atlantic Ocean. Biology 2023, 12, 521.
- Pflugfelder, O. Histogenetische und organogenetische Prozesse bei der Regeneration polychaeter Anneliden. I. Regeneration des Vorderendes von Diopatra amboinensis Aud. et M. Edw. Zeitsch. Wiss. Zool. 1929, 133, 121–210.
- Paxton, H. Phylogeny of Eunicida (Annelida) based on morphology of jaws. Zoosymposia 2009, 2, 241–264.
- Paxton, H.; Eriksson, M.E. Ghosts from the past—Ancestral features reflected in the jaw ontogeny of the polychaetous annelids Marphysa fauchaldi (Eunicidae) and Diopatra aciculata (Onuphidae). GFF Scand. J. Earth Sci. 2012, 134, 309–316.
- Paxton, H.; Safarik, M. Jaw growth and replacement in Diopatra aciculata (Annelida: Onuphidae). Beagle Rec. Mus. Art. Gall. North. Territ. 2008, 24, 15–21.
- Clemo, W.C.; Dorgan, K.M. Functional morphology of eunicidan (Polychaeta) jaws. Biol. Bull. 2017, 233, 227–241.
- Monro, C.C.A. Polychaete worms. Discov. Rep. 1930, 2, 1–222.
- Day, J.H. The polychaete fauna of South Africa. Part 5. Errant species dredged off Cape coasts. Ann. S. Afr. Mus. 1960, 45, 261–373.
- Kirkegaard, J.B. The Polychaeta of West Africa. Part II. Errant species. 2. Nephtyidae to Dorvilleidae. Atlantide Rep. 1988, 14, 1–82.
- Pettibone, M.H. Marine polychaete worms of the New England region. I. Aphroditidae through Trochochaetidae. Bull. U. S. Nat. Mus. 1963, 227, 1–356.
- Escobar-Ortega, D.; Couceiro, L.; Muíno, R.; No, E.; Fernández, N. Insights into environmental drivers on the reproductive cycle of Diopatra neapolitana (Polychaeta: Onuphidae). Biology 2022, 11, 1504.
- Fauchald, K.; Berke, S.K.; Woodin, S.A. Diopatra (Onuphidae: Polychaeta) from intertidal sediments in southwestern Europe. Zootaxa 2012, 3395, 47–58.
- Paxton, H.; Fadlaoui, S.; Lechapt, J.P. Diopatra marocensis, a new brooding species of Onuphidae (Annelida: Polychaeta). J. Mar. Biol. Assoc. UK 1995, 75, 949–955.
- Pires, A.; Quintino, V.; Gentil, F.; Freitas, R.; Rodrigues, A.M. Reproductive biology of a brooding Diopatra species: Diopatra marocensis Paxton et al., 1995. Estuar. Coast. Shelf Sci. 2012, 110, 85–92.
- Charnov, E.L. The Theory of Sex Allocation; Princeton University Press: Princeton, NJ, USA, 1982.
- Ghiselin, M.T. Evolutionary aspects of marine invertebrate reproduction. In Reproduction of Marine Invertebrates, General Aspects: Seeking Unity in Diversity, Vol. IX; Giese, A.C., Pierce, J.S., Pierce, V.B., Eds.; Blakewell Scientific: Palo Alto, CA, USA; Boxwood Press: Pacific Grove, CA, USA, 1987; pp. 609–665.
- Sella, G. Sexual strategies and mating systems. In Reproductive Biology and Phylogeny of Annelida, Reproductive Biology and Phylogeny: 4; Rouse, G., Pleijel, F., Eds.; Science Publishers: London, UK, 2006; pp. 82–84.
- Pires, A.; Gentil, F.; Quintino, V.; Rodrigues, A.M. Reproductive biology of Diopatra neapolitana (Annelida, Onuphidae), an exploited natural resource in Ria de Aveiro (Northwestern Portugal). Mar. Ecol. 2012, 33, 56–65.
- Dağli, E.; Ergen, Z.; Çinar, M.E. One-year observation on the population structure of Diopatra neapolitana Delle Chiaje (Polychaeta: Onuphidae) in Izmir Bay (Aegean Sea, eastern Mediterranean). Mar. Ecol. 2005, 26, 265–272.
- Lieber, A. Zur Oogenese einiger Diopatraarten. Z. Wiss. Zool. 1931, 138, 580–649.
- Budaeva, N.; Fauchald, K. Diopatra tuberculantennata, a new species of Onuphidae (Polychaeta) from Belize with a key to onuphids from the Caribbean Sea. Zootaxa 2008, 1795, 29–45.
- Fadlaoui, S.; Lechapt, J.P.; Retiere, C. Larval development of the Onuphid Diopatra marocensis (Annelida: Polychaeta) from the Atlantic Coast of Morocco. J. Mar. Biol. Assoc. UK 1995, 75, 957–966.
- Allen, M.J. Embryological development of the polychaetous annelid, Diopatra cuprea (Bosc). Biol. Bull. Mar. Biol. Lab. Woods Hole 1959, 116, 339–361.
- Cazaux, C. Recherches sur l’ecologie et le developpement larvaires des polychetes de la region d’Arcachon. Fac. Des. Sci. Bordx. 1970, 1, 355.
- Conti, G.; Massa, F. Esperienze di allevamento del polichete Diopatra neapolitana Delle Chiaje, 1841 Nella Laguna di S. Gilla (Sardegna Meridionale). Biol. Mar. Medit. 1998, 5, 1473–1480.
- Emerson, R.R. The Biology of a Population of Diopatra Ornata at Santa Catalina Island, California, Biology; University of Southern California: Los Angeles, CA, USA, 1975; p. 325.
- Fauchald, K. Life diagram patterns in benthic polychaetes. Proc. Biol. Soc. Wash. 1983, 96, 160–177.
- Choe, S. On the life history of the polychaete worm Diopatra neapolitana Delle Chiaje. Bull. Jpn. Soc. Sci. Fish. 1960, 26, 430–437.
- Krishnan, G. The development of Diopatra variabilis (Southern). Z. Wiss. Zool. 1936, 147, 513–525.
- Wethey, D.S.; Woodin, S.A.; Berke, S.K.; Dubois, S.F. Climate hindcasts: Exploring the disjunct distribution of Diopatra biscayensis. Invert. Biol. 2016, 135, 345–356.
- Mangum, C.P.; Cox, C.D. Analysis of the feeding response in the onuphid polychaete Diopatra cuprea (Bosc). Biol. Bull. 1971, 140, 215–229.
- Mangum, C.P.; Santos, S.L.; Rhodes, W.R., Jr. Distribution and feeding in the onuphid polychaete, Diopatra cupea (Bosc). Mar. Biol. 1968, 2, 33–40.
- Brenchley, G.A.; Tidball, J.A. Tube-cap orientations of Diopatra cuprea (Bosc) (Polychaeta): The compromise between physiology and foraging. Mar. Behav. Physiol. 1980, 7, 1–13.
- Moore, J.P. The polychaetous annelids dredged by the U.S.S. “Albatross” off the coast of southern California in 1904; III. Euphrosynidae to Goniadidae. Proc. Acad. Nat. Sci. Phila. 1911, 63, 234–318.
- Brenchley, G.A. Predator detection and avoidance: Ornamentation of tube-caps of Diopatra spp. (Polychaeta: Onuphidae). Mar. Biol. 1976, 38, 179–188.
- Brenchley, G.A. Competition, Disturbance, and Community Structure: The Importance of Physical Structure in a Marine Epifaunal Assemblage, Biological Sciences; University of Maryland: College Park, MD, USA, 1975; p. 98.
- Berke, S.K.; Woodin, S.A. Tube decoration may not be cryptic for Diopatra cuprea (Polychaeta: Onuphidae). Biol. Bull. 2008, 214, 50–56.
- Bailey-Brock, J.H. Ecology of the tube-building polychaete Diopatra leuckarti Kinberg, 1865 (Onuphidae) in Hawaii: Community structure, and sediment stabilizing properties. Zool. J. Linn. Soc. 1984, 80, 191–199.
- Woodin, S.A. Disturbance and Community Structure in a Shallow Water Sand Flat. Ecology 1981, 62, 1052–1066.
- Berke, S. Biogeographic variability in ecosystem engineering: Patterns in the abundance and behavior of the tube-building polychaete Diopatra cuprea. Mar. Ecol. Progr. Ser. 2012, 447, 1–13.
- Thomsen, M.S.; McGlathery, K. Facilitation of macroalgae by the sedimentary tube forming polychaete Diopatra cuprea. Estuar. Coast. Shelf Sci. 2005, 62, 63–73.
- Arias, A. Personal Observation; University of Oviedo: Oviedo, Spain, 2018.
- Rangel, L.G.; Santos, M.J. Diopatra neapolitana (Polychaeta: Onuphidae) as a second intermediate host of Gymnophallus choledochus (Digenea: Gymnophallidae) in the Aveiro Estuary (Portugal): Distribution within the host and histopathology. J. Parasitol. 2009, 95, 123–1236.
- Watanabe, K.; Tamai, K.; Numachi, K. Feeding-habits and distribution pattern of 2 forms of Flatfishes (Genus Pleuronichthys) in Kiisuido Channel waters off the Pacific Coast of Central Japan. Nippon Suisan Gakkaishi 1992, 58, 2253–2260.
- Bowman, R.E.; Stillwell, C.E.; Michaels, W.L.; Grosslein, M.D. Food of Northwest Atlantic Fishes and Two Common Species of Squid. NOAA Tech Memos. NMFS-NE-155. 2000. Available online: https://repository.library.noaa.gov/view/noaa/3140 (accessed on 6 June 2023).
- Steimle, F. Demersal Fish and American Lobster Diets in the Lower Hudson-Raritan Estuary; US Department of Commerce; National Oceanic and Atmospheric Administration, National Marine Fisheries Service, Northeast Region, Northeast Fisheries Science Center: Woods Hole, MA, USA, 2000.
- Paxton, H.; Bailey-Brock, J.H. Diopatra dexiognatha, a new species of Onuphidae (Polychaeta) from Oahu, Hawaiian Islands. Pac. Sci. 1986, 40, 1–6.
- Luckenbach, M.W. Biogenic structure and foraging by five species of shorebirds (Charadrii). Estuar. Coast. Shelf Sci. 1984, 19, 691–696.
- Woodin, S.A.; Bell, S.S.; Grant, J.; Snelgrove, P.V.R.; Wethey, D.S. Interactions and processes in shallow water muddy sands of the northwestern Atlantic coast. In Interactions in the Marine Benthos; Hawkins, S.J., Bohn, K., Firth, L.B., Williams, G.A., Eds.; Cambridge University Press: Cambridge, UK, 2019; pp. 128–163.
- Berke, S.K. A review of Diopatra ecology: Current knowledge, open question, and future threats for an ecosystem engineering polychaete. Biology 2022, 11, 1485.
More
Information
Subjects:
Biology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
898
Revisions:
3 times
(View History)
Update Date:
05 Sep 2023
Table of Contents




Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No